No Influence of Overweight/Obesity on Exercise Lipid Oxidation: A Systematic Review


Abstract
:1. Introduction
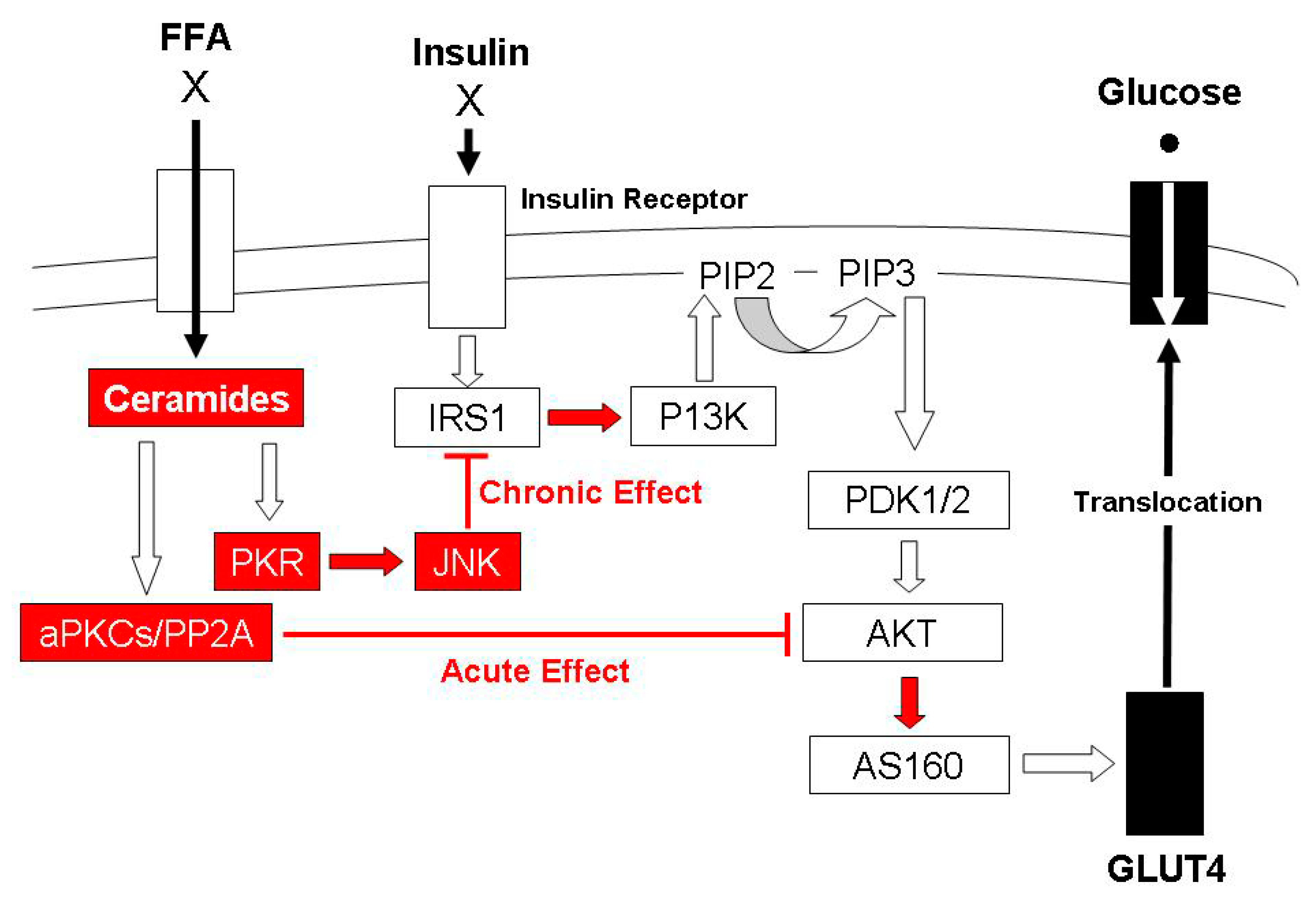
1.1. The Influence of Elevated Intramuscular Triglycerides on the Insulin Signal Transduction Pathway
1.2. The Cause of Increased Accumulation of Lipotoxic Metabolites with Overweight/Obesity
1.3. The Influence of Overweight/Obesity on Exercise Lipid Oxidation
2. Results
2.1. Methodological Characteristics of Included Studies
2.1.1. Stratification Criteria for Body Fatness
2.1.2. Dietary Control
2.1.3. Criterion Exercise Challenge
2.2. Conclusions from Included Studies Regarding the Influence of Overweight/Obesity on Exercise Lipid Use
2.3. Quality Assessment of Included Studies
3. Discussion
3.1. Rationale for and Evidence of Decreased Exercise Lipid Oxidation for Overweight/Obese Individuals
3.2. Rationale for and Evidence of Increased Exercise Lipid Oxidation for Overweight/Obese Individuals
3.3. Individual Characteristics that Might Influence Exercise Lipid Use Irrespective of Body Fatness
3.3.1. Sex
3.3.2. Race
3.3.3. Fat-Free Mass
3.3.4. Aerobic Fitness
3.3.5. Insulin Resistance
3.3.6. Menstrual Phase
3.3.7. Fat-Deposition Pattern
3.3.8. Acute and ‘Long-Term’ Dietary Habits
3.4. Exercise Variables that Might Influence Exercise Lipid Use Irrespective of Body Fatness
3.4.1. Mode
3.4.2. Duration
3.4.3. Intensity
4. Materials and Methods
4.1. Search Strategy
4.2. Inclusion and Exclusion Criteria
4.3. Data Extraction and Synthesis
4.4. Quality Assessment
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | African American |
| Akt | Protein kinase B |
| AM | Morning assessment |
| BM | Body mass |
| BMI | Body mass index |
| CRF | Cardiorepiratory fitness |
| CWR | Constant-work-rate exercise |
| DAG | Diacylglycerols |
| FATmax | Maximal rate of lipid oxidation |
| FFA | Free fatty acid |
| FFM | Fat-free mass |
| FQ | Food quotient |
| HR | Heart rate |
| IMTG | Intramuscular triglyceride |
| IR | Insulin resistance |
| IRS1 | Insulin receptor substrate 1 |
| LBO | Lower body obese |
| PKR | Protein kinase R |
| PPO | Peak power output |
| PTS | Preferred walk-run transition speed |
| RER | Respiratory exchange ratio |
| RQ | Respiratory quotient |
| Tlim | Limit of tolerance |
| T2D | Type 2 diabetes |
| VO2max | Maximal rate of oxygen consumption |
| VO2peak | Peak rate of oxygen consumption |
| VT | Ventilatory threshold |
| WHO | World Health Organization |
| WRmax(est) | Maximal work rate estimated according to prediction equations |
| NW | Normal weight |
| O | Overweight/obese |
| PM | Evening assessment |
| UBO | Upper body obese |
References
- World Health Organization. Obesity and Overweight; World Health Organization; Available online: https://www.who.int/bulletin/releases/NFM0715/en/ (accessed on 1 July 2015).
