Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress


Abstract
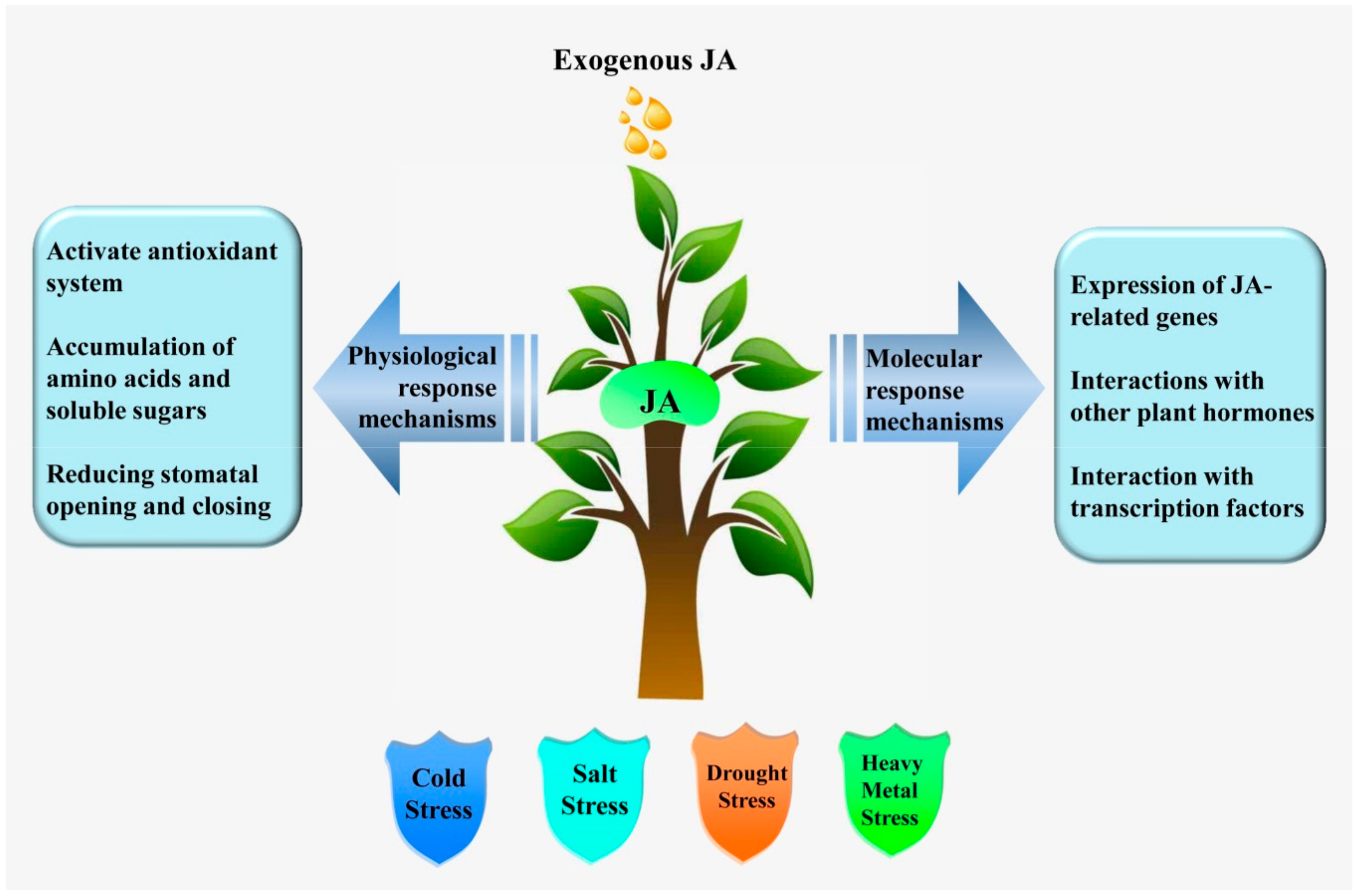
:1. Introduction
2. JA-Mediated Abiotic Stress Responses
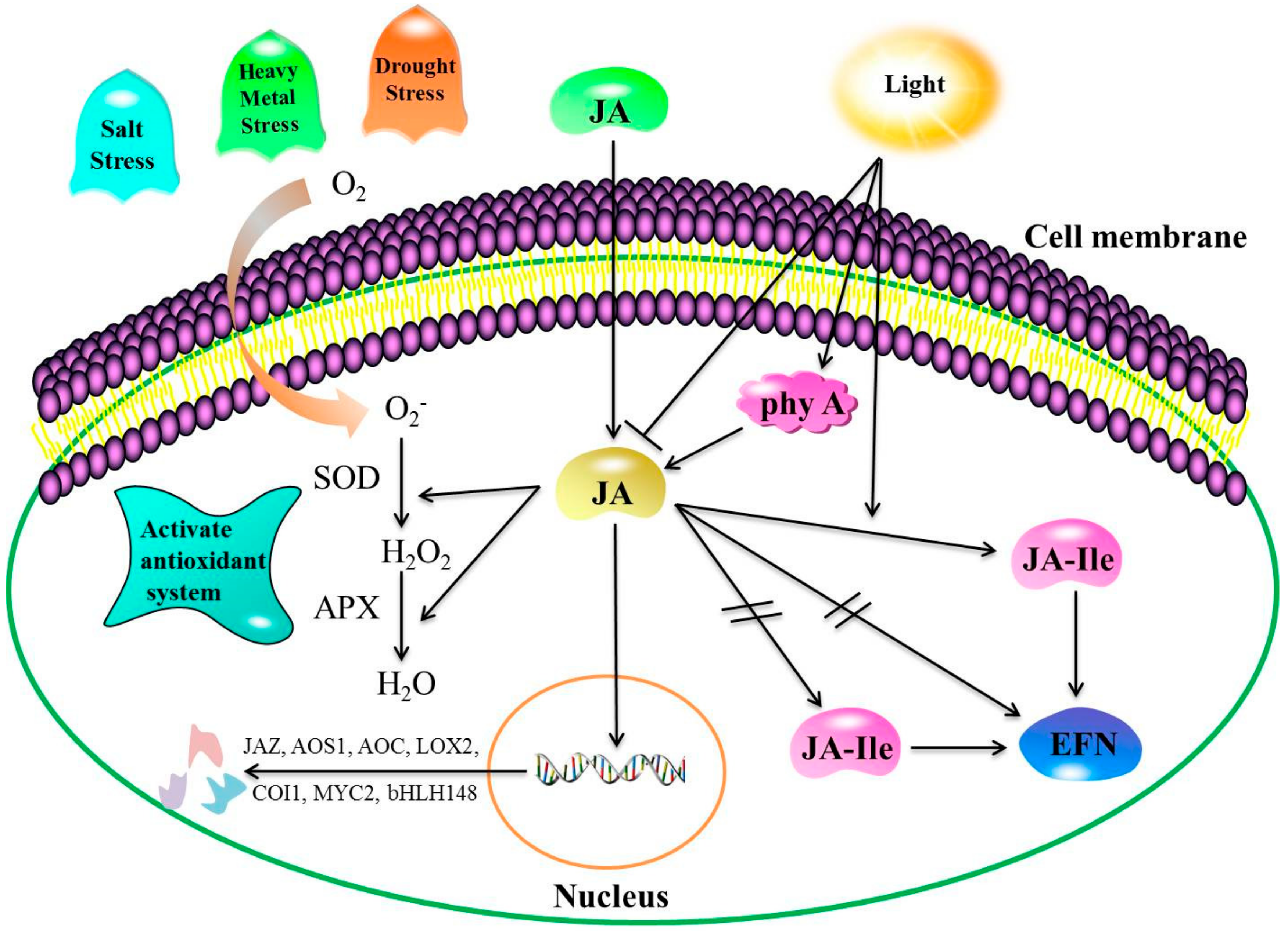
2.1. Cold Stress
2.2. Drought Stress
2.3. Salt Stress
2.4. Heavy Metal Stress
2.5. Light Stress
2.6. Other Stress Factors
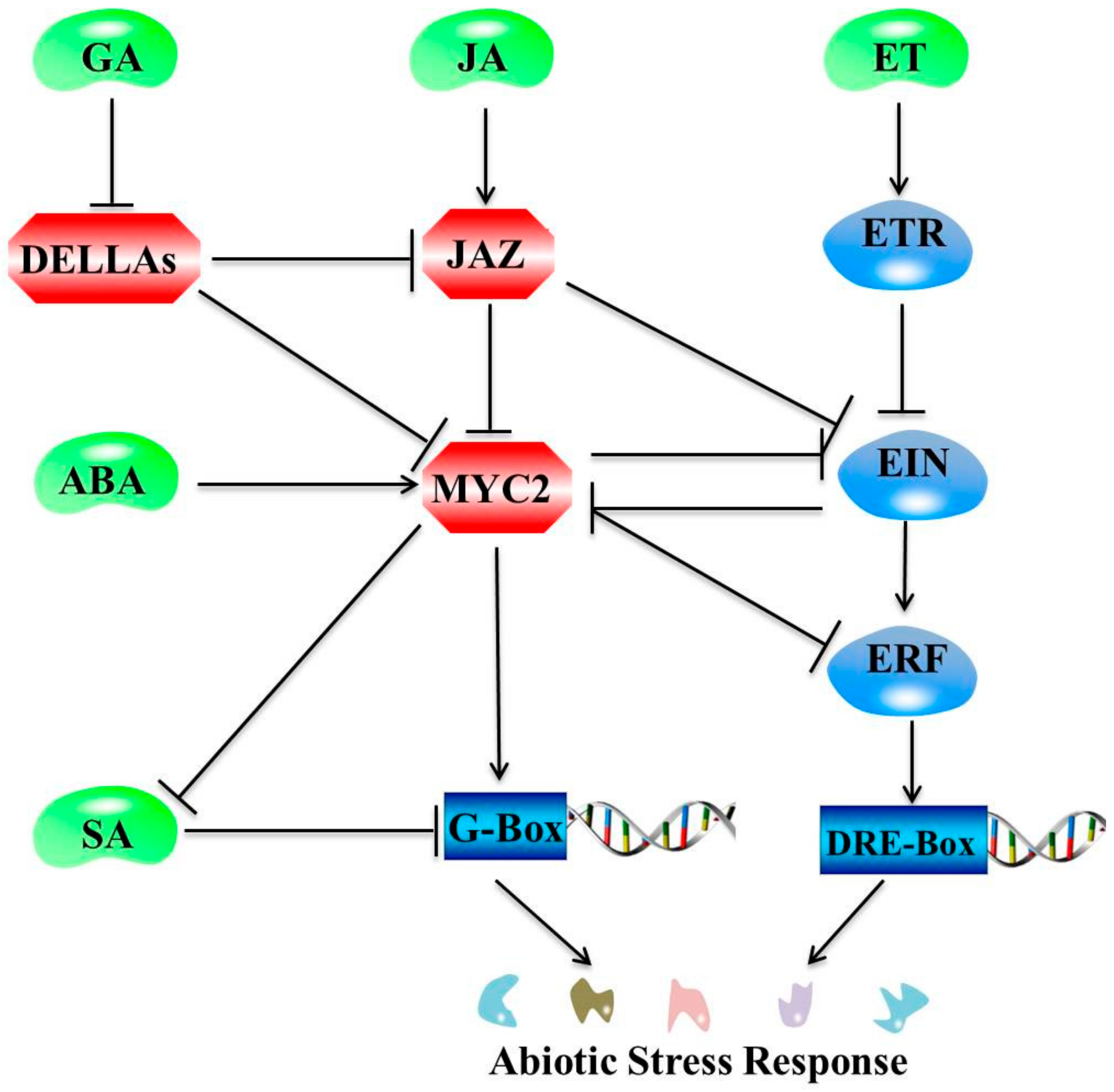
3. Interactions between JA and Plant Hormone Pathways under Abiotic Stresses
3.1. Interactions between JA and ABA Pathways in Response to Abiotic Stresses
3.2. Interactions between JA and Ethylene Pathways under Abiotic Stresses
3.3. The Interactions between JA and SA Pathways under Abiotic Stresses
3.4. Interactions between JA and other Plant Hormone Pathways under Abiotic Stress
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| JA | Jasmonic acid |
| JAZ | Jasmonate ZIM-domain proteins |
| MeJA | Methyl jasmonate |
| JA-Ile | Jasmonate isoleucine conjugate |
| JAs | Jasmonates |
| TF | Transcription factor |
| ABA | Abscisic acid |
| ET | Ethylene |
| SA | Salicylic acid |
| GA | Gibberellin |
| IAA | Indole-3-acetic acid |
| BR | Brassinosteroids |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| bHLH148 | basic helix-loop-helix protein 148 |
| AOC | Allene oxide cyclase |
| AOS1 | Allene oxide synthase1 |
| LOX2 | Lipoxygenase2 |
| CBF | C-repeat binding factor |
| ICE-CBF | Inducer of CBF expression |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| APX | Ascorbate peroxidase |
| JAZ | Jasmonate ZIM-domain proteins |
| ROS | Reactive oxygen species |
| MDA | Malondialdehyde |
| EFN | Extra-floral nectar |
| FR | Far-red |
| FIN219 | Far-red insensitive 219 |
| CCT1 | C terminus of cryptochrome 1 |
| ETR | Electron transport rate |
| Pb | Lead |
| Ni | Nickel |
| Cd | Cadmium |
| Mn | Manganese |
| Se | Selenium |
| HO-1 | Hemeoxygenase-1 |
| O3 | Ozone |
| PIF3 | Phytochrome-interacting factor 3 |
| 12-OPDA | 12-oxo-phytodienoic acid |
| POD | Peroxidase |
| GPX | Glutathione peroxidase |
| GR | Glutathione reductase |
| CRY1 | Cryptochrome 1 |
| PSII | Photosystem II |
| NPQ | Non-photochemical quenching |
| EIN | Ethylene insensitive |
| AP2/ERF | APETALA2/Ethylene responsive factor |
References
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Duan, G.H.; Li, C.Q.; Liu, L.; Han, G.Y.; Zhang, Y.L.; Wang, C.M. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.J.; Zhou, Y.X.; Zhou, M.L.; Yan, J.; Khurshid, M.; Weng, W.F.; Cheng, J.P.; Zhang, K.X. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, M.L.; Kang, J.H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Llanes, A.; Andrade, A.; Alenabo, S.; Luna, V. Alterations of endogenous hormonal levels in plants under drought and salinity. Am. J. Plant Sci. 2016, 7, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in jasmonate signaling for multistress resilience. Annu. Rev. Entomol. 2018, 69, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.K.; Pi, Y.; Hou, S.; Tang, K.X. Jasmonate biosynthetic pathway: Its physiological role and potential application in plant secondary metabolic engineering. Chin. Bull. Bot. 2010, 45, 137–148. [Google Scholar]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Karpets, Y.V.; Kolupaev, Y.E.; Lugovaya, A.A.; Oboznyi, A.I. Effect of jasmonic acid on the pro-/antioxidant system of wheat coleoptiles as related to hyperthermia tolerance. Russ. J. Plant Physiol. 2014, 61, 339–346. [Google Scholar] [CrossRef]
