Molecular Interactions of the Polysialytransferase Domain (PSTD) in ST8Sia IV with CMP-Sialic Acid and Polysialic Acid Required for Polysialylation of the Neural Cell Adhesion Molecule Proteins: An NMR Study

Abstract
:1. Introduction
2. Results and Discussion
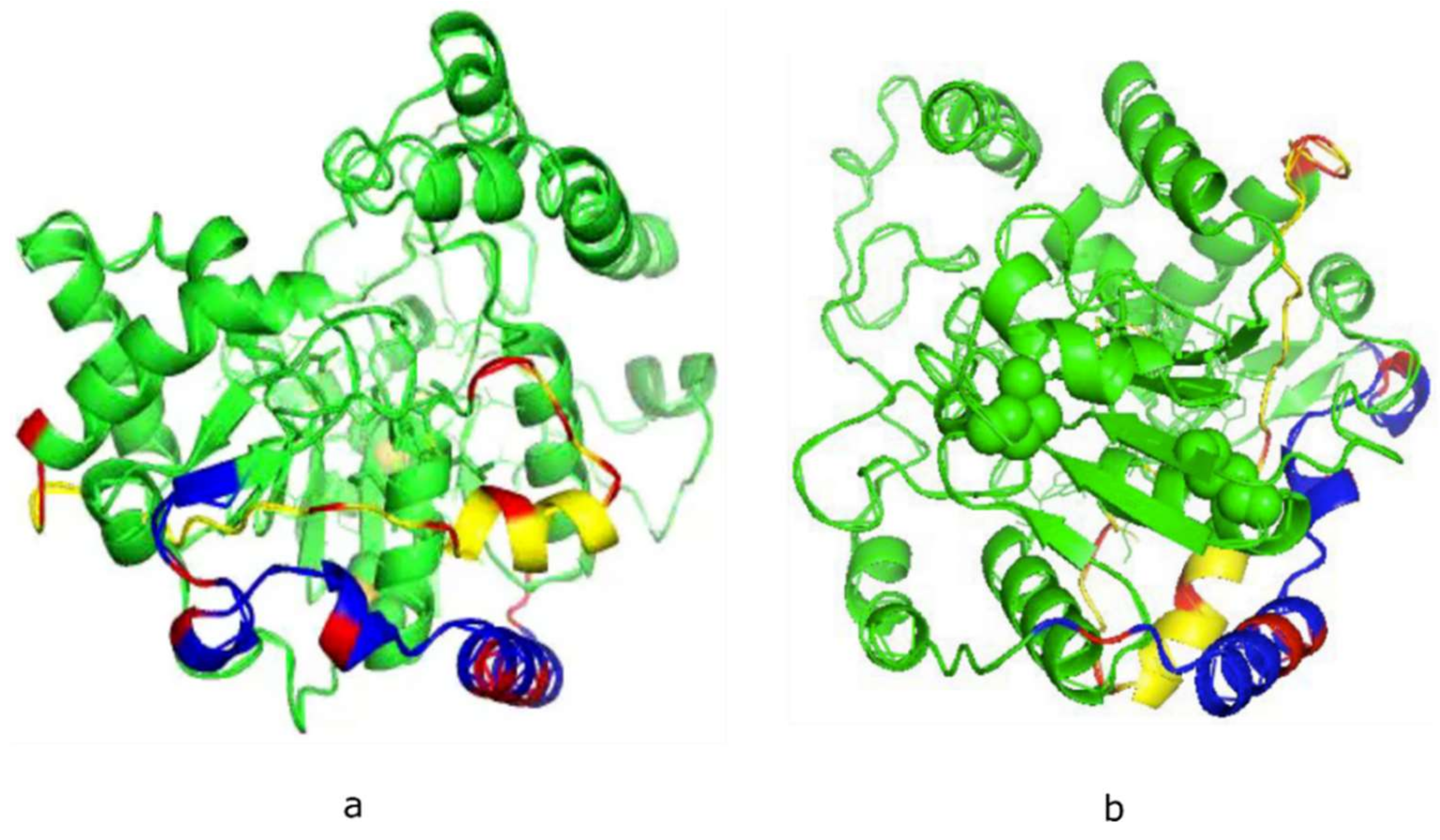
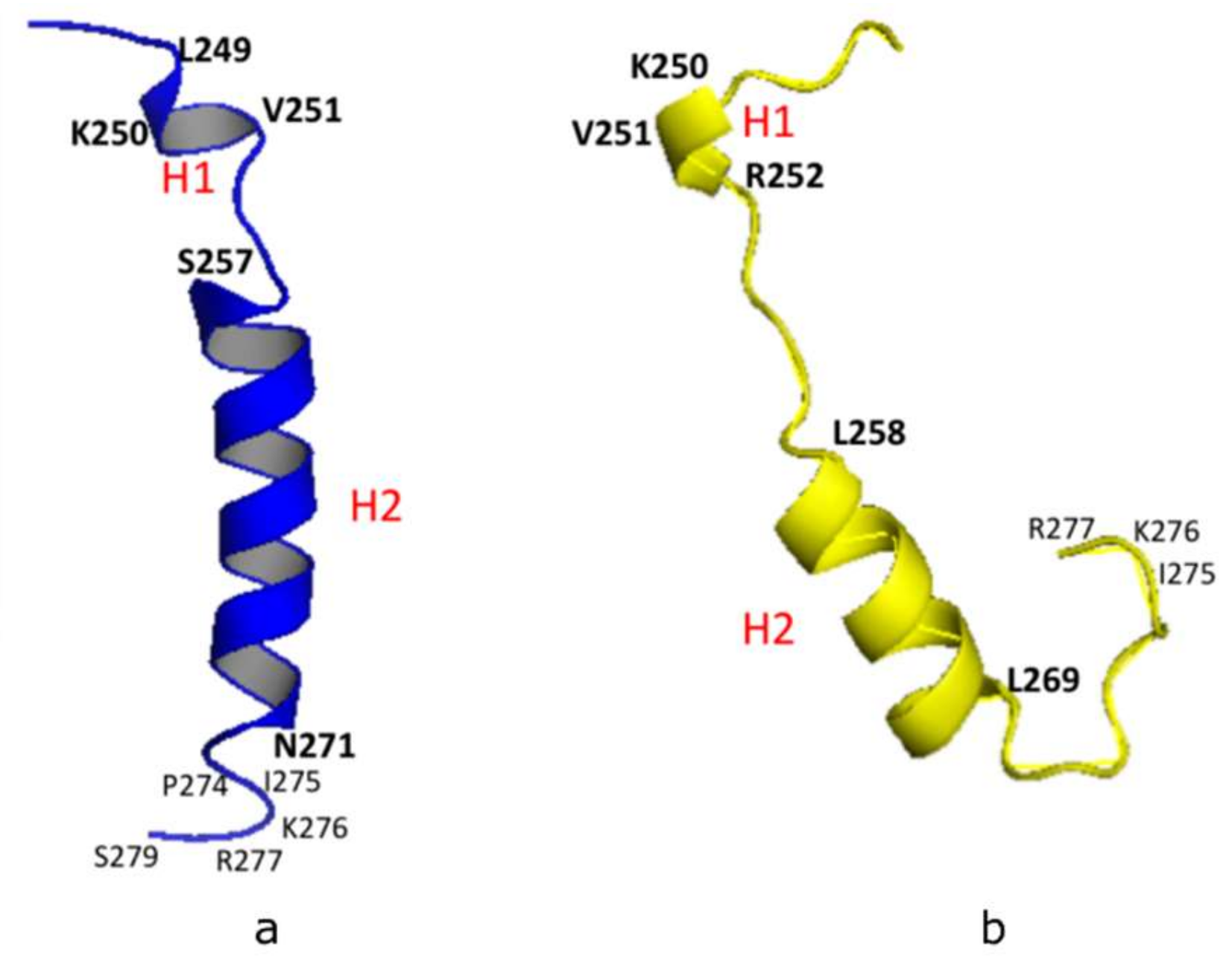