- Zimmet, P.; Alberti, K.G.M.M.; Shaw, J. Global and societal implication of the Diabetes Epidemic. Nature 2001, 14, 782–787. [Google Scholar] [CrossRef]
- Qatanani, M.; Lazar, M.A. Mechanisms of obesity-associated insulin resistance: Many choices on the menu. Genes Dev. 2007, 21, 1443–1455. [Google Scholar] [CrossRef] [Green Version]
- Moro, C.; Bajpeyi, S.; Smith, S.R. Determinants of intramyocellular triglyceride turnover: Implications for insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E203–E213. [Google Scholar] [CrossRef]
- Pan, D.A.; Lillioja, S.; Kriketos, A.D.; Milner, M.R.; Baur, L.A.; Bogardus, C.; Jenkins, A.B.; Storlien, L.H. Skeletal muscle triglyceride levels are inversely related to insulin action. Diabetes 1997, 46, 983–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodpaster, B.H.; He, J.; Watkins, S.; Kelley, D.E. Skeletal muscle lipid content and insulin resistance: Evidence for a paradox in endurance-trained athletes. J. Clin. Endocrinol. Metab. 2001, 86, 5755–5761. [Google Scholar] [CrossRef] [PubMed]
- Amati, F.; Dubé, J.J.; Alvarez-Carnero, E.; Edreira, M.M.; Chomentowski, P.; Coen, P.M.; Switzer, G.E.; Bickel, P.E.; Stefanovic-Racic, M.; Toledo, F.G.; et al. Skeletal muscle triglycerides, diacylglycerols, and ceramides in insulin resistance: Another paradox in endurance-trained athletes? Diabetes 2011, 60, 2588–2597. [Google Scholar] [CrossRef] [Green Version]
- Hage Hassan, R.; Pacheco de Sousa, A.C.; Mahfouz, R.; Hainault, I.; Blachnio-Zabielska, A.; Bourron, O.; Koskas, F.; Górski, J.; Ferré, P.; Foufelle, F.; et al. Sustained action of ceramide on the insulin signaling pathway in muscle cells: Implication of the double-stranded rna-activated protein kinase. J. Biol. Chem. 2016, 291, 3019–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasumov, T.; Solomon, T.P.; Hwang, C.; Huang, H.; Haus, J.M.; Zhang, R.; Kirwan, J.P. Improved insulin sensitivity after exercise training is linked to reduced plasma C14:0 ceramide in obesity and type 2 diabetes. Obesity 2015, 23, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Zierath, J.R. The path to insulin resistance: Paved with ceramides? Cell Metab. 2007, 5, 161–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kien, C.L.; Bunn, J.Y.; Poynter, M.E.; Stevens, R.; Bain, J.; Ikayeva, O.; Fukagawa, N.K.; Champagne, C.M.; Crain, K.I.; Koves, T.R.; et al. A lipidomics analysis of the relationship between dietary fatty acid composition and insulin sensitivity in young adults. Diabetes 2013, 62, 1054–1063. [Google Scholar] [CrossRef] [Green Version]
- Walls, S.M., Jr.; Attle, S.J.; Brulte, G.B.; Walls, M.L.; Finley, K.D.; Chatfield, D.A.; Herr, D.R.; Harris, G.L. Identification of sphingolipid metabolites that induce obesity via misregulation of appetite, caloric intake and fat storage in Drosophila. PLoS Genet. 2013, 9, e1003970. [Google Scholar] [CrossRef]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Hai Hoang, B.; Sanders, P.; Siddall, P.; Shang Kuo, M.; Perreault, L. Serum sphingolipids: Relationships to insulin sensitivity and changes with exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E398–E408. [Google Scholar] [CrossRef] [Green Version]
- Jansson, E.; Kaijser, L. Substrate utilization and enzymes in skeletal muscle of extremely endurance-trained men. J. Appl. Physiol. 1987, 62, 999–1005. [Google Scholar] [CrossRef]
- Coggan, A.R.; Raguso, C.A.; Gastaldelli, A.; Sidossis, L.S.; Yeckel, C.W. Fat metabolism during high-intensity exercise in endurance-trained and untrained men. Metabolism 2000, 49, 122–128. [Google Scholar] [CrossRef]
- Henriksson, J. Training induced adaptation of skeletal muscle and metabolism during submaximal exercise. J. Physiol. 1977, 270, 661–675. [Google Scholar] [CrossRef]
- Kiens, B.; Essen-Gustavsson, B.; Christensen, N.J.; Saltin, B. Skeletal muscle substrate utilization during submaximal exercise in man: Effect of endurance training. J. Physiol. 1993, 469, 459–478. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Katsiaras, A.; Kelley, D.E. Enhanced fat oxidation through physical activity is associated with improvements in insulin sensitivity in obesity. Diabetes 2003, 52, 2191–2197. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.E.; Goodpaster, B.; Wing, R.R.; Simoneau, J.A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am. J. Physiol. 1999, 277, E1130–E1141. [Google Scholar] [CrossRef]
- Galgani, J.E.; Moro, C.; Ravussin, E. Metabolic flexibility and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1009–E1017. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Hickner, R.C.; Cortright, R.L.; Dohm, G.L.; Houmard, J.A. Lipid oxidaiton is reduced in obese human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1039–E1044. [Google Scholar] [CrossRef] [Green Version]
- Simoneau, J.A.; Veerkamp, J.H.; Turcotte, L.P.; Kelley, D.E. Markers of capacity to utilize fatty acids in human skeletal muscle: Relation to insulin resistance and obesity and effects of weight loss. FASEB J. 1999, 13, 2051–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.; Harber, V. Lower excess postexercise oxygen consumption and altered growth hormone and cortisol responses to exercise in obese men. J. Clin. Endocrinol. Metab. 2006, 91, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, A.B.; Farrell, P.A.; Kalkhoff, R.K. Impaired plasma catecholamine response to submaximal treadmill exercise in obese women. Metabolism 1990, 39, 410–417. [Google Scholar] [CrossRef]
- Connacvher, A.A.; Bennet, W.M.; Jung, R.T.; Bier, D.M.; Smith, C.C.; Scrimgeour, C.M.; Rennie, M.J. Effect of adrenaline infusion on fatty acid and glucose turnover in lean and obese human subjects in the post-absorptive and fed states. Clin. Sci. 1991, 81, 635–644. [Google Scholar] [CrossRef]
- Thyfault, J.P.; Kraus, R.M.; Hickner, R.C.; Howell, A.W.; Wolfe, R.R.; Dohm, G.L. Impaired plasma fatty acid oxidation in extremely obese women. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E1076–E1081. [Google Scholar] [CrossRef] [Green Version]
- Goodpaster, B.H.; Wolfe, R.R.; Kelley, D.E. Effects of obesity on substrate utilization during exercise. Obes. Res. 2002, 10, 575–584. [Google Scholar] [CrossRef]