- Wasternack, C. Action of jasmonates in plant stress responses and development—Applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.R.; Assmann, S.M. Hormone interactions in stomatal function. Plant Mol. Biol. 2009, 69, 451–462. [Google Scholar] [CrossRef]
- Hu, Y.R.; Jiang, Y.J.; Han, X.; Wang, H.P.; Pan, J.J.; Yu, D.Q. Jasmonate regulates leaf senescence and tolerance to cold stress: Crosstalk with other phytohormones. J. Exp. Bot. 2017, 68, 1361–1369. [Google Scholar] [CrossRef]
- Robson, F.; Okamoto, H.; Patrick, E.; Harris, S.R.; Wasternack, C.; Brearley, C.; Turner, J.G. Jasmonate and phytochrome a signaling in Arabidopsis wound and shade responses are integrated through JAZ1 stability. Plant Cell 2010, 22, 1143–1160. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.L.; Wang, J.N.; Shan, W.; Fan, J.G.; Kuang, J.F.; Wu, K.Q.; Li, X.P.; Chen, W.X.; He, F.Y.; Chen, J.Y.; et al. Induction of jasmonate signalling regulators MaMYC2s and their physical interactions with MaICE1 in methyl jasmonate-induced chilling tolerance in banana fruit. Plant Cell Environ. 2013, 36, 30–51. [Google Scholar] [CrossRef]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef]
- Huang, B.R.; DaCosta, M.; Jiang, Y.W. Research advances in mechanisms of turfgrass tolerance to abiotic stresses: From physiology to molecular biology. Crit. Rev. Plant Sci. 2014, 33, 141–189. [Google Scholar] [CrossRef]
- Trischuk, R.G.; Schilling, B.S.; Low, N.H.; Gray, G.R.; Gusta, L.V. Cold acclimation, de-acclimation and re-acclimation of spring canola, winter canola and winter wheat: The role of carbohydrates, cold-induced stress proteins and vernalization. Environ. Exp. Bot. 2014, 106, 156–163. [Google Scholar] [CrossRef]
- Hincha, D.K.; Zuther, E. Plant Cold Acclimation: Methods and Protocols; Springer: New York, NY, USA, 2014; p. 1166. [Google Scholar]
- Li, S.M.; Yang, Y.; Zhang, Q.; Liu, N.F.; Xu, Q.G.; Hu, L.X. Differential physiological and metabolic response to low temperature in two zoysiagrass genotypes native to high and low latitude. PLoS ONE 2018, 13, e0198885. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.F.; Zheng, Y.H.; Wang, K.T.; Jin, P.; Rui, H.J. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chem. 2009, 115, 1458–1463. [Google Scholar] [CrossRef]
- Abdullah, A.S.; Aziz, M.M.; Siddique, K.H.M.; Flower, K.C. Film antitranspirants increase yield in drought stressed wheat plants by maintaining high grain number. Agric. Water Manag. 2015, 159, 11–18. [Google Scholar] [CrossRef]
- Sun, X.G.; Shi, J.; Ding, G.J. Combined effects of arbuscular mycorrhiza and drought stress on plant growth and mortality of forage sorghum. Appl. Soil Ecol. 2017, 119, 384–391. [Google Scholar] [CrossRef]
- Javed, N.; Ashraf, M.; Akram, N.A.; Ai-Qurainy, F. Alleviation of adverse effects of drought stress on growth and some potential physiological attributes in Maize (Zea mays L.) by seed electromagnetic treatment. Photochem. Photobiol. 2011, 87, 1354–1362. [Google Scholar] [CrossRef]
- Huang, J.; Chen, C.; Zhang, W.X.; Ding, C.J.; Su, X.H.; Huang, Q.J. Effects of drought stress on anatomical structure and photosynthetic characteristics of transgenic JERF36 Populus alba × P. berolinensis seedling leaves. Sci. Silvae Sin. 2017, 119, 384–391. [Google Scholar]
- Munné-bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 8808–8818. [Google Scholar] [CrossRef]
- Ma, X.Q.; Zhang, J.; Burgess, P.; Rosso, S.; Huang, B. Interactive effects of melatonin and cytokinin on alleviating drought-induced leaf senescence in creeping bentgrass (Agrostis stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Lei, Y. Physiological responses of Populus przewalskii to oxidative burst caused by drought stress. Russ. J. Plant Physiol. 2008, 55, 857–864. [Google Scholar] [CrossRef]