2.1. Structural Conformational Features of PSTD in ST8Sia IV
2.2. Molecular Interaction between CMP-Sia and PSTD in ST8Sia IV
2.3. Specificity of the Interaction of PSTD-Sia, PSTD- DP3, PST- CMP-Sia and PSTD-polySia
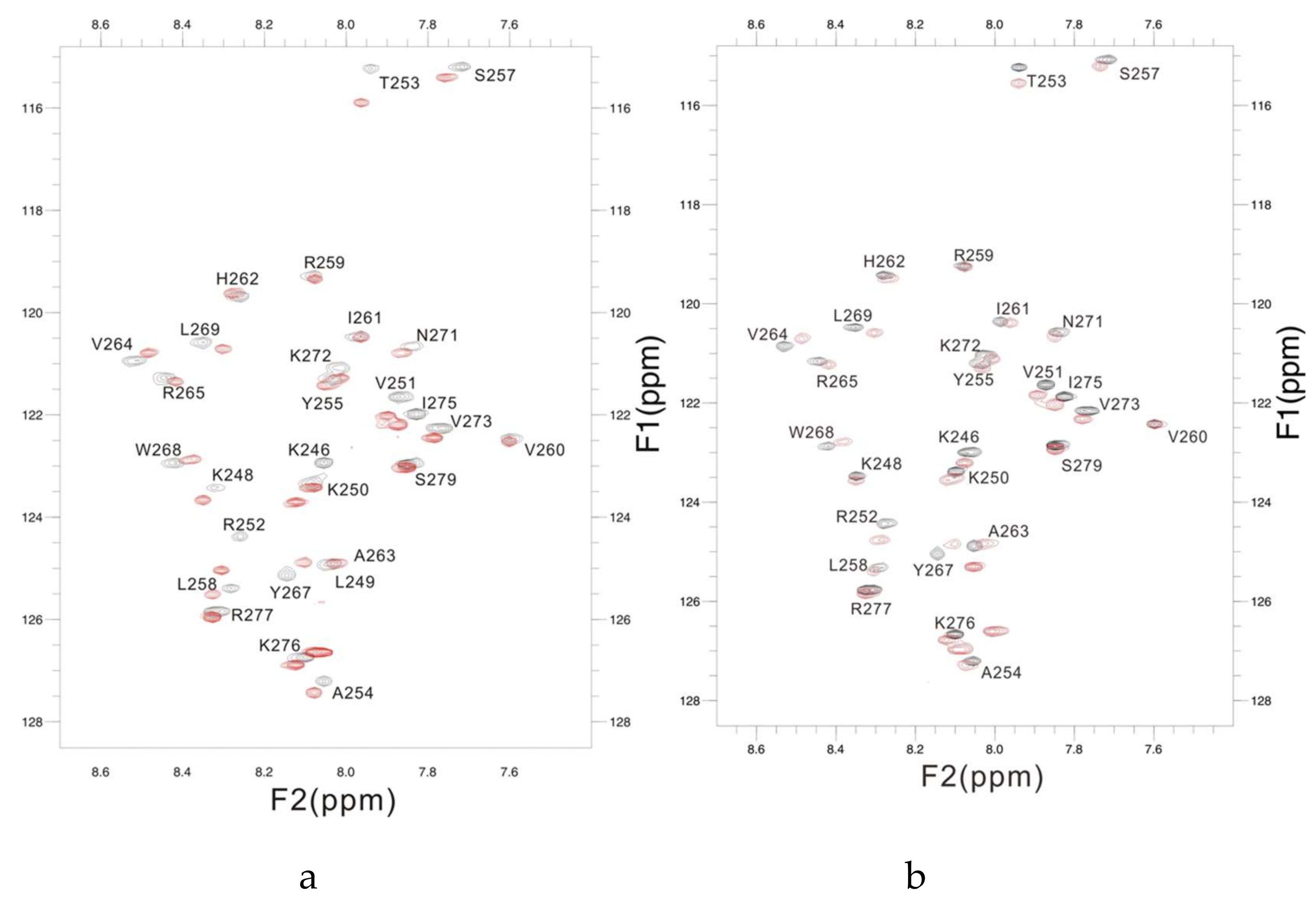
2.4. Interaction between PSTD and PolySia
- 1.