- Horowitz, J.F.; Klein, S. Oxidation of nonplasma fatty acids during exercise is increased in women with abdominal obesity. J. Appl. Physiol. 2000, 89, 2276–2282. [Google Scholar] [CrossRef]
- Braun, B.; Sharoff, C.; Chipkin, S.R.; Beaudoin, F. Effects of insulin resistance on substrate utilization during exercise in overweight women. J. Appl. Physiol. 2004, 97, 991–997. [Google Scholar] [CrossRef] [Green Version]
- Gregory, S.; Wood, R.; Matthews, T.; VanLangen, D.; Sawyer, J.; Headley, S. Substrate utilization is influenced by acute dietary carbohydrate intake in active, healthy females. J. Sports Sci. Med. 2011, 10, 59–65. [Google Scholar]
- Helge, J.W.; Watt, P.W.; Richter, E.A.; Rennie, M.J.; Kiens, B. Fat utilization during exercise: Adaptation to a fat-rich diet increases utilization of plasma fatty acids and very low density lipoprotein-triaglycerol in humans. J. Physiol. 2001, 537, 1009–1020. [Google Scholar] [CrossRef]
- Van Loon, L.J.; Koopman, R.; Stegen, J.H.; Wagenmakers, A.J.; Keizer, H.A.; Saris, W.H. Intramyocellular lipids form an important substrate source during moderate intensity exercise in endurance-trained males in a fasted state. J. Physiol. 2003, 553, 611–625. [Google Scholar] [CrossRef] [Green Version]
- Guesbeck, N.R.; Hickey, M.S.; MacDonald, K.G.; Pories, W.J.; Harper, I.; Ravussin, E.; Lynis Dohm, G.; Houmard, J.A. Substrate utilization during exercise in formerly morbidly obese women. J. Appl. Physiol. 2001, 90, 1007–1012. [Google Scholar] [CrossRef]
- Hickner, R.C.; Privette, J.; McIver, K.; Barakat, H. Fatty acid oxidation in African-American and Caucasian women during physical activity. J. Appl. Physiol. 2001, 90, 2319–2324. [Google Scholar] [CrossRef]
- Zurlo, F.; Lillioja, S.; Esposito-Del Puente, A.; Nyomba, B.L.; Raz, I.; Saad, M.F.; Swinbrun, B.A.; Knowler, W.C.; Bogardus, C.; Ravussin, E. Low ratio of fat to carbohydrate oxidation as predictor of weight gain: Study of 24-h RQ. Am. J. Physiol. 1990, 259, E650–E657. [Google Scholar] [CrossRef]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef]
- Little, J.P.; Gillen, J.B.; Percival, M.E.; Safdar, A.; Tarnopolsky, M.A.; Punthakee, Z.; Jung, M.E.; Gibala, M.J. Low-volume high intensity interval training reduces hyperglycemia and increases muscle mitochondrial capacity in patients with type 2 diabetes. J. Appl. Physiol. 2011, 111, 1554–1560. [Google Scholar] [CrossRef] [Green Version]
- Bogdanis, G.C.; Vangelakoudi, A.; Maridaki, M. Peak fat oxidation rate during walking in sedentary overweight men and women. J. Sports Sci. Med. 2008, 7, 525–531. [Google Scholar]
- Dumortier, M.; Brandou, F.; Perez-Martin, A.; Fedou, C.; Mercier, J.; Brun, J.F. Low intensity endurance exercise targeted for lipid oxidation improves body composition and insulin sensitivity in patients with the metabolic syndrome. Diabetes Metab. 2003, 29, 509–518. [Google Scholar] [CrossRef]
- Thompson, D.L.; Townsend, K.M.; Boughey, R.; Patterson, K.; Bassett, D.R., Jr. Substrate use during and following moderate– and low-intensity exercise: Implications for weight control. Eur. J. Appl. Physiol. Occup. Physiol. 1998, 78, 43–49. [Google Scholar] [CrossRef]
- Devries, M.C.; Samjoo, I.A.; Hamadeh, M.J.; McCready, C.; Raha, S.; Watt, M.J.; Steinberg, G.L.; Tarnopolsky, M.A. Endurance training modulates intramyocellular lipid compartmentalization and morphology in skeletal muscle of lean and obese women. J. Clin. Endocrinol. Metab. 2013, 98, 4852–4862. [Google Scholar] [CrossRef] [Green Version]
- Ardévol, A.; Adan, C.; Franco, L.; Garcia-Lorde, P.; Rubio, F.; Remesar, X. During intense exercise, obese women rely more than lean women on aerobic energy. Pflugers Arch. 1998, 43, 495–502. [Google Scholar] [CrossRef]
- Balci, Ş.S. Comparison of substrate oxidation during walking and running in normal-weight and overweight/obese men. Obes. Facts 2012, 5, 327–338. [Google Scholar] [CrossRef]
- Chatzinikolaou, A.; Fatouros, I.; Petridou, A.; Jamurtas, A.; Avloniti, A.; Douroudos, I.; Mastorakos, G.; Lazaropoulou, C.; Papassotiriou, I.; Tournis Mitrakou, A.; et al. Adipose tissue lipolysis is upregulated in lean and obese men during acute resistance exercise. Diabetes Care 2008, 31, 1397–1399. [Google Scholar] [CrossRef] [Green Version]
- Colberg, S.R.; Hagberg, J.M.; McCole, S.D.; Zmuda, J.M.; Thompson, P.D.; Kelley, D.E. Utilization of glycogen but not plasma glucose is reduced in individuals with NIDDM during mild-intensity exercise. J. Appl. Physiol. 1996, 81, 2027–2033. [Google Scholar] [CrossRef] [Green Version]
- Ezell, D.M.; Geiselman, P.J.; Anderson, A.M.; Dowdy, M.L.; Womble, L.G.; Greenway, F.L.; Zachwieja, J.J. Substrate oxidation and availability during acute exercise in non-obese, obese, and post-obese sedentary females. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Grams, L.; Kück, M.; Haufe, S.; Tegtbur, U.; Nelius, A.K.; Kerling, A. Peak fat oxidation during self-paced activities of daily life: Influence of sex and body composition. J. Sports Med. Phys. Fitness 2017, 57, 624–632. [Google Scholar]
- Kanaley, J.A.; Cryer, P.E.; Jensen, M.D. Fatty acid kinetics responses to exercise. Effects of obesity, body fat distribution, and energy-restricted diet. J. Clin. Invest. 1993, 92, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Kanaley, J.A.; Weatherup-Dentes, M.M.; Alvarado, C.R.; Whitehead, G. Substrate oxidation during acute exercise and with exercise training in lean and obese women. Eur. J. Appl. Physiol. 2001, 85, 68–73. [Google Scholar] [CrossRef]
- Melanson, E.L.; Gozansky, W.S.; Barry, D.W.; Maclean, P.S.; Grunwald, G.K.; Hill, J.O. When energy balance is maintained, exercise does not induce negative fat balance in lean sedentary, obese sedentary, or lean endurance-trained individuals. J. Appl. Physiol. 2009, 107, 1847–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanzi, S.; Codecasa, F.; Cornacchia, M.; Maestrini, S.; Salvadori, A.; Brunani, A.; Malatesta, D. Fat oxidation, hormonal and plasma metabolite kinetics during a submaximal incremental test in lean and obese adults. PLoS ONE 2014, 9, e88707. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.; Ara, I.; Rabøl, R.; Andersen, J.L.; Boushel, R.; Dela, F.; Helge, J.W. Are substrate use during exercise and mitochondrial respiratory capacity decreased in arm and leg muscle in type 2 diabetes? Diabetologia 2009, 52, 1400–1408. [Google Scholar] [CrossRef] [Green Version]