- Wang, L.; He, J.; Ding, H.D.; Liu, H.; Lv, B.; Liang, J.S.; Wang, L.; He, J.; Ding, H.D.; Liu, H.; et al. Overexpression of AT14A confers tolerance to drought stress-induced oxidative damage in suspension cultured cells of Arabidopsis thaliana. Protoplasma 2014, 252, 1111–1120. [Google Scholar] [CrossRef]
- Savchenko, T.; Kolla, V.A.; Wang, C.Q.; Nasafi, Z.; Hicks, D.R.; Phadungchob, B.; Chehab, W.E.; Brandizzi, F.; Froehlich, J.; Dehesh, K. Functional convergence of oxylipin and abscisic acid pathways controls stomatal closure in response to drought. Plant Physiol. 2014, 164, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wu, H.; Ma, S.Q.; Xiang, D.H.; Liu, R.Y.; Xiong, L.Z. OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in Rice. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.X.; Zhang, L.J.; Li, F.H.; Chen, Z.B.; Wang, C.; Yao, Y.C.; Han, Z.H.; Zhang, J.; Shi, Z.S. Relationship between jasmonic acid accumulation and senescence in drought-stress. Afr. J. Agric. Res. 2010, 5, 1978–1983. [Google Scholar]
- Mohamed, H.I.; Latif, H.H. Improvement of drought tolerance of soybean plants by using methyl jasmonate. Physiol. Mol. Biol. Plants 2017, 23, 545–556. [Google Scholar] [CrossRef]
- Syeed, S.; Anjum, N.A.; Nazar, R.; Iqbal, N.; Masood, A.; Khan, N.A. Salicylic acid-mediated changes in photosynthesis, nutrients content and antioxidant metabolism in two mustard (Brassica juncea L.) cultivars differing in salt tolerance. Acta Physiol. Plant. 2011, 33, 877–886. [Google Scholar] [CrossRef]
- Kadri, K.; Abdellaoui, R.; Mhamed, H.C.; Da Silva, J.A.T.; Naceur, M.B. Analysis of salt-induced mRNA transcripts in tunisian local barley (Hordeum vulgare) leaves identified by differential display RT-PCR. Biochem. Genet. 2014, 52, 106–115. [Google Scholar] [CrossRef]
- Domenico, S.D.; Taurino, M.; Gallo, A.; Poltronieri, P.; Pastor, V.; Flors, V.; Santino, A. Oxylipin dynamics in Medicago truncatula in response to salt and wounding stresses. Physiol. Plant. 2019, 165, 198–208. [Google Scholar] [CrossRef]
- Pedranzani, H.; Racagni, G.; Alemano, S.; Miersch, O.; Ramirez, I.; Pena-Cortes, H.; Taleisnik, E.; Machado-Domenech, E.; Abdala, G. Salt tolerant tomato plants show increased levels of jasmonic acid. Plant Growth Regul. 2003, 41, 149–158. [Google Scholar] [CrossRef]
- Abouelsaad, I.; Renault, S. Enhanced oxidative stress in the jasmonic acid-deficient tomato mutant def-1 exposed to NaCl stress. J. Plant Physiol. 2018, 226, 136–144. [Google Scholar] [CrossRef]
- Bandurska, H.; Stroiński, A.; Kubiś, J. The effect of jasmonic acid on the accumulation of ABA, proline and spermidine and its influence on membrane injury under water deficit in two barley genotypes. Acta Physiol. Plant. 2003, 25, 279–285. [Google Scholar] [CrossRef]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X.; Ismail, A.M.; Close, T.J. Large- scale expression profiling and physiological characterization of jasmonic acid-mediated adaptation of barley to salinity stress. Plant Cell Environ. 2007, 30, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Syeed, S.; Nazar, R.; Anjum, N.A. An insight into the role of salicylic acid and Jasmonic acid in salt stress tolerance. In Phytohormones and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 277–300. [Google Scholar]