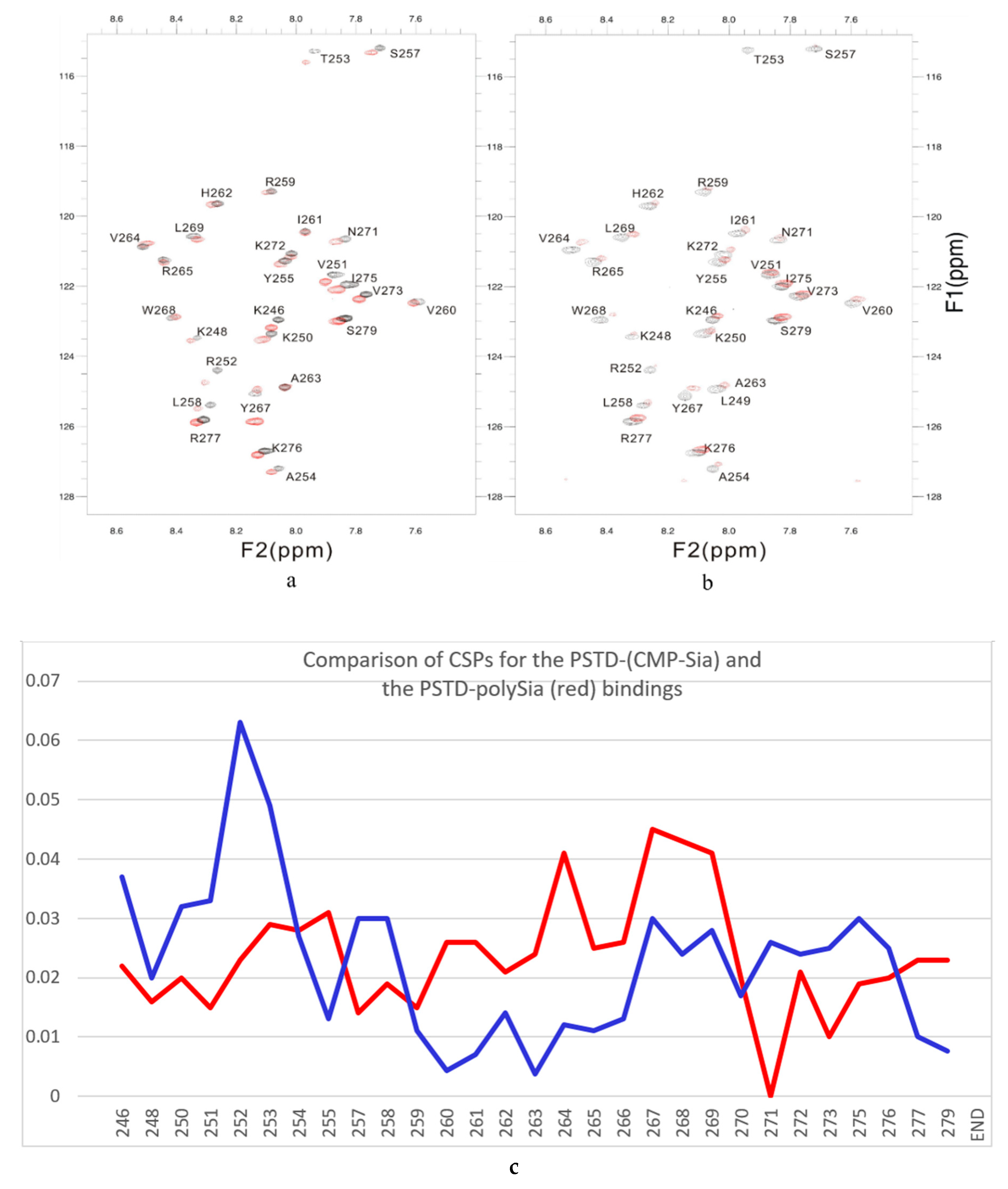
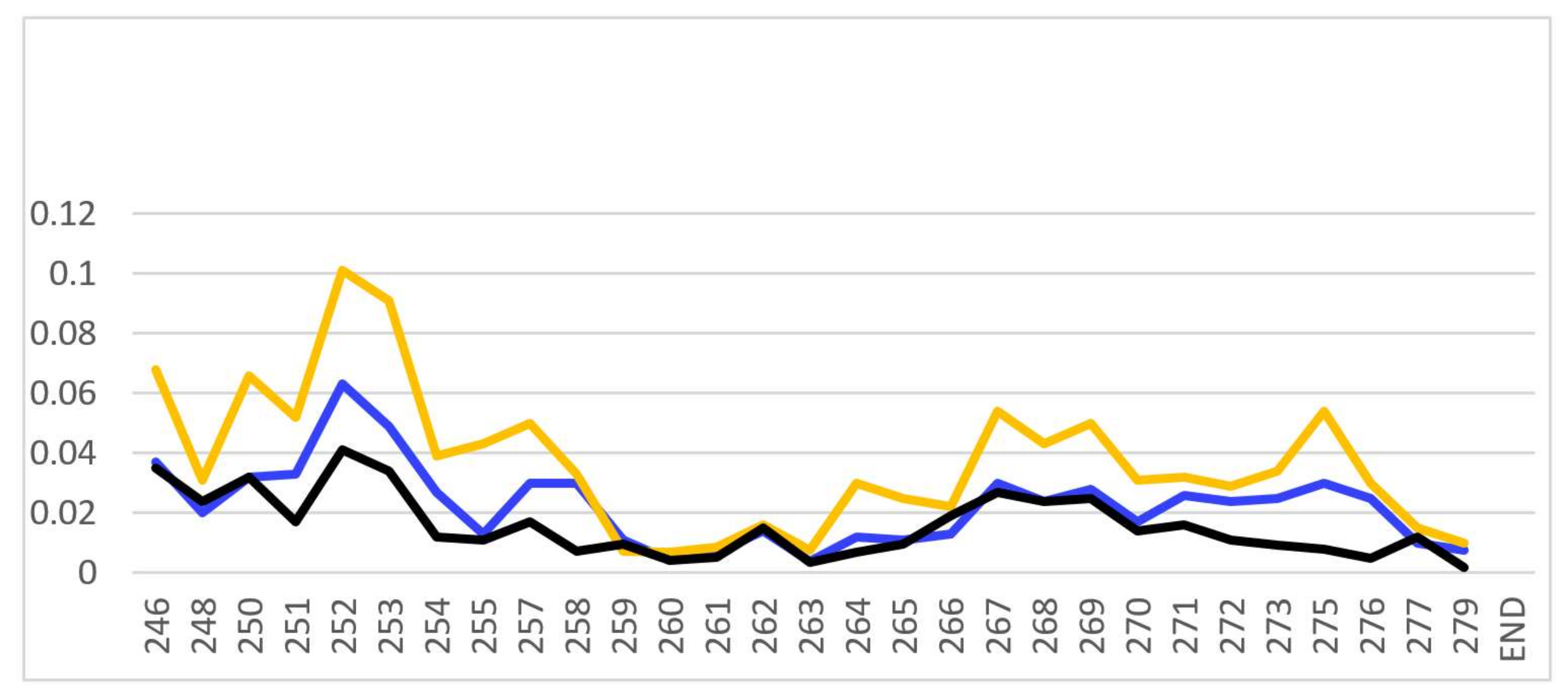
- The large chemical-shift perturbations (CSP>0.02) are most prominent in the N-terminus amino acids K248 to A254, for the PSTD-polySia interaction, and are less than for the interactions between PSTD-CMP-Sia, PSTD-Sia and PSTD-DP3 (Figure A2). The CSP values for amino acids V260-T270 are also much greater for the interaction between PSTD-polySia than for the interactions between PSTD-CMP-Sia and PSTD-DP 3 (Table 1; Figure 4c). This finding is consistent with a conformational change in the long H2 helix in PSTD, following its interaction with polySia.