- Mittendorfer, B.; Fields, D.A.; Klein, S. Excess body fat in men decreases plasma fatty acid availability and oxidation during endurance exercise. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E354–E362. [Google Scholar] [CrossRef] [PubMed]
- Mohebbi, H.; Azizi, M. Maximal fat oxidation at the different exercise intensity in obese and normal weight men in the morning and evening. J. Hum. Sport Exerc. 2011, 6, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Martin, A.; Dumortier, M.; Raynaud, E.; Brun, J.F.; Fédou, C.; Bringer, J.; Mercier, J. Balance of substrate oxidation during submaximal exercise in lean and obese people. Diabetes Metab. 2001, 27, 466–474. [Google Scholar]
- Santiworakul, A.; Chuaychoo, B.; Kriengsinyos, W.; Saengsirisuwan, V.; Jalayondeja, W. Substrate utilization during and after high intensity exercise in healthy lean and obese men. J. Med. Assoc. Thai. 2014, 97, S50–S54. [Google Scholar]
- Slusher, A.L.; Whitehurst, M.; Zoeller, R.F.; Mock, J.T.; Maharaj, A.; Huang, C.J. Brain-derived neurotrophic factor and substrate utilization following acute aerobic exercise in obese individuals. J. Neuroendocrinol. 2015, 27, 370–376. [Google Scholar] [CrossRef]
- Steffan, H.G.; Elliott, W.; Miller, W.C.; Fernhall, B. Substrate utilization during submaximal exercise in obese and normal-weight women. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 233–239. [Google Scholar] [CrossRef]
- Keim, N.L.; Belko, A.Z.; Barbieri, T.F. Body fat percentage and gender: Associations with exercise energy expenditure, substrate utilization, and mechanical work efficiency. Int. J. Sport Nutr. 1996, 6, 356–369. [Google Scholar] [CrossRef] [Green Version]
- Eaves, A.D.; Colon, A.; Dubose, K.D.; Collier, D.; Houmard, J.A. Substrate utilization during submaximal exercise in children with a severely obese parent. Nutr. Metab. 2012, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Galgani, J.E.; Johannsen, N.M.; Bajpeyi, S.; Costford, S.R.; Zhang, Z.; Gupta, A.K.; Ravussin, E. Role of skeletal muscle mitochondrial density on exercise-stimulated lipid oxidation. Obesity 2012, 20, 1387–1393. [Google Scholar] [CrossRef]
- Boushel, R.; Gnaiger, E.; Schjerling, P.; Skovbro, M.; Kraunsoe, R.; Dela, F. Patients with type 2 diabetes have normal mitochondrial function in skeletal muscle. Diabetologia 2007, 50, 790–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogensen, M.; Sahlin, K.; Fernström, M.; Glintborg, D.; Vind, B.F.; Beck-Nielsen, H.; Højlund, K. Mitochondrial respiration is decreased in skeletal muscle of patients with type 2 diabetes. Diabetes 2007, 56, 1592–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phielix, E.; Schrauwen-Hinderling, V.B.; Mensink, M.; Lenaers, E.; Meex, R.; Hoeks, J.; Kooi, M.E.; Moonen-Kornips, E.; Sels, J.P.; Hesselink, M.K.; et al. Lower intrinsic ADP-stimulated mitochondrial respiration underlies in vivo mitochondrial dysfunction in muscle of male type 2 diabetic patients. Diabetes 2008, 57, 2943–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Goodpaster, B.H.; Thaete, F.L.; Simoneau, J.A.; Kelley, D.E. Subcutaneaous abdominal fat and thigh muscle composition predict insulin sensitivity independent of visceral fat. Diabetes 1997, 46, 1579–1585. [Google Scholar] [CrossRef]
- Blaak, E.E.; van Aggel-Leijssen, D.P.; Wagenmakers, A.J.; Saris, W.H.; van Baak, M.A. Impaired oxidation of plasma-derived fatty acids in type 2 diabetic subjects during moderate-intensity exercise. Diabetes 2000, 49, 2102–2107. [Google Scholar] [CrossRef] [Green Version]
- Blatchford, F.K.; Knowlton, R.G.; Schneider, D.A. Plasma FFA responses to prolonged walking in untrained men and women. Eur. J. Appl. Physiol. Occup. Physiol. 1985, 53, 343–347. [Google Scholar] [CrossRef]
- Froberg, K.; Pedersen, P.K. Sex differences in endurance capacity and metabolic response to prolonged, heavy exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1984, 52, 446–450. [Google Scholar] [CrossRef]
- Horton, T.J.; Pagliassotti, M.J.; Hobbs, K.; Hill, J.O. Fuel metabolism in men and women during and after long-duration exercise. J. Appl. Physiol. 1998, 85, 1823–1832. [Google Scholar] [CrossRef]
- Tarnopolsky, L.J.; MacDougall, J.D.; Atkinson, S.A.; Tarnopolsky, M.A.; Sutton, J.R. Gender differences in substrate for endurance exercise. J. Appl. Physiol. 1990, 68, 302–308. [Google Scholar] [CrossRef]
- Hellström, L.; Blaak, E.; Hagström-Toft, E. Gender differences in adrenergic regulation of lipid mobilization during exercise. Int. J. Sports Med. 1996, 17, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Mittendorfer, B.; Horowitz, J.F.; Klein, S. Effect of gender on lipid kinetics during endurance exercise of moderate intensity in untrained subjects. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E58–E65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitwood, L.F.; Brown, S.P.; Lundy, M.J.; Dupper, M.A. Metabolic propensity toward obesity in black vs. white females: Responses during rest, exercise and recovery. Int. J. Obes. Relat. Metab. Disord. 1996, 20, 455–462. [Google Scholar] [PubMed]
- Weyer, C.; Snitker, S.; Bogardus, C.; Ravussin, E. Energy metabolism in African Americans: Potential risk factors for obesity. Am. J. Clin. Nutr. 1999, 70, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, E.S.; Kovera, A.J.; Boozer, C.N.; Pi-Sunyer, F.X.; Albu, J.B. Metabolic inflexibility in substrate use is present in African-American but not Caucasian healthy, premenopausal, nondiabetic women. J. Clin. Endocrinol. Metab. 2006, 91, 4099–4106. [Google Scholar] [CrossRef]
- DeLany, J.P.; Dubé, J.J.; Standley, R.A.; Distefano, G.; Goodpaster, B.H.; Stefanovic-Racic, M.; Coen, P.M.; Toledo, F.G.S. Racial differences in peripheral insulin sensitivity and mitochondrial capacity in the absence of obesity. J. Clin. Endocrinol. Metab. 2014, 99, 4307–4314. [Google Scholar] [CrossRef]
- Ama, P.F.; Simoneau, J.A.; Boulay, M.R.; Serresse, O.; Thériault, G.; Bouchard, C. Skeletal muscle characteristics in sedentary black and Caucasian males. J. Appl. Physiol. 1986, 61, 1758–1761. [Google Scholar] [CrossRef]