- Shahzad, A.N.; Pitann, B.; Ali, H.; Qayyum, M.F.; Fatima, A.; Bakhat, H.F. Maize genotypes differing in salt resistance vary in jasmonic acid accumulation during the first phase of salt stress. J. Agron. Crop Sci. 2015, 201, 443–451. [Google Scholar] [CrossRef]
- Qiu, Z.B.; Guo, J.L.; Zhu, A.J.; Zhang, L.; Zhang, M.M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, M.; Yang, H.H.; Li, X.X.; Cui, Z.J. Physiological responses of Suaeda glauca and Arabidopsis thaliana in phytoremediation of heavy metals. J. Environ. Manag. 2018, 223, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Ali, R.; Alothman, Z.A.; Ali, J.; Habila, M.A. Assessment of toxic metals in wheat crops grown on selected soils, irrigated by different water sources. Arab. J. Chem. 2012, 9, 1555–1562. [Google Scholar]
- Zhao, S.Y.; Ma, Q.F.; Xu, X.; Li, G.Z.; Hao, L. Tomato Jasmonic acid-deficient mutant spr2 seedling response to Cadmium stress. J. Plant Growth Regul. 2016, 35, 603–610. [Google Scholar] [CrossRef]
- Sirhindi, G.; Mir, M.A.; Sharma, P.; Gill, S.S.; Kaur, H.; Mushtaq, R. Modulatory role of jasmonic acid on photosynthetic pigments, antioxidants and stress markers of Glycine max L. under nickel stress. Physiol. Mol. Biol. Plants 2015, 21, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azeem, U. Ameliorating Nickel stress by Jasmonic acid treatment in Zea mays L. Russ. Agric. Sci. 2018, 44, 209–215. [Google Scholar] [CrossRef]
- Noriega, G.; Cruz, D.S.; Batlle, A.; Tomaro, M.; Balestrasse, K. Heme oxygenase is involved in the protection exerted by Jasmonic acid against Cadmium stress in Soybean roots. J. Plant Growth Regul. 2012, 31, 79–89. [Google Scholar] [CrossRef]
- Carvalho, R.F.; Campos, M.L.; Azevedo, R.A. The role of phytochromes in stress tolerance. In Salt Stress Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 283–299. [Google Scholar]
- Kazan, K.; Manners, J.M. The interplay between light and jasmonate signalling during defence and development. J. Exp. Bot. 2011, 62, 4087–4100. [Google Scholar] [CrossRef] [Green Version]
- Izaguirre, M.M.; Mazza, C.A.; Svatos, A.; Baldwin, I.T.; Ballaré, C.L. Solar Ultraviolet-B radiation and insect herbivory trigger partially overlapping phenolic responses in Nicotiana attenuata and Nicotiana longiflora. Ann. Bot. 2007, 99, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhika, V.; Kost, C.; Mithofer, A.; Boland, W. Regulation of extrafloral nectar secretion by jasmonates in lima bean is light dependent. Proc. Natl. Acad. Sci. USA 2010, 107, 17228–17233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemann, M.; Axel, M.; Korte, A.; Furuya, M.; Weiler, E.W.; Nick, P. Impaired induction of the Jasmonate pathway in the rice mutant hebiba. Plant Physiol. 2003, 133, 1820–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.J.; Fu, T.Y.; Yang, S.L.; Hsien, H.L. FIN219/JAR1 and cryptochrome1 antagonize each other to modulate photomorphogenesis under blue light in Arabidopsis. PLoS Genet. 2018, 14, e1007248. [Google Scholar]
- Liu, X.; Chi, H.; Yue, M.; Zhang, X.F.; Li, W.J.; Jia, E.P. The regulation of exogenous Jasmonic acid on UV-B stress tolerance in Wheat. Plant Growth Regul. 2012, 31, 436–447. [Google Scholar] [CrossRef]
- Ahmad, P.; Alyemeni, M.N.; Vijaya, L.; Alam, P.; Ahanger, M.A.; Alamri, S.A. Jasmonic acid alleviates negative impacts of cadmium stress by modifying osmolytes and antioxidants in faba bean (Vicia faba L.). Arch. Agron. Soil Sci. 2017, 63, 1–11. [Google Scholar] [CrossRef]