- 2.
- These findings also show that the CSP values for the PSTD-polySia interaction in the C-terminus region (from 270 to S279) show small chemical shift perturbations (CSP < 0.02), which are less than for the interaction between PSTD-CMP-Sia and PSTD-Sia (Figure 4c, Figure A2). These results indicate that the predominantly small CSP values are located in the non-structural domains, while the larger CSP values are located on the long H2 helix for the interaction between PSTD-polySia (Figure 4c).
- 3.
- The amide cross-peak intensities for the 20 amino acid PSTD peptide were significantly decreased after interaction with polySia. The binding interaction between PSTD and polySia was characterized by broadening NMR signals that appeared in most amino acid residues except V251, V273, I275, K276, and R277 (Figure 4b and Table 1). This suggests a very slow conformational exchange or slow motion during the PSTD-polySia interaction. It is reasonable to expect that a decrease in the rate of tumbling occurs when a significantly larger ligand, such as the polydisperse polySia chains are bound to a protein or peptide [57,58,59,60,61]. The polySia-PSTD interaction is particularly evident, as seen in both the large chemical-shift perturbations, and the significant decrease in cross-peak intensity of amides that are observed in the 4 amino acid residues, V264, Y267, W268 and L269, consistent with the occurrence of both a conformational change and a slower motion in the C-terminus of the long H2 helix.
2.5. Functional Importance of the Degree of Polymerization (DP) of α2,8-linked PolySia Chains Covalently Modifying NCAM Proteins Related to Present Studies
3. Materials and Methods
3.1. Preparations of Samples for NMR Studies
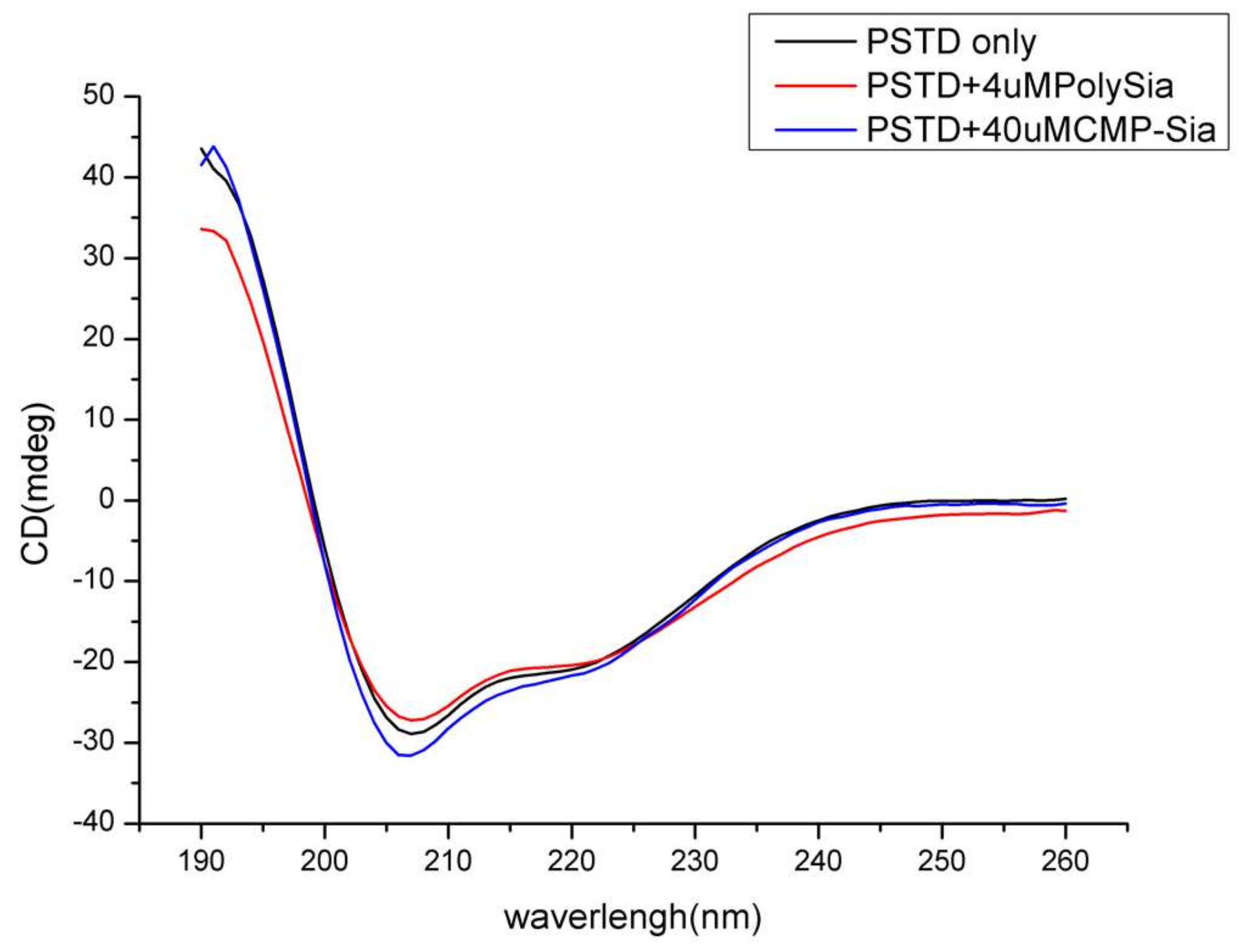
3.2. Circular Dichroism (CD) Spectroscopic Studies
3.3. NMR Spectroscopy
3.4. Structural Calculation and Analysis of the PSTD from ST8Sia IV
3.5. Structural Predictions of ST8Sia II and ST8Sia IV
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| polySia | α2,8-linked polysialic acid |
| Sia | sialic acid (Neu5Ac; N-acetylneuraminic acid) |
| DP3 | TriSia |
| CMP-Sia | cytidine monophosphate-sialic acid |
| NCAM | neural cell adhesion molecule proteins |
| polySTs | polysialyltransferases (ST8Sia II (STX) & ST8Sia IV (PST) |
| PSTD | polysialyltransferase domain |
| PBR | polybasic region |
| DP | degree of polymerization |
| E. coli | Escherichia coli |
Appendix A


References
- Lepers, A.H.; Petit, D.; Mollicone, R.; Delannoy, P.; Petit, J.M.; Oriol, R. Evolutionary history of the alpha2,8-sialyltransferase (ST8Sia) gene family: Tandem duplications in early deuterostomes explain most of the diversity found in the vertebrate ST8Sia genes. BMC Evol. Biol. 2008, 8, 258. [Google Scholar]
- Jeanneau, C.; Chazalet, V.; Auge, C.; Soumpasis, D.M.; Harduin-Lepers, A.; Delannoy, P.; Imberty, A.; Breton, C. Structure-function analysis of the human sialyltransferase ST3Gal I: Role of N-glycosylation and a novel conserved sialylmotif. J. Biol. Chem. 2004, 279, 13461–13468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Kurata, K.; Kojima, N.; Kurosawa, N.; Ohta, S.; Hanai, N.; Tsuji, S.; Nishi, T. Expression cloning of a GM3-specific alpha-2,8-sialyltransferase (GD3 synthase). J. Biol. Chem. 1994, 269, 15950–15956. [Google Scholar]
- Nakayama, J.; Fukuda, M.N.; Hirabayashi, Y.; Kanamori, A.; Sasaki, K.; Nishi, T.; Fukuda, M. Expression cloning of a human GT3 synthase. GD3 and GT3 are synthesized by a single enzyme. J. Biol. Chem. 1996, 271, 3684–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.; Xie, B.; Wang, B.; Troy, F.A., II. LC-MS/MS Glycomic Analyses of Free and Conjugated Forms of the Sialic Acids, Neu5Ac, Neu5Gc and KDN in Human Throat Cancers. Glycobiology 2015, 25, 1362–1374. [Google Scholar]