- Costill, D.L.; Fink, W.J.; Getchell, L.H.; Ivy, J.L.; Witzmann, F.A. Lipid metabolism in skeletal muscle of endurance-trained males and females. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1979, 47, 787–791. [Google Scholar] [CrossRef]
- Martin, W.H., 3rd; Dalsky, G.P.; Hurley, B.F.; Matthews, D.E.; Bier, D.M.; Hagberg, J.M.; Rogers, M.A.; King, D.S.; Holloszy, J.O. Effect of endurance training on plasma free fatty acid turnover and oxidation during exercise. Am. J. Physiol. 1993, 265, E708–E714. [Google Scholar] [CrossRef]
- Phillips, S.M.; Green, H.J.; Tarnopolsky, M.A.; Heigenhauser, G.F.; Hill, R.E.; Grant, S.M. Effects of training duration on substrate turnover and oxidation during exercise. J. Appl. Physiol. 1996, 81, 2182–2191. [Google Scholar] [CrossRef]
- Bassett, D.R., Jr.; Howley, E.T. Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc. 2000, 32, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.D. New ideas on limitations to VO2max. Exerc. Sport Sci. Rev. 2000, 28, 10–14. [Google Scholar] [PubMed]
- Henriksson, J.; Reitman, J.S. Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 1977, 99, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Ara, I.; Larsen, S.; Stallknecht, B.; Guerra, B.; Morales-Alamo, D.; Andersen, J.L.; Ponce-González, J.G.; Guadalupe-Grau, A.; Galbo, H.; Calbet, J.A.L.; et al. Normal mitochondrial function and increased fat oxidation capacity in leg and arm muscles in obese humans. Int. J. Obes. 2011, 35, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Carey, D.G.; Jenkins, A.B.; Campbell, L.V.; Freund, J.; Chisholm, D.J. Abdominal fat and insulin resistance in normal and overweight women: Direct measurements reveal a strong relationship in subjects at both low and high risk of NIDDM. Diabetes 1996, 45, 633–638. [Google Scholar] [CrossRef]
- Goodyear, L.J.; Kahn, B.B. Exercise, glucose transport, and insulin sensitivity. Annu. Rev. Med. 1998, 49, 235–261. [Google Scholar] [CrossRef]
- Soeters, M.R.; Soeters, P.B. The evolutionary benefit of insulin resistance. Clin. Nutr. 2012, 31, 1002–1007. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [Green Version]
- Campbell, S.E.; Angus, D.J.; Febbraio, M.A. Glucose kinetics and exercise performance during phases of the menstrual cycle: Effect of glucose ingestion. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E817–E825. [Google Scholar] [CrossRef]
- Wenz, M.; Berend, J.Z.; Lynch, N.A.; Chappell, S.; Hackney, A.C. Substrate oxidation at rest and during exercise: Effects of menstrual cycle phase and diet composition. J. Physiol. Pharmacol. 1997, 48, 851–860. [Google Scholar]
- Zderic, T.W.; Coggan, A.R.; Ruby, B.C. Glucose kinetics and substrate oxidation during exercise in the follicular and luteal phases. J. Appl. Physiol. 2001, 90, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devries, M.C.; Hamadeh, M.J.; Phillips, S.M.; Tarnopolsky, M.A. Menstrual cycle phase and sex influence muscle glycogen utilization and glucose turnover during moderate-intensity endurance exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1120–R1128. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Abate, N. Role of subcutaneous adipose tissue in the pathogenesis of insulin resistance. J. Obes. 2013, 4, 489187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, N.; Schick, F.; Häring, H.U. Characteristics, and Consequences of Metabolically Unhealthy Normal Weight in Humans. Cell Metab. 2017, 26, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.D.; Haymond, M.W.; Rizza, R.A.; Cryer, P.E.; Miles, J.M. Influence of body fat distribution on free fatty acid metabolism in obesity. J. Clin. Invest. 1989, 83, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- Isacco, L.; Thivel, D.; Duclos, M.; Aucouturier, J.; Boisseau, N. Effects of adipose tissue distribution on maximum lipid oxidation rate during exercise in normal-weight women. Diabetes Metab. 2014, 40, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Isacco, L.; Duche, P.; Thivel, D.; Meddahi-Pelle, A.; Lemoine-Morel, S.; Duclos, M.; Boisseau, M. Fat mass localization alters fuel oxidation during exercise in normal weight women. Med. Sci. Sports Exerc. 2013, 45, 1887–1896. [Google Scholar] [CrossRef]
- Black, A.E.; Prentice, A.M.; Coward, W.A. Use of food quotients to predict respiratory quotients for the doubly-labelled water method of measuring energy expenditure. Hum. Nutr. Clin. Nutr. 1986, 40, 381–391. [Google Scholar]
- Miles-Chan, J.L.; Dulloo, A.G.; Schutz, Y. Fasting substrate oxidation at rest assessed by indirect calorimetry: Is prior dietary macronutrient level and composition a confounder? Int. J. Obes. 2015, 39, 1114–1117. [Google Scholar] [CrossRef] [Green Version]
- Abbott, W.G.; Howard, B.V.; Christin, L.; Freymond, D.; Lillioja, S.; Boyce, V.L.; Anderson, T.E.; Bogardus, C.; Ravussin, E. Short-term energy balance: Relationship with protein, carbohydrate, and fat balances. Am. J. Physiol. 1988, 255, E332–E337. [Google Scholar] [CrossRef]
- Acheson, K.J.; Schutz, Y.; Bessard, T.; Ravussin, E.; Jéquier, E.; Flatt, J.P. Nutritional influences on lipogenesis and thermogenesis after a carbohydrate meal. Am. J. Physiol. 1984, 246, E62–E70. [Google Scholar] [CrossRef] [PubMed]
- Flatt, J.P. Use and storage of carbohydrate and fat. Am. J. Clin. Nutr. 1995, 61, 952S–959S. [Google Scholar] [CrossRef] [PubMed]
- Schrauwen, P.; van Marken Lichtenbelt, W.D.; Saris, W.H.; Westerterp, K.R. Changes in fat oxidation in response to a high-fat diet. Am. J. Clin. Nutr. 1997, 66, 276–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.D.; Peters, J.C.; Reed, G.W.; Abumrad, N.N.; Sun, M.; Hill, J.O. Nutrient balance and energy expenditure during ad libitum feeding of high-fat and high-carbohydrate diets in humans. Am. J. Clin. Nutr. 1992, 55, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Ukropcova, B.; Sereda, O.; de Jonge, L.; Bogacka, I.; Nguyen, T.; Xie, H.; Bray, G.A.; Smith, S.R. Family history of diabetes links impaired substrate switching and reduced mitochondrial content in skeletal muscle. Diabetes 2007, 56, 720–727. [Google Scholar] [CrossRef] [Green Version]