- Demkura, P.V.; Abdala, G.; Baldwin, I.T.; Ballaré, C.L. Jasmonate-dependent and -independent pathways mediate specific effects of solar ultraviolet B radiation on leaf phenolics and antiherbivore defense. Plant Physiol. 2010, 152, 1084–1095. [Google Scholar] [CrossRef] [Green Version]
- Kanna, M.; Tamaoki, M.; Kubo, A.; Nakajima, N.; Rakwal, R.; Agrawal, G.K.; Tamogami, S.; Ioki, M.; Ogawa, D.; Saji, H.; et al. Isolation of an ozone-sensitive and jasmonate-semi-insensitive Arabidopsis mutant (oji1). Plant Cell Physiol. 2003, 44, 1301–1310. [Google Scholar] [CrossRef]
- Rao, M.V.; Lee, H.; Creelman, R.A.; Mullet, J.E.; Davis, K.R. Jasmonic acid signaling modulates ozone-induced hypersensitive cell death. Plant Cell 2000, 12, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Grantz, D.A.; Vu, H.B. Root and shoot gas exchange respond additively to moderate ozone and methyl jasmonate without induction of ethylene: Ethylene is induced at higher O3 concentrations. J. Exp. Bot. 2012, 63, 4303–4313. [Google Scholar] [CrossRef]
- Kaya, A.; Doganlar, Z.B. Exogenous jasmonic acid induces stress tolerance in tobacco (Nicotiana tabacum) exposed to imazapic. Ecotoxicol. Environ. Saf. 2016, 124, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, S.; Cortleven, A.; Iven, T.; Feussner, I.; Havaux, M.; Riefler, M.; Schmülling, T. Circadian stress regimes affect the circadian clock and cause jasmonic acid-dependent cell death in cytokinin-deficient Arabidopsis plants. Plant Cell 2016, 28, 1616–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleman, F.; Yazaki, J.; Lee, M.; Takahashi, Y.; Kim, A.Y.; Li, Z.; Kinoshita, T.; Ecker, J.R.; Schroeder, J.I. An ABA-increased interaction of the PYL6 ABA receptor with MYC2 transcription factor: A putative link of ABA and JA signaling. Sci. Rep. 2016, 6, 28941. [Google Scholar] [CrossRef] [PubMed]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.P.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. Npr3 and npr4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, J.Q.; Zhai, Q.Z.; Zhou, W.K.; Qi, L.L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H.L.; Qi, J.; et al. The basic helix-loop-helix transcription factor MYC2 directly represses plethora expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.P.; Yu, J.Q.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Benefits of brassinosteroid crosstalk. Trends Plant Sci. 2012, 17, 594–605. [Google Scholar] [CrossRef]
- Wasternack, C.; Strnad, M. Jasmonate signaling in plant stress responses and development—Active and inactive compounds. New Biotechnol. 2016, 33, 604–613. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, N.; Virlouvet, L.; Riethoven, J.J.; Fromm, M.; Avramova, Z. Four distinct types of dehydration stress memory genes in Arabidopsis thaliana. BMC Plant Boil. 2013, 13, 229. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cadenas, A.; Vives, V.; Zandalinas, S.I.; Manzi, M.; Sanchez-Perez, A.M.; Perez-Clemente, R.M.; Arbona, V. Abscisic acid: A versatile phytohormone in plant signaling and beyond. Curr. Protein Pept. Sci. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Q.; Wu, W.R.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Exp. Bot. 2008, 59, 2991–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackman, P.; González-Guzmán, M.; Tilleman, S.; Carqueijeiro, I.; Pérez, A.C.; Moses, T.; Seo, M.; Kanno, Y.; Häkkinen, S.T.; Van Montagu, M.C.; et al. Jasmonate signaling involves the abscisic acid receptor PYL4 to regulate metabolic reprogramming in Arabidopsis and tobacco. Proc. Natl. Acad. Sci. USA 2011, 108, 5891–5896. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.R.; Jiang, L.Q.; Wang, F.; Yu, D.Q. Jasmonate regulates the INDUCER OF CBF EXPRESSION-CREPEAT BINDING FACTOR/DRE BINDING FACTOR1 cascade and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, H.; Honda, C.; Kondo, S. Effect of low-temperature stress on abscisic acid, jasmonates, and polyamines in apples. Plant Growth Regul. 2007, 52, 199–206. [Google Scholar] [CrossRef]