- Chen, Y.; Ren, H.; Zhang, N.; Troy, F.A., II; Wang, B. Biochemical Characterization and Analyses of Polysialic Acid-Associated Carrier Proteins and Genes in Piglets During Neonatal Brain Development. ChemBioChem 2017, 18, 1270–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, P.; Fang, W.; Chen, Y.; Zhang, N.; Qiao, Z.; Troy, F.A., II; Wang, B. Molecular Mechanisms Underlying How Sialyllactose Intervention Promotes Intestinal Maturity by Up-regulating GDNF Through a CREB-dependent Pathway in Neonatal Piglets. J. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Gluer, S.; Schelp, C.; Gerardy-Schahn, R.; von Schweinitz, D. Polysialylated neural cell adhesion molecule as a marker for differential diagnosis in pediatric tumors. J. Pediatr. Surg. 1998, 33, 1516. [Google Scholar] [CrossRef]
- Angata, K.; Suzuki, M.; McAuliffe, J.; Ding, Y.; Hindsgaul, O.; Fukuda, M. Differential biosynthesis of polysialic acid on neural cell adhesion molecule (NCAM) and oligosaccharide acceptors by three distinct alpha 2,8-sialyltransferases, ST8Sia IV (PST), ST8Sia II (STX), and ST8Sia III. J. Biol. Chem. 2000, 275, 18594–18601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashima1, S.; Matsumoto, T.; Tsujimoto, M.; Tsuji, S. Effects of amino acid substitutions in the sialylmotifs on molecular expression and enzymatic activities of α2,8-sialyltransferases ST8Sia-I and ST8Sia-VI. Glycobiology 2013, 23, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Petit, D.; Teppa, E.; Mir, A.M.; Vicogne, D.; Thisse, C.; Thisse, B.; Filloux, C.; Harduin-Lepers, A. Integrative View of α2,3-Sialyltransferases (ST3Gal) Molecular and Functional Evolution in Deuterostomes: Significance of Lineage-Specific Losses. Mol. Biol. Evol. 2015, 32, 906–927. [Google Scholar] [CrossRef] [Green Version]
- Harduin-Lepers, A. Comprehensive analysis of sialyltransferases in vertebrate genomes. Glycobiol Insights 2010, 2, 29–61. [Google Scholar] [CrossRef]
- Troy, F.A., II. Polysialylation: From bacteria to brains. Glycobiology 1992, 2, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Kauskot, A.; Pascreau, T.; Adam, F.; Bruneel, A.; Reperant, C.; Lourenco-Rodrigues, M.D.; Rosa, J.P.; Petermann, R.; Maurey, H.; Auditeau, C.; et al. A mutation in the gene coding for the sialic acid transporter SLC35A1 is required for platelet life span but not proplatelet formation. Haematologica 2018, 103, e613–e617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.L.; Newstead, S. Gateway to the Golgi:molecular mechanisms of nucleotide sugar transporters. Curr. Opin. Struct. Biol. 2019, 57, 127–134. [Google Scholar] [CrossRef]
- Parker, J.L.; Newstead, S. Structural basis of nucleotide sugar transport across the Golgi membrane. Nature 2017, 551, 521–524. [Google Scholar] [CrossRef] [Green Version]
- Eckhardt, M.; Gotza, B.; Gerardy-Schahn, R. Mutants of the CMP- sialic acid transporter causing the Lec2 phenotype. J. Biol. Chem. 1998, 273, 20189–20195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Ishida, N.; Kawakita, M. Substrate recognition by nucleotide sugar transporters: Further characterization of substrate recognition regions by analyses of UDP-galactose/CMP-sialic acid transporter chimeras and biochemical analysis of the substrate specificity of parental and chimeric transporters. J. Biol. Chem. 2003, 278, 22887–22893. [Google Scholar] [PubMed] [Green Version]
- Aoki, K.; Ishida, N.; Kawakita, M. Substrate recognition by UDP- galactose and CMP-sialic acid transporters. Different sets of transmembrane helices are utilized for the specific recognition of UDP-galactose and CMP-sialic acid. J. Biol. Chem. 2001, 276, 21555–21561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, A. The Beginnings of Sialic Acid. Biology of the Sialic Acids; Springer: Berlin, Germany, 1995; pp. 1–5. [Google Scholar]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Freeze, H.H.; Stanley, P.; Bertozzi, C.R.; Hart, G.W.; Etzler, M.E. Essentials of Glycobiology, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; pp. 199–216. [Google Scholar]