- Montain, S.J.; Hopper, M.K.; Coggan, A.R.; Coyle, E.F. Exercise metabolism at different time intervals after a meal. J. Appl. Physiol. 1991, 70, 882–888. [Google Scholar] [CrossRef]
- Beltz, N.M.; Gibson, A.L.; Janot, J.M.; Kravitz, L.; Mermier, C.M.; Dalleck, L.C. Graded Exercise Testing Protocols for the Determination of VO2max: Historical Perspectives, Progress, and Future Considerations. J. Sports Med. 2016, 3968393. [Google Scholar] [CrossRef] [Green Version]
- Olsen, D.B.; Sacchetti, M.; Dela, F.; Ploug, T.; Saltin, B. Glucose clearance is higher in arm than leg muscle in type 2 diabetes. J. Physiol. 2005, 565, 555–562. [Google Scholar] [CrossRef]
- Reynolds, T.H., 4th; Supiano, M.A.; Dengel, D.R. Regional differences in glucose clearance: Effects of insulin and resistance training on arm and leg glucose clearance in older hypertensive individuals. J. Appl. Physiol. 2007, 102, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Gaster, M.; Staehr, P.; Beck-Nielsen, H.; Schrøder, H.D.; Handberg, A. GLUT4 is reduced in slow muscle fibers of type 2 diabetic patients: Is insulin resistance in type 2 diabetes a slow, type 1 fiber disease? Diabetes 2001, 50, 1324–1329. [Google Scholar] [CrossRef] [Green Version]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 1993, 265, E380–E391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillard, F.; Moro, C.; Harant, I.; Garrigue, E.; Lafontan, M.; Berlan, M.; Crampes, F.; de Glisezinski, I.; Rivière, D. Lipid oxidation according to intensity and exercise duration in overweight men and women. Obesity 2007, 15, 2256–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venables, M.C.; Achten, J.; Jeukendrup, A.E. Determinants of fat oxidation during exercise in healthy men and women: A cross-sectional study. J. Appl. Physiol. 2005, 98, 160–167. [Google Scholar] [CrossRef] [Green Version]
- DiMenna, F.J.; Jones, A.M. “Linear” versus “Nonlinear” O2 responses to exercise: Reshaping traditional beliefs. J. Exerc. Sci. Fit. 2009, 7, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Lansley, K.E.; DiMenna, F.J.; Bailey, S.J.; Jones, A.M. A ‘new’ method to normalise exercise intensity. Int. J. Sports Med. 2011, 32, 535–541. [Google Scholar] [CrossRef]
- Binder, R.K.; Wonisch, M.; Corrà, U.; Cohen-Solal, A.; Vanhees, L.; Saner, H.; Schmid, J.-P. Methodological approach to the first and second lactate threshold in incremental cardiopulmonary exercise testing. Eur. J. Cardiovasc. Prev. Rehabil. 2008, 15, 726–734. [Google Scholar] [CrossRef]
- Whipp, B.J.; Davis, J.A.; Wasserman, K. Ventilatory control of the ‘isocapnic buffering’ region in rapidly-incremental exercise. Respir. Physiol. 1989, 76, 357–367. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Study | Participants | Exercise for Assessment | Relevant Outcome Measure(s) | Lipid Use for Overweight/Obese | |
|---|---|---|---|---|---|
| Ardévol et al. (1998) [42] | Female | Leg cycling | ↔ | ||
| Obese: n = 8, age 30.0 years, BMI 32.3 ± 0.8 kg/m2, body fat 35.7% ± 1.1% | Control: n = 8, age 25.4 years, BMI 20.9 ± 0.5 kg/m2, body fat 23.4% ± 1.3% | Incremental O W + 30, W↑/3-min stage → Tlim | RQ | ||
| Balci (2012) [43] | Male | Treadmill | ↔ | ||
| Overweight/Obese: n = 9, age 21.4 ± 0.6 years, BMI 31.6 ± 1.1 kg/m2, body fat 24.2% ± 1.3% | Normal Weight: n = 10, age 21.9 ± 0.7 years, BMI 22.6 ± 0.4 kg/m2, body fat 13.8% ± 0.5% | CWR (1/session) 45 min at 1 km below PTS, 45 min at 1 km above PTS, Incremental Modified Bruce Protocol | Lipid oxidation rate (g∙min−1), Maximal lipid oxidation rate (g∙min−1, mg∙kgFFM−1∙min−1, RER, VO2∙kg−1BM, %VO2max, HR) | ||
| Chatzinikolaou et al. (2008) [44] | Male | Resistance | ↓ | ||
| Obese: n = 8, age 23.4 ± 0.8 years, BMI 31.2 ± 1.0 kg/m2, body fat 27.7% ± 1.2% | Lean: n = 9, age 23.8 ± 1.2 years, BMI 23.7 ± 0.5 kg/m2, body fat 11.1% ± 1.4% | Circuit 3 cycles × 10 exercises w/10–12 reps/set separated by 30-s rest | RER | ||
| Colberg et al. (1996) [45] | Male/Female | Leg cycling | ↔ | ||
| Obese: n = 4/3, age 50 ± 3 years, BMI 35.0 ± 1.4 kg/m2 | Lean: n = 3/4, age 48 ± 3 years, BMI 23.4 ± 1.0 kg/m2 | CWR 40 min at 40% VO2peak | RER Lipid oxidation rate (mg∙BM−1∙min−1). Lipid energy expenditure (% total) | ||
| Devries et al. (2013) [41] | Female | Leg cycling | ↔ | ||
| Obese: n = 11, age 40 ± 3 years, BMI pre 34 ± 2 kg/m2, BMI post 34 ± 2 kg/m2, body fat pre 49% ± 1%, body fat post 50% ± 2% | Lean: n = 12, age 41 ± 2 years, BMI pre 23 ± 1 kg/m2, BMI post 23 ± 1 kg/m2, body fat pre 32% ± 2%, body fat post 32% ± 2% | CWR 60 min at 50% VO2peak | RER Lipid oxidation rate (mg∙BM−1∙min−1, mg∙FFM−1∙min−1) | ||
| Ezell et al. (1999) [46] | Female | Leg cycling | ↔ | ||
| Obese: n = 5, age 26.2 ± 2.8 years, BMI 30.0 ± 2.7 kg/m2, body fat 44.4% ± 1.8% | Never obese: n = 5, age 25.6 ± 3.5 years, BMI 20.6 ± 0.9 kg/m2, body fat 25.0% ± 2.8% | CWR 60 min at 60–65% VO2peak | RER Total lipid oxidation (g, g∙BM−1, g∙FFM−1) | ||
| Goodpaster et al. (2002) [27] | Male | Leg Cycling | ↑ ↔ | ||
| Obese: n = 7, age 39.3 ± 3.2 years, BMI 33.7 ± 1.1 kg/m2, body fat 30.3% ± 1.3% | Lean: n = 7, age 34.3 ± 3.3 years, BMI 23.7 ± 0.7 kg/m2, body fat 20.4% ± 2.3% | CWR 60 min at 50% VO2max | RER Lipid oxidation rate (µmol∙FFM−1∙min−1), Lipid energy expenditure (MJ, % total) | ||
| Grams et al. (2017) [47] | Male/Female | Vacuuming/Floor Walking/Platform Stepping/Leg Cycling | ↓ ↔ | ||
| Hickner et al. (2001) [34] | Female | Leg cycling | ↓ ↔ | ||
| Obese African American: n = 11, age 30.9 ± 2.2 years, BMI 38.0 ± 1.8 kg/m2, body fat 41.2% ± 1.3%, Obese Caucasian: n = 9, age 34.1 ± 2.5 years, BMI 34.8 ± 0.9 kg/m2, body fat 39.3% ± 2.7% | Lean African American: n = 7; age 28.4 ± 2.8 years; BMI 23.1 ± 1.2 kg/m2; body fat 25.8% ± 2.8%. Lean Caucasian: n = 9; age 24.7 ± 1.8 years; BMI 23.5 ± 1.0 kg/m2; body fat 26.4% ± 2.0% | CWR (2 in succession). 10 min at 15 W;10 min at 65% VO2peak | RER Lipid oxidation rate (g∙min−1, g∙FFM−1∙h−1) | ||
| Horowitz et al. (2000) [28] | Female | Leg cycling (recumbent) | ↑ | ||
| Obese: n = 5; age premenopausal. BMI 37.7 ± 0.8 kg/m2, body fat 48.6% ± 1.9% | Lean: n = 5; age premenopausal. BMI 20.9 ± 0.4 kg/m2, body fat 25.4% ± 1.5% | CWR 90 min at 50% VO2peak | Lipid oxidation rate (µmol∙FFM−1∙min−1) | ||