- Abe, H. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2002, 15, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Ding, Y.; Fromm, M.; Avramova, Z. Different gene-specific mechanisms determine the ‘revised-response’ memory transcription patterns of a subset of A. thaliana dehydration stress responding genes. Nucleic Acids Res. 2014, 42, 5556–5566. [Google Scholar] [CrossRef] [PubMed]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ollas, C.; Arbona, V.; Gómez-Cadenas, A. Jasmonic acid interacts with abscisic acid to regulate plant responses to water stress conditions. Plant Signal. Behav. 2015, 10, e1078953. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Hsieh, E.J.; Cheng, M.C.; Chen, C.Y.; Hwang, S.Y.; Lin, T.P. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. New Phytol. 2016, 211, 599–613. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.S.; Kim, S.K.; Jang, S.W.; Choo, Y.S.; Sohn, E.Y.; Lee, I.J. Effect of jasmonic acid on endogenous gibberellins and abscisic acid in rice under NaCl stress. Biol. Plant. 2005, 49, 447–450. [Google Scholar] [CrossRef]
- Kim, S.K.; Sohn, E.Y.; Joo, G.J.; Lee, I.J. Influence of jasmonic acid on endogenous gibberellin and abscisic acid in salt-stressed chard plant. J. Environ. Biol. 2009, 30, 333–338. [Google Scholar] [PubMed]
- Wang, Y.; Mopper, S.; Hasenstein, K.H. Effects of salinity on endogenous ABA, IAA, JA, and SA in Iris hexagona. J. Chem. Ecol. 2001, 27, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Brossa, R.; López-Carbonell, M.; Jubany-Marí, T.; Alegre, L. Interplay between Abscisic acid and Jasmonic acid and its role in water-oxidative stress in tild-type, ABA-deficient, JA-deficient, and Ascorbate-deficient Arabidopsis Plants. J. Plant Growth Regul. 2011, 30, 322–333. [Google Scholar] [CrossRef]
- Zhai, Y.; Wang, Y.; Li, Y.J.; Lei, T.T.; Yan, F.; Su, L.T.; Li, X.W.; Zhao, Y.; Sun, X.; Li, J.W.; et al. Isolation and molecular characterization of GmERF7, a soybean ethylene-response factor that increases salt stress tolerance in tobacco. Gene 2013, 513, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Van der Fits, L.; Memelink, J. ORCA3, a Jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 2000, 289, 295–297. [Google Scholar] [CrossRef]
- Fits, L.V.D.; Memelink, J. The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via interaction with a jasmonate-responsive promoter element. Plant J. 2001, 25, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, O.; Piqueras, R.; Sánchez-serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and Jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pre, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.M.; Memelink, J. The AP2/ERF domain transcription factor ORA59 integrates Jasmonic acid and Ethylene signals in plant defense. Plant Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaoki, M.; Freeman, J.L.; Pilon-Smits, E.A. Cooperative ethylene and jasmonic acid signaling regulates selenite resistance in Arabidopsis. Plant Physiol. 2008, 146, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Górnik, K.; Badowiec, A.; Weidner, S. The effect of seed conditioning, short-term heat shock and salicylic, jasmonic acid or brasinolide on sunflower (Helianthus annuus L.) chilling resistance and polysome formation. Acta Physiol. Plant. 2014, 36, 2547–2554. [Google Scholar] [CrossRef] [Green Version]