- McCoy, R.D.; Vimr, E.R.; Troy, F.A., II. CMP-NeuNAc:Poly-α,8-Sialosyl Sialyltransferase and the Biosynthesis of Polysialosyl Units in Neural Cell Adhesion Molecules. J. Biol. Chem. 1985, 260, 12695–12699. [Google Scholar]
- Drake, P.; Nathan, J.; Stock, C.; Chang, P.; Muench, M.; Nakata, D.; Reader, R.; Gip, P.; Golden, K.; Weinhold, B.; et al. The Highly Restricted Glycan, Polysialic Acid, is Differentially Expressed on Human and Murine Leukocytes and Modulated Immune Response. J. Immunol. 2008, 181, 6850–6858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevigny, M.B.; Ye, J.; Kitazume-Kawaguchi, S.; Troy, F.A., II. Developmental Expression and Characterization of the α2,8 Polysialyltransferase Activity in Embryonic Chick Brain. Glycobiology 1998, 8, 857–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troy, F.A., II. Polysialic Acid in Molecular Medicine. Encycl. Biol. Chem. 2004, 3, 407–414. [Google Scholar]
- Zhou, G.P.; Huang, R.B.; Troy, F.A., II. 3D Structural Conformation and Functional Domains of Polysialyltransferase ST8Sia IV Required for Polysialylation of Neural Cell Adhesion Molecules. Protein Pept. Lett. 2015, 22, 137–148. [Google Scholar] [CrossRef]
- Huang, R.B.; Chen, D.; Liao, S.M.; Lu, B.; Wang, Q.Y.; Xie, N.Z.; Troy, F.A., II; Zhou, G.P. The Intrinsic Relationship Between Structure and Function of the Sialyltransferase ST8Sia Family Members. Curr. Top. Med. Chem. 2017, 17, 2465–2475. [Google Scholar] [CrossRef]
- Nakata, D.; Zhang, L.; Troy, F.A. Molecular basis for polysialylation: A novel polybasic polysialyltransferase domain (PSTD) of 32 amino acids unique to the α2,8-polysialyltransferases is essential for polysialylation. Glycoconj. J. 2006, 23, 423–436. [Google Scholar] [CrossRef]
- Foley, D.A.; Swartzentruber, K.G.; Colley, K.J. Identification of Sequences in the Polysialyltransferases ST8Sia II and ST8Sia IV That Are Required for the Protein-specific Polysialylation of the Neural Cell Adhesion Molecule, NCAM. J. Biol. Chem. 2009, 204, 15505–15516. [Google Scholar] [CrossRef] [Green Version]
- Bhide, G.P.; Prehna, G.; Ramirez, B.E.; Colley, K.J. The Polybasic Region of the Polysialyltransferase ST8Sia-IV Binds Directly to the Neural Cell Adhesion Molecule, NCAM. Biochemistry 2017, 56, 1504. [Google Scholar] [CrossRef] [Green Version]
- Kelley, L.A.; Sternberg, M.J.E. Protein structure prediction on web: A case study using Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Werneburg, S.; Muhlenhoff, M.; Stangel, M.; Hildebrandt, H. Polysialic acid on SynCAM 1 in NG2 cells and on neuropilin-2 in microglia is confined to intracellular pools that are rapidly depleted upon stimulation. GLIA 2015, 63, 1240–1255. [Google Scholar] [CrossRef]
- Roger, S.; Heike, S.; Konrad, S. Gangliosides in Health and Disease. In Progress in Molecular Biology and Translational Science; Schnaar, R.L., Lopez, P.H.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–462. [Google Scholar]
- Prem Raj, B.J.; Krishna Mohan, P.; Krishna Mohan, S.; Krishna, R. Characterizing protein-glycosaminoglycan interactions using solution NMR Spectroscopy. Methods Mol. Biol. 2015, 1229, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Vaynberg, J.; Qin, J. Weak protein—Protein interactions as probed by NMR spectroscopy. Trends Biotechnol. 2006, 24, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Zhou, G.P.; Kupferman, J.; Surks, H.K.; Christensen, E.N.; Chou, J.J.; Mendelsohn, M.E.; Rigby, A.C. Probing the interaction between the coiled coil leucine zipper of cGMP-dependent protein kinase I alpha and the C terminus of the myosin binding subunit of the myosin light chain phosphatase. J. Biol. Chem. 2008, 283, 32860–32869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wüthrich, K. NMR of Proteins and Nucleic Acids; Wiley: Hoboken, NJ, USA, 1996; ISBN: 978-0-471-82893-8; Available online: https://www.wiley.com/en-us/NMR+of+Proteins+and+Nucleic+Acids-p-9780471828938 (accessed on 15 January 2020).