| Kanaley et al. (1993) [48] | Female | Leg Cycling | ↔ | ||
| Obese lower body: n = 11; age 36 ± 2 years; BMI 31.5 ± 0.4 kg/m2; body fat 50% ± 3%; Obese upper body: n = 13; age 36 ± 2 years. BMI 33.4 ± 0.5 kg/m2; body fat 48% ± 2% | Non-obese: n = 8; age 36 ± 1 years; BMI 22.1 ± 0.6 kg/m2; body fat 30% ± 1% | CWR 150 min at 45% VO2peak | Lipid oxidation rate (µmol∙min−1)Total lipid oxidation (mmol) | ||
| Kanaley et al. (2001) [49] | Female | Treadmill | ↑ ↔ | ||
| Obese lower body: n = 11; age 32.0 ± 1.7 years; BMI 32.5 ± 0.5 kg/m2; body fat 38.2% ± 0.6%; Obese upper body: n = 12; age 32.5 ± 1.7 years; BMI 33.5 ± 1.0 kg/m2; body fat 38.9% ± 0.5% | Non-obese: n = 8; age 35.7 ± 1.4 years; BMI 21.7 ± 1.7 kg/m2; body fat 20.8% ± 1.4% | CWR 30 min at 70% VO2peak | RER Lipid oxidation rate (µmol∙FFM−1∙min−1) | ||
| Keim et al. (1996) [50] | Male/Female | Leg cycling | ↓ ↔ | ||
| Fatter: n = 8/8; age 34 ± 1/29 ± 2 years; body fat 22.1% ± 0.6/36.2% ± 1.8% | Leaner: n = 8/8; age 29 ± 1 years/32 ± 1 years; body fat 12.4% ± 0.8/20.6% ± 0.9% | Intermittent Incremental; 30, 60, 90, 120 W/5-min stage (female); 30, 60, 90, 120, 150 W/5-min stage (male) | RER Lipid oxidation rate (mg∙FFM−1∙min−1) | ||
| Lanzi et al. (2014) [51] | Male | Leg cycling | ↑ ↓ ↔ | ||
| Obese: n = 16; age 34.5 ± 2.1 years; BMI 39.0 ± 1.4 kg/m2; body fat 42.4% ± 1.4% | Lean: n = 16; age 33.1 ± 1.6 years; BMI 22.9 ± 0.3 kg/m2; body fat 19.8% ± 1.3% | Incremental 20% PPO + 7.5%↑/6-min stage → 65% or RER = 1.0 | RER Lipid oxidation rate (g∙min−1, mg∙FFM−1∙min−1); Maximal lipid oxidation rate (mg∙FFM−1∙min−1, %VO2peak, %HRmax, RER); Maximal lipid oxidation rate zone (%VO2peak) | ||
| Larsen et al. (2009) [52] | Male | Leg cycling/Arm cranking | ↑ ↔ | ||
| Obese: n = 8; age 37 ± 2 years; BMI 32 ± 1 kg/m2; body fat 32% ± 1% | Lean: n = 7; age 43 ± 3 years; BMI 25 ± 1 kg/m2; body fat 23% ± 1% | Incremental (leg); 95 W + 35 W↑/5-min stage → RER = 1.0 + 35 W↑/2-min stage → Tlim; Incremental (arm); 20 W + 15 W↑/6-min stage → 65 W + 5-min rest + 15 W↑/1-min stage → Tlim | Lipid oxidation rate (g∙min−1); Maximal lipid oxidation rate (g∙min−1, g∙BM−1∙min−1, g∙FFM−1∙min−1, %VO2max) | ||
| Melanson et al. (2009) [50] | Male/Female | Leg cycling | ↔ | ||
| Obese: n = 4/3; age 34 ± 5/44 ± 1 years; BMI 37.2 ± 3.3/31.7 ± 2.6 kg/m2; body fat 38.7% ± 3.1/40.8% ± 4.4% | Lean: n = 4/6; age 32 ± 12/30 ± 5 years; BMI 22.7 ± 2.9/22.4 ± 1.8 kg/m2; body fat 21.6% ± 6.6/29.9% ± 4.2% | CWR 60 min at 55% VO2peak | RER | ||
| Mittendorfer et al. (2003) [53] | Male | Leg cycling (recumbent) | ↔ | ||
| Overweight: n = 5; age 37 ± 4 years; BMI 27 ± 1 kg/m2; body fat 26% ± 1%; Obese: n = 5; age 38 ± 2 years; BMI 34 ± 1 kg/m2; body fat 30% ± 1% | Lean: n = 5; age 31 ± 3 years; BMI 21 ± 1 kg/m2; body fat 16% ± 2% | CWR 90 min at 50% VO2peak | Lipid oxidation rate (mg∙FFM−1∙min−1) | ||
| Mohebbi and Azizi (2011) [54] | Male | Treadmill | ↓ ↔ | ||
| Obese: n = 10; age 22.7 ± 2.0 years; BMI 32.5 ± 2.2 kg/m2; body fat 29.9% ± 5.3% | Normal Weight: n = 12; age 22.1 ± 1.5 years; BMI 22.3 ± 1.1 kg/m2; body fat 14.8% ± 3.9% | Incremental 3.5 km∙h−1 @ 1% + 1.0 km∙h−1↑/3-min stage × 4 stages + 2%↑/3-min stage → RER = 1.0 + speed ↑ → Tlim | Lipid oxidation rate (mg∙FFM−1∙min−1); Maximal lipid oxidation rate (mg∙FFM−1∙min−1, %VO2max); Minimal lipid oxidation rate (%VO2max) | ||
| Pérez-Martin et al. (2001) [55] | Male/Female | Leg Cycling | ↓ | ||
| Overweight: n = 15/17; age 44.0 ± 2.6/43.2 ± 2.6 years; BMI 32.1 ± 1.4/29.6 ± 0.9 kg/m2; body fat 32.7% ± 1.4/41.6% ± 0.9% | Control: n = 11/15; age 36.2 ± 3.7/41.1 ± 3.3 years; BMI 23.0 ± 0.6/23.0 ± 0.4 kg/m2; body fat 18.3% ± 1.5/25.9% ± 0.8% | Incremental 20% WRmax(est) + 10% WRmax(est) ↑/6-min stage ×4 stages | RER Lipid oxidation rate (mg∙min−1, mg∙FFM−1∙min−1); Power at maximal lipid oxidation rate (%WRmax(est), W); Crossover point (%WRmax(est), W, HR) | ||
| Santiworakul et al. (2014) [56] | Male | Mode not stated | ↔ | ||
| Obese: n = 10; age 25.6 ± 3.9 years; BMI 31.9 ± 2.5 kg/m2; body fat 35.9% ± 5.1% | Lean: n = 10; age 25.7 ± 4.0 years; BMI 21.6 ± 1.2 kg/m2; body fat 19.9% ± 8.1% | CWR x min at VT* (x = time to 300 kcal expenditure) | Lipid energy expenditure (% total, kcals) | ||
| Slusher et al. (2015) [57] | Male/Female | Treadmill | ↔ | ||
| Obese: n = 11; age 22.9 ± 1.6 years; BMI 35.7 ± 4.2 kg/m2 | Normal weight: n = 11; age 23.3 ± 2.2 years; BMI 22.0 ± 1.6 kg/m2 | CWR 30 min at 75% VO2max | Lipid oxidation rate (g∙min−1) | ||
| Steffan et al. (1999) [58] | Female | Treadmill | ↔ | ||
| Obese: n = 20; age 29.8 ± 1.3 years; BMI 31.0 ± 1.7 kg/m2; body fat 41.0% ± 1.5% | Normal weight: n = 15; age 25.1 ± 1.1 years; BMI 22.1 ± 0.7 kg/m2; body fat 26.1% ± 0.9% | Incremental Modified Bruce Protocol; CWR (1/session) 15 min at 50% VO2max15 min at 75% VO2max | RER | ||
| Thyfault et al. (2004) [26] | Female | Leg cycling | ↔ | ||
| Obese: n = 10; age 38.9 ± 1.9 years; BMI 40.8 ± 1.7 kg/m2 | Lean: n = 7; age 38.6 ± 2.3 years; BMI 22.6 ± 0.8 kg/m2 | CWR 60 min at 50% VO2max | RER Lipid oxidation rate (µmol∙BM−1∙min−1) | ||
| Wong et al. (2006) [23] | Male | Leg cycling | ↔ | ||
| Obese: n = 7; age 36.1 ± 3.4 years; BMI 31.9 ± 3.8 kg/m2; body fat 32.2% ± 5.7% | Lean: n = 6; age 34.5 ± 2.6 years; BMI 21.7 ± 1.7 kg/m2; body fat 15.4% ± 1.0% | CWR 30 min at VT** | RER | ||
| Determining Factor | Study | O < NW | O = NW | O > NW | Qualifications |
|---|---|---|---|---|---|
| Exercise duration (min) | Kanaley et al. 2001 [49] | 15 | 30 | For lipid oxidation rate (LBO and UBO) or RER (UBO) | |
| Exercise intensity (%VO2peak/max) | Hickner et al. 2001 [34] | 65 | ~40 | For lipid oxidation rate for Caucasians; O < NW (RER) for Caucasians at both intensities and O = NW (lipid oxidation rate, RER) for AA at both intensities | |
| Keim et al. 1996 [59] | 40–60 | 30 | For men; O = NW at all intensities for women | ||
| Lanzi et al. 2014 [51] | - | 60–85 | 20–55 | For RER | |
| 85 | 50–80 | 20–45 | For lipid oxidation rate (g∙min−1) | ||
| 65–85 | 35–60 | 20–30 | For lipid oxidation rate (mg∙kgFFM−1∙min−1) | ||
| Mohebbi and Azizi 2011 [54] | 60–80 | 20–50 | For AM and PM | ||