- Ilyas, N.; Gull, R.; Mazhar, R.; Saeed, M.; Kanwal, S.; Shabir, S.; Bibi, F. Influence of salicylic acid and jasmonic acid on wheat under drought stress. Commun. Soil Sci. Plant Anal. 2017, 48, 2715–2723. [Google Scholar] [CrossRef]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Martínez-Romeroa, D.; Guilléna, F.; Serranob, M.; Valeroa, D. Vapour treatments with methyl salicylate or methyl jasmonate alleviated chilling injury and enhanced antioxidant potential during postharvest storage of pomegranates. Food Chem. 2010, 124, 964–970. [Google Scholar] [CrossRef]
- Ndamukong, I.; Abdallat, A.A.; Thurow, C.; Fode, B.; Zander, M.; Weigel, R.; Gatz, C. SA-inducible Arabidopsis glutaredoxin interacts with TGA factors and suppresses JA-responsive PDF1.2 transcription. Plant J. 2010, 50, 128–139. [Google Scholar] [CrossRef]
- Tuteja, N.; Gill, S.S. Salicylic acid: A novel plant growth regulator-role in physiological processes and abiotic stresses under changing environments. In Climate Change and Plant Abiotic Stress Tolerance; Wiley-VCH: Weinheim, Germany, 2013; pp. 939–990. [Google Scholar]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotox. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef]
- Meldau, S.; Ullman-Zeunert, L.; Govind, G.; Bartram, S.; Baldwin, I.T. MAPK-dependent JA and SA signalling in Nicotiana attenuate affects plant growth and fitness during competition with conspecifics. BMC Plant Biol. 2012, 12, 213. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.L.; Yao, J.; Mei, C.S.; Tong, X.H.; Zeng, L.J.; Li, Q.; Xiao, L.T.; Sun, T.P.; Li, J.G.; Deng, X.W.; et al. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, 1192–1200. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.L.; Ding, L.H.; Yu, H. Crosstalk between GA and JA signaling mediates plant growth and defense. Plant Cell Rep. 2013, 32, 1067–1074. [Google Scholar] [CrossRef]
- Guo, Q.; Major, I.T.; Howe, G.A. Resolution of growth-defense conflict: Mechanistic insights from jasmonate signaling. Curr. Opin. Plant Biol. 2018, 44, 72–81. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of Stress | Plant Species | JA | Regulation Mechanism | Reference |
|---|---|---|---|---|
| Freezing | Arabidopsis thaliana | Endogenous | Positively regulated the C-repeat binding factor (CBF) transcriptional pathway to up-regulate downstream cold-responsive genes | [15] |
| Chilling | Musa acuminata | Endogenous | Induced MaMYC2 and inducer of CBF expression (ICE-CBF) cold-responsive pathway gene expression, including MaCBF1, MaCBF2, MaCOR1, MaKIN2, MaRD2, and MaRD5 | [17] |
| Chilling and freezing | Zoysia japonica | Endogenous | Up-regulated ZjCBF, ZjDREB1, and ZjLEA expression | [22] |
| Chilling | Eriobotrya japonica | Exogenous (10 μM) | Enhanced antioxidant enzyme activity and higher unsaturated/saturated fatty acid ratio | [23] |
| Drought | Arabidopsis thaliana | Endogenous | Produced higher 12-OPDA levels and reduced stomatal aperture | [32] |
| Drought | Oryza sativa. | Endogenous | OsJAZ1 was a negative regulator via the abscisic acid (ABA)-dependent and JA-dependent pathways. | [33] |
| Drought | Oryza sativa. | Endogenous | OsbHLH148 acted on the JA signaling pathway with OsJAZ1 and OsCOI1, constituting an OsbHLH148–OsJAZ–OsCOI1 signaling module | [18] |