- Zhou, G.P.; Troy, F.A., II. NMR studies on how the binding complex of polyisoprenol recognition sequence peptides and polyisoprenols can modulate membrane structure. Curr. Protein Pept. 2005, 6, 399–411. [Google Scholar] [CrossRef]
- Oxenoid, K.; Dong, Y.; Cao, C.; Cui, T.; Sancak, Y.; Markhard, A.L.; Grabarek, Z.; Kong, L.; Liu, Z.; Ouyang, B.; et al. Architecture of the mitochondrial calcium uniporter. Nature 2016, 533, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Van Horn, W.D.; Kim, H.J.; Ellis, C.D.; Hadziselimovic, A.; Sulistijo, E.S.; Karra, M.D.; Tian, C.; Sonnichsen, F.D.; Sanders, C.R. Solution nuclear magnetic resonance structure of membrane-integral diacylglycerol kinase. Science 2009, 324, 1726–1729. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.; Tian, C.; Sonnichsen, F.D.; Smith, J.A.; Meiler, J.; George, A.L., Jr.; Vanoye, C.G.; Kim, H.J.; Sanders, C.R. Structure of KCNE1 and implications for how it modulates the KCNQ1 potassium channel. Biochemistry 2008, 47, 7999–8006. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.P.; Surks, H.K.; Schnell, J.R.; Chou, J.J.; Mendelsohn, M.E.; Rigby, A.C. The Three-Dimensional Structure of the cGMP-Dependent Protein Kinase I - α Leucine Zipper Domain and Its Interaction with the Myosin Binding Subunit. Blood 2004, 104, 3539. [Google Scholar] [CrossRef]
- Chou, J.J.; Li, S.; Klee, C.B.; Bax, A. Solution structure of Ca(2+)-calmodulin reveals flexible hand-like properties of its domains. Nat. Struct. Biol. 2001, 8, 990–997. [Google Scholar] [CrossRef]
- Chou, J.J.; Li, H.; Salvesen, G.S.; Yuan, J.; Wagner, G. Solution structure of BID, an intracellular amplifier of apoptotic signaling. Cell 1999, 96, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Fu, T.M.; Cruz, A.C.; Sengupta, P.; Thomas, S.K.; Wang, S.; Siegel, R.M.; Wu, H.; Chou, J.J. Structural Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 Death Receptor. Mol. Cell 2016, 61, 602–613. [Google Scholar] [CrossRef] [Green Version]
- Pielak, R.M.; Schnell, J.R.; Chou, J.J. Mechanism of drug inhibition and drug resistance of influenza A M2 channel. Proc. Natl. Acad. Sci. USA 2009, 106, 7379–7384. [Google Scholar] [CrossRef] [Green Version]
- Dev, J.; Park, D.; Fu, Q.; Chen, J.; Ha, H.J.; Ghantous, F.; Herrmann, T.; Chang, W.; Liu, Z.; Frey, G.; et al. Structural basis for membrane anchoring of HIV-1 envelope spike. Science 2016, 353, 172–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, J.J.; Matsuo, H.; Duan, H.; Wagner, G. Solution structure of the RAIDD CARD and model for CARD/CARD interaction in caspase-2 and caspase-9 recruitment. Cell 1998, 94, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Gagnon, E.; Call, M.E.; Schnell, J.R.; Schwieters, C.D.; Carman, C.V.; Chou, J.J.; Wucherpfennig, K.W. Regulation of T cell receptor activation by dynamic membrane binding of the CD3epsilon cytoplasmic tyrosine-based motif. Cell 2008, 135, 702–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, E.; Xu, C.; Yang, W.; Chu, H.H.; Call, M.E.; Chou, J.J.; Wucherpfennig, K.W. Response multilayered control of T cell receptor phosphorylation. Cell 2010, 142, 669–671. [Google Scholar] [CrossRef] [Green Version]
- Pielak, R.M.; Chou, J.J. Flu channel drug resistance: A tale of two sites. Protein Cell 2010, 1, 246–258. [Google Scholar]
- Berardi, M.J.; Chou, J.J. Fatty acid flippase activity of UCP2 is essential for its proton transport in mitochondria. Cell Metab. 2014, 20, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Mantsyzov, A.B.; Shen, Y.; Lee, J.H.; Hummer, G.; Bax, A. MERA: A webserver for evaluating backbone torsion angle distributions in dynamic and disordered proteins from NMR data. J. Biomol. NMR 2015, 63, 85–95. [Google Scholar] [CrossRef]
- Shen, Y.; Bax, A. Protein structural information derived from NMR chemical shift with the neural network program TALOS-N. Methods Mol. Biol. 2015, 1260, 17–32. [Google Scholar]
- Bjorndahl, T.C.; Zhou, G.P.; Liu, X.H.; Pineiro, R.P.; Semenchenko, V.; Saleem, F.; Acharya, S.; Bujold, A.; Sobsey, C.A.; Wishart, D.S. Detailed Biophysical Characterization of the Acid-Induced PrPc to PrPβConversion Process. Biochemistry 2011, 50, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Nakata, D.; Troy, F.A., II. Degree of Polymerization (DP) of Polysialic Acid (PolySia) on Neural Cell Adhesion Molecules (N-CAMs): Development and Application of a New Strategy to Accurately Determine the DP of PolySia Chains on NCAMs. J. Biol. Chem. 2006, 280, 38305–38316. [Google Scholar] [CrossRef] [Green Version]
- Mike, P.; Williamson, M.P. Using chemical shift perturbation to characterise ligand binding. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 73, 1–16. [Google Scholar]
- Rao, B.D.N. Nuclear magnetic resonance line-shape analysis and determination of exchange rates. Meth. Enzymol. 1989, 176, 279–311. [Google Scholar] [PubMed]
- Sandstrm, J. Dynamic NMR Spectroscopy; Academic Press: London, UK, 1982; X, 226 S., geb. [Google Scholar]
- Peng, L.X.; Liu, X.H.; Lu, B.; Liao, S.M.; Huang, J.M.; Zhou, F.; Chen, D.; Troy, F.A., II; Huang, R.B.; Zhou, G.P. The Inhibition of polysialyltransferase ST8SiaIV through heparin binding to polysialyltransferase domain (PSTD). Med. Chem. 2019, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Volkers, G.; Worrall, L.J.; Kwan, D.H.; Yu, C.C.; Baumann, L.; Lameignere, E.; Wasney, G.A.; Scott, N.E.; Wakarchuk, W.; Foster, L.J.; et al. Structure of human ST8Sia III sialyltransferase provides insight into cell-surface polysialylation. Nat. Struct. Mol. Biol. 2015, 22, 627–635. [Google Scholar]
- Castilho, A.; Pabst, M.; Leonard, R.; Veit, C.; Altmann, F.; Mach, L.; Glossl, J.; Strasser, R.; Steinkellner, H. Construction of a Functional CMP-Sialic Acid Biosynthesis Pathway in Arabidopsis. Plant Physiol. 2008, 147, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Nagae, M.; Ikeda, A.; Hane, M.; Hanashima, S.; Kitajima, K.; Sato, C.; Yamaguchi, Y. Crystal structure of anti-polysialic acid antibody single chain Fv fragment complexed with octasialic acid: Insight into the binding preference for polysialic acid. J. Biol. Chem. 2013, 288, 33784–33796. [Google Scholar] [CrossRef] [Green Version]