| Exercise mode | Grams et al. 2017 [47] | Platform stepping | Vacuuming Floor Walking Leg cycling | For women; O < NW for men for stepping and cycling (lipid oxidation rate) or all four activities (% lipid energy) | |
| Larsen et al. 2009 [52] | Leg cycling | Arm cranking | For fatmax % VO2max; O = NW for Fatmax g∙min−1, mg∙kgBM−1∙min−1, mg∙kgFFM−1∙min−1 | ||
| Exercise work rate (W) | Keim et al. 1996 [59] | 120 | 30–90, 150 | For men | |
| Lanzi et al. 2014 [51] | 150 | 75–150 | 30–60 | For RER | |
| 90–135 | 30–75 | For lipid oxidation rate | |||
| Outcome measure | Goodpaster et al. 2002 [27] | Lipid energy (MJ); Lipid oxidation rate (µmol∙FFM−1∙min−1) | Lipid energy (%) RER | p = 0.08 | |
| Grams et al. 2017 [47] | Lipid energy (%); Lipid oxidation rate (kcal∙min−1); Lipid energy (%) | Lipid oxidation rate (kcal∙min−1); Peak lipid oxidation rate (kcal∙min−1, mg∙kgFFM−1∙min−1, %VO2max); Peak lipid oxidation rate (kcal∙min−1, mg∙kgFFM−1∙min−1, %VO2max) | For walking (men); For stepping (both sexes) and cycling (men); For stepping (both sexes) and vacuuming, walking, cycling (men) | ||
| Hickner et al. 2001 [34] | RER | Lipid oxidation rate (g∙min−1, g∙FFM−1∙h−1) | For obese Caucasian and AA v. lean Caucasian | ||
| Kanaley et al. 2001 [49] | RER | Lipid oxidation rate (µmol∙FFM−1∙min−1) | For LBO at 30 min | ||
| Lanzi et al. 2014 [51] | Lipid oxidation rate (g∙min−1); Lipid oxidation rate (mg∙FFM−1∙min−1); Maximal lipid oxidation rate (%VO2peak, %HRmax, RER) and zone (%VO2peak) | RER RER Maximal lipid oxidation rate (mg∙FFM−1∙min−1) | At 150 W and 85% VO2peakAt 65–85% VO2peak | ||
| Larsen et al. 2009 [52] | Lipid oxidation rate (g∙min−1); Maximal lipid oxidation rate (g∙min−1, g∙BM−1∙min−1, g∙FFM−1∙min−1) | Maximal lipid oxidation rate (%VO2max) | For arm cranking | ||
| Mohebbi and Azizi 2011 [54] | Lipid oxidation rate (mg∙FFM−1∙min−1); Maximal lipid oxidation rate (%VO2max); Maximal lipid oxidation rate (%VO2max) | Maximal lipid oxidation rate (mg∙FFM−1∙min−1, %VO2max; Lipid oxidation rate (mg∙FFM−1∙min−1); Maximal lipid oxidation rate (mg∙kgFFM−1∙min−1); Minimal lipid oxidation rate (%VO2max) | For lipid oxidation rate at 60–80% VO2max For lipid oxidation rate at 20%–50% VO2max | ||
| Participant fat deposition | Kanaley et al. 2001 [49] | LBO | UBO | For RER at 30 min; O < NW for LB and UB Obese for lipid oxidation rate at 30 min | |
| Participant race | Hickner et al. 2001 [34] | Caucasian O | AA O | For cycling at 15 W (RER, lipid oxidation rate) and 65%VO2peak (lipid oxidation rate) | |
| Participant sex | Grams et al. 2017 [47] | Male O | Female O | For cycling (% lipid energy, lipid oxidation rate) and vacuuming, walking (% lipid energy) | |
| Keim et al. 1996 [59] | Male O | Female O | For cycling at 40–60% VO2max; O = NW for men and women at 30% VO2max |
| Study | Aim Clearly Stated and Defined | Eligibility and Inclusion Criteria Explained | Study Population Clearly Specified and Defined | Sample Size Justification Provided | Participants Recruited from Same or Similar Population | Independent Variable Clearly Defined, Valid and Reliable | Dependent Variable Clearly Defined, Valid, and Reliable | Avg Score |
|---|---|---|---|---|---|---|---|---|
| Ardévol et al. 1998 [42] | 2.0 | 2.0 | 2.0 | 0.0 | 0.5 | 1.5 | 2.0 | 1.43 |
| Balci 2012 [43] | 2.0 | 2.0 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.50 |
| Chatzinikolaou et al. 2008 [44] | 2.0 | 1.0 | 2.0 | 0.0 | 0.5 | 1.5 | 2.0 | 1.29 |
| Colberg et al. 1996 [45] | 2.0 | 2.0 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.50 |
| Devries et al. 2013 [41] | 2.0 | 2.0 | 2.0 | 0.0 | 1.0 | 2.0 | 2.0 | 1.57 |
| Ezell et al. 1999 [46] | 2.0 | 2.0 | 2.0 | 0.0 | 0.5 | 1.5 | 2.0 | 1.43 |
| Goodpaster et al. 2002 [27] | 2.0 | 2.0 | 2.0 | 1.0 | 1.5 | 2.0 | 2.0 | 1.79 |
| Grams et al. 2017 [47] | 2.0 | 2.0 | 2.0 | 0.5 | 1.0 | 2.0 | 2.0 | 1.64 |
| Hickner et al. 2001 [34] | 2.0 | 1.5 | 2.0 | 0.0 | 0.5 | 1.5 | 2.0 | 1.36 |
| Horowitz et al. 2000 [28] | 2.0 | 1.5 | 2.0 | 2.0 | 1.0 | 2.0 | 2.0 | 1.79 |
| Kanaley et al. 1993 [48] | 2.0 | 1.5 | 2.0 | 0.0 | 0.0 | 2.0 | 2.0 | 1.36 |
| Kanaley et al. 2001 [49] | 2.0 | 2.0 | 2.0 | 0.0 | 2.0 | 2.0 | 2.0 | 1.71 |
| Keim et al. 1996 [59] | 2.0 | 2.0 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.50 |
| Lanzi et al. 2014 [51] | 2.0 | 2.0 | 2.0 | 0.0 | 1.5 | 2.0 | 2.0 | 1.64 |
| Larsen et al. 2009 [52] | 2.0 | 1.5 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.43 |
| Melanson et al. 2009 [50] | 2.0 | 2.0 | 2.0 | 0.0 | 2.0 | 2.0 | 2.0 | 1.71 |
| Mittendorfer et al. 2003 [53] | 2.0 | 1.0 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.36 |
| Mohebbi and Azizi 2011 [54] | 2.0 | 2.0 | 2.0 | 0.0 | 1.0 | 2.0 | 2.0 | 1.57 |
| Pérez-Martin et al. 2001 [55] | 2.0 | 1.0 | 1.5 | 0.0 | 1.0 | 2.0 | 2.0 | 1.36 |
| Santiworakul et al. 2014 [56] | 2.0 | 1.5 | 1.5 | 2.0 | 0.0 | 1.5 | 2.0 | 1.50 |
| Slusher et al. 2015 [57] | 2.0 | 2.0 | 2.0 | 0.0 | 0.0 | 2.0 | 2.0 | 1.43 |
| Steffan et al. 1999 [58] | 2.0 | 1.0 | 2.0 | 0.0 | 1.0 | 2.0 | 2.0 | 1.43 |
| Thyfault et al. 2004 [26] | 2.0 | 2.0 | 2.0 | 0.0 | 0.0 | 2.0 | 2.0 | 1.43 |
| Wong et al. 2006 [23] | 2.0 | 1.5 | 2.0 | 0.0 | 0.5 | 2.0 | 2.0 | 1.43 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arad, A.D.; Basile, A.J.; Albu, J.; DiMenna, F.J. No Influence of Overweight/Obesity on Exercise Lipid Oxidation: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 1614. https://doi.org/10.3390/ijms21051614
Arad AD, Basile AJ, Albu J, DiMenna FJ. No Influence of Overweight/Obesity on Exercise Lipid Oxidation: A Systematic Review. International Journal of Molecular Sciences. 2020; 21(5):1614. https://doi.org/10.3390/ijms21051614
Chicago/Turabian StyleArad, Avigdor D., Anthony J. Basile, Jeanine Albu, and Fred J. DiMenna. 2020. "No Influence of Overweight/Obesity on Exercise Lipid Oxidation: A Systematic Review" International Journal of Molecular Sciences 21, no. 5: 1614. https://doi.org/10.3390/ijms21051614
APA StyleArad, A. D., Basile, A. J., Albu, J., & DiMenna, F. J. (2020). No Influence of Overweight/Obesity on Exercise Lipid Oxidation: A Systematic Review. International Journal of Molecular Sciences, 21(5), 1614. https://doi.org/10.3390/ijms21051614