| Drought | Prunus armeniaca | Exogenous (50 µM) | Increased malondialdehyde (MDA) levels and promoted leaf senescence | [34] |
| Drought | Glycine max | Exogenous (20 μM) | Increased cell wall fractionation, saturated and unsaturated fatty acid, flavonoid, phenolic acid, and sugar fraction content | [35] |
| Salt | Lycopersicon esculentum | Endogenous | Increased lipoxygenase (LOX), AOS-mRNA, and Pin2-mRNA accumulation | [39] |
| Salt | Solanum lycopersicum | Endogenous | Activated both enzymatic and non-enzymatic ROS antioxidants | [40] |
| Salt | Zea mays | Exogenous (25 μM) | Improved Na+ exclusion by decreasing Na+ uptake | [44] |
| Salt | Triticum aestivum | Exogenous (2 mM) | Decreased the concentration of MDA and H2O2, and increased the transcript levels and activities of SOD, POD, catalase (CAT), and APX | [45] |
| Heavy metal (cadmium) | Lycopersicon esculentum | Endogenous | JA played a positive regulatory role in tomato plant response to Cd stress by regulating the antioxidant system | [48] |
| Heavy metal (nickel) | Glycine max | Exogenous (1 μM and 1 nM) | Managed the antioxidant machinery and protected the DNA synthesis of total proteins to mitigate Ni stress | [49] |
| Heavy metal (nickel) | Zea mays | Exogenous (10 μM) | JA alleviated the negative impact of Ni-treated plants by improving the activity of antioxidant enzymes SOD, CAT, APX, GPX, and GR | [50] |
| Heavy metal (cadmium) | Vicia faba | Exogenous (10 μM) | Inhibited the accumulation of Cd, H2O2, and MDA, and enhanced osmolyte and antioxidant activities that reduce oxidative stress | [59] |
| Heavy metal (cadmium) | Glycine max | Exogenous (20 μM) | Augmented the activities of antioxidant enzymes CAT and SOD to Cd treatment | [51] |
| Light and darkness | Phaseolus lunatus | Endogenous | JA-Ile enhanced EFN secretion under light conditions, yet did not reduce EFN secretion in the dark | [55] |
| Light and darkness | Oryza sativa | Endogenous | JA and phytochrome A signaling were integrated through degradation of the JAZ1 protein | [56] |
| Far-red | Arabidopsis thaliana | Exogenous (50 μM) | Interaction of the photoreceptor CRY1 and the JA-conjugating enzyme FR-insensitive219/JAR1 | [57] |
| UV-B | Triticum aestivum | Exogenous (1 and 2.5 mM) | Increased reaction centers’ excitation energy capture efficiency, effective PSII, and electron transport rate (ETR), and decreased NPQ | [58] |
| Ozone stress | Arabidopsis thaliana | Exogenous (100 μM) | Inhibited the spread of programmed cell death | [62] |
| Imazapic stress | Nicotiana tabacum | Exogenous (45 μM) | Increased antioxidant activity and phytohormone level and decreased MDA content | [64] |
| Circadian stress | Arabidopsis thaliana | Endogenous | Reduced the cell death phenotype | [65] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 1446. https://doi.org/10.3390/ijms21041446
Wang J, Song L, Gong X, Xu J, Li M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. International Journal of Molecular Sciences. 2020; 21(4):1446. https://doi.org/10.3390/ijms21041446
Chicago/Turabian StyleWang, Jia, Li Song, Xue Gong, Jinfan Xu, and Minhui Li. 2020. "Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress" International Journal of Molecular Sciences 21, no. 4: 1446. https://doi.org/10.3390/ijms21041446
APA StyleWang, J., Song, L., Gong, X., Xu, J., & Li, M. (2020). Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. International Journal of Molecular Sciences, 21(4), 1446. https://doi.org/10.3390/ijms21041446