- Ono, S.; Hane, M.; Kitajima, K.; Sato, C. Novel regulation of fibroblast growth factor 2 (FGF2)-mediated cell growth by polysialic acid. J. Biol. Chem. 2012, 287, 3710–3722. [Google Scholar] [CrossRef] [Green Version]
- Kanato, Y.; Kitajima, K.; Sato, C. Direct binding of polysialic acid to a brain-derived neurotrophic factor depends on the degree of polymerization. Glycobiology 2008, 18, 1044–1053. [Google Scholar] [CrossRef] [Green Version]
- Kuhnle, A.; Veelken, R.; Galuska, C.E.; Saftenberger, M.; Verleih, M.; Schuppe, H.C.; Rudloff, S.; Kunz, C.; Galuska, S.P. Polysialic acid interacts with lactoferrin and supports its activity to inhibit the release of neutrophil extracellular traps. Carbohydr. Polym. 2019, 208, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Zlatina, K.; Lütteke, T.; Galuska, S.P. Individual Impact of Distinct Polysialic Acid Chain Lengths on the Cytotoxicity of Histone H1, H2A, H2B, H3 and H4. Polymers 2017, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Galuska, C.E.; Dambon, J.A.; Kühnle, A.; Bornhöfft, K.F.; Prem, G.; Zlatina, K.; Lütteke, T.; Galuska, S.P. Artificial Polysialic Acid Chains as Sialidase-Resistant Molecular-Anchors to Accumulate Particles on Neutrophil Extracellular Traps. Front. Immunol. 2017, 8, 1229. [Google Scholar] [CrossRef] [Green Version]
- Haselhorst, T.; Stummeyer, K.; Muhlenhoff, M.; Schaper, W.; Gerardy-Schahn, R.; von Itzstein, M. Endosialidase NF appears to bind polySia DP5 in a helical conformation. Chembiochem 2006, 7, 1875–1877. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic acids in the brain: Gangliosides and polysialic Acid in nervous system development, stability, disease, and regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.P. Current Advances of Drug Target Research in Medicinal Chemistry. CTMC 2019, 19, 2269–2270. [Google Scholar] [CrossRef] [PubMed]
- Schnell, J.R.; Zhou, G.P.; Zweckstetter, M.; Rigby, A.C.; Chou, J.J. Rapid and accurate structure determination of coiled-coil domains using NMR dipolar couplings: Application to cGMP-dependent protein kinase Iα. Protein Sci. 2005, 14, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Krajewski, M.; Ozdowy, P.; D’Silva, L.; Rothweiler, U.; Holak, T.A. NMR indicates that the small molecule RITA does not block p53-MDM2 binding in vitro. Nat. Med. 2005, 11, 1135–1136. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.P.; Troy, F.A., II. NMR study of the preferred membrane orientation of polyisoprenols (dolichol) and the impact of their complex with polyisoprenyl recognition sequence peptides on membrane structure. Glycobiology 2005, 15, 347–359. [Google Scholar] [CrossRef] [Green Version]
- Close, B.E.; Colley, K.J. In vivo autopolysialylation and localization of the polysialyltransferases PST and STX. J. Biol. Chem. 1998, 273, 34586–34593. [Google Scholar] [CrossRef] [Green Version]
- Bhide, G.P.; Zapater, J.L.; Colley, K.J. Autopolysialylation of polysialyltransferases is required for polysialylation and polysialic acid chain elongation on select glycoprotein substrates. J. Biol. Chem. 2018, 293, 701–716. [Google Scholar] [CrossRef] [PubMed]
- Werneburg, S.; Buettner, F.F.; Erben, L.; Mathews, M.; Neumann, H.; Muhlenhoff, M.; Hildebrandt, H. Polysialylation and lipopolysaccharide-induced shedding of E-selectin ligand-1 and neuropilin-2 by microglia and THP-1 macrophages. Glia 2016, 64, 1314–1330. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.P.; Liao, S.M.; Chen, D.; Huang, R.B. The Cooperative Effect between Polybasic Region (PBR) and Polysialyltransferase Domain (PSTD) within Tumor-Target Polysialyltranseferase ST8Sia II. CTMC 2019, 19. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand Interacting with PSTD | Amino Acid Residues Displaying Large Chemical Shift Perturbation (CSP > 0.02) | Amino Acid Residues Displaying the Largest CSP Values | Amino Acid Residues Exhibiting Significant Decrease in Peak Intensities |
|---|---|---|---|
| polySia | R252-P256 V260-T270 | A263-T270 | 20 residues: K246, K248, K250, R252, T253, A254, Y255, S257, L258, R259, I261, H262, A263, V264, R265, Y267, W268, L269, N271, K272. |
| CMP-Sia | K246-A254, P256-L258, Y267-L269, N271-R277 | V251-A254 | None |
| Sia | K246-L258, V264-R277 | V251-A254 | None |
| DP 3 | K246-T253, G266-T270 | V251-A254 | None |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, S.-M.; Lu, B.; Liu, X.-H.; Lu, Z.-L.; Liang, S.-J.; Chen, D.; Troy, F.A., II; Huang, R.-B.; Zhou, G.-P. Molecular Interactions of the Polysialytransferase Domain (PSTD) in ST8Sia IV with CMP-Sialic Acid and Polysialic Acid Required for Polysialylation of the Neural Cell Adhesion Molecule Proteins: An NMR Study. Int. J. Mol. Sci. 2020, 21, 1590. https://doi.org/10.3390/ijms21051590
Liao S-M, Lu B, Liu X-H, Lu Z-L, Liang S-J, Chen D, Troy FA II, Huang R-B, Zhou G-P. Molecular Interactions of the Polysialytransferase Domain (PSTD) in ST8Sia IV with CMP-Sialic Acid and Polysialic Acid Required for Polysialylation of the Neural Cell Adhesion Molecule Proteins: An NMR Study. International Journal of Molecular Sciences. 2020; 21(5):1590. https://doi.org/10.3390/ijms21051590
Chicago/Turabian StyleLiao, Si-Ming, Bo Lu, Xue-Hui Liu, Zhi-Long Lu, Shi-Jie Liang, Dong Chen, Frederic A. Troy, II, Ri-Bo Huang, and Guo-Ping Zhou. 2020. "Molecular Interactions of the Polysialytransferase Domain (PSTD) in ST8Sia IV with CMP-Sialic Acid and Polysialic Acid Required for Polysialylation of the Neural Cell Adhesion Molecule Proteins: An NMR Study" International Journal of Molecular Sciences 21, no. 5: 1590. https://doi.org/10.3390/ijms21051590
APA StyleLiao, S. -M., Lu, B., Liu, X. -H., Lu, Z. -L., Liang, S. -J., Chen, D., Troy, F. A., II, Huang, R. -B., & Zhou, G. -P. (2020). Molecular Interactions of the Polysialytransferase Domain (PSTD) in ST8Sia IV with CMP-Sialic Acid and Polysialic Acid Required for Polysialylation of the Neural Cell Adhesion Molecule Proteins: An NMR Study. International Journal of Molecular Sciences, 21(5), 1590. https://doi.org/10.3390/ijms21051590