Transcriptional and Metabolomic Analysis of L-Arginine/Nitric Oxide Pathway in Inflammatory Bowel Disease and Its Association with Local Inflammatory and Angiogenic Response: Preliminary Findings

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Systemic Concentrations of L-arginine/NO Pathway Metabolites
2.1.1. L-arginine/NO Pathway Metabolites in IBD and in Irritable Bowel Syndrome (IBS)
2.1.2. L-Arginine/NO Pathway Metabolites and IBD Phenotype and Clinical Activity
2.1.3. L-arginine/NO Pathway Metabolites and the Endoscopic Activity of UC
2.1.4. Interrelationship between L-arginine/NO Pathway Metabolites
2.2. L-arginine/NO Pathway-Associated Enzymes in IBD—Transcriptional Analysis
2.2.1. L-arginine/NO Pathway-Associated Enzymes in Inflamed and Quiescent Small Bowel
2.2.2. L-arginine/NO Pathway-Associated Enzymes in Inflamed and Quiescent Large Bowel
2.2.3. Interrelationship between Expression Levels of L-arginine/NO Pathway-Associated Enzymes in the Small and Large Bowel
2.2.4. Expression of L-arginine/NO Pathway-Associated Enzymes in the Bowel and Local Inflammatory and Angiogenic Response, Hypoxia, Proliferation, and Apoptosis
2.3. Expression of L-arginine/NO Pathway-Associated Enzymes in the Small and Large Bowel and Systemic Concentrations of Pathway Metabolites
3. Discussion
4. Materials and Methods
4.1. Patients
4.1.1. Metabolomic Analysis
4.1.2. Transcriptional Analysis
4.2. Ethical Considerations
4.3. Analytical Methods
4.3.1. Sample Collection and Preparation
4.3.2. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
4.3.3. Reverse-Transcription Quantitative (Real-Time) Polymerase Chain Reaction (RT-qPCR)
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IBD | Inflammatory bowel disease |
| IBS | Irritable bowel syndrome |
| CD | Crohn’s disease |
| UC | Ulcerative colitis |
| CDAI | Crohn’s Disease Activity Index |
| RI | Rachmilewitz index |
| ADMA | Asymmetric dimethylarginine |
| SDMA | Symmetric dimethylarginine |
| DMA | Dimethylamine |
| PRMT | Arginine N-methyltransferase |
| DDAH | Dimethylarginine dimethylaminohydrolase |
| NOS | Nitric oxide synthase |
| ODC | Ornithine decarboxylase |
| ARG | Arginase |
References
- Windsor, J.W.; Kaplan, G.G. Evolving epidemiology of IBD. Curr. Gastroenterol. Rep. 2019, 21, 40. [Google Scholar] [CrossRef] [PubMed]
- Gecse, K.B.; Vermeire, S. Differential diagnosis of inflammatory bowel disease: Imitations and complications. Lancet. Gastroenterol. Hepatol. 2018, 3, 644–653. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.; Chan, F.K.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.-F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef]
- Fichtner-Feigl, S.; Kesselring, R.; Strober, W. Chronic inflammation and the development of malignancy in the GI tract. Trends Immunol. 2015, 36, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Predonzani, A.; Calì, B.; Agnellini, A.H.; Molon, B. Spotlights on immunological effects of reactive nitrogen species: When inflammation says nitric oxide. World J. Exp. Med. 2015, 5, 64–76. [Google Scholar] [CrossRef]
- Middleton, S.J.; Shorthouse, M.; Hunter, J.O. Increased nitric oxide synthesis in ulcerative colitis. Lancet 1993, 341, 465–466. [Google Scholar] [CrossRef]
- Rachmilewitz, D.; Stamler, J.S.; Bachwich, D.; Karmeli, F.; Ackerman, Z.; Podolsky, D.K. Enhanced colonic nitric oxide generation and nitric oxide synthase activity in ulcerative colitis and Crohn’s disease. Gut 1995, 36, 718–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model. Mech. 2018, 11, dmm033332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, S.; Wang, X.; Liu, P.; Ke, C.; Xu, S. Arginine metabolism and deprivation in cancer therapy. Biomed. Pharmacother. 2019, 118, 109210. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Arginine metabolism: Boundaries of our knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurpinar, E.; Grizzle, W.E.; Piazza, G.A. COX-Independent mechanisms of cancer chemoprevention by anti-inflammatory drugs. Front. Oncol. 2013, 3, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyskens, F.L., Jr.; McLaren, C.E.; Pelot, D.; Fujikawa-Brooks, S.; Carpenter, P.M.; Hawk, E.; Kelloff, G.; Lawson, M.J.; Kidao, J.; McCracken, J.; et al. Difluoromethylornithine plus sulindac for the prevention of sporadic colorectal adenomas: A randomized placebo-controlled, double-blind trial. Cancer Prev. Res. (Phila) 2008, 1, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Gerner, E.W.; Bruckheimer, E.; Cohen, A. Cancer pharmacoprevention: Targeting polyamine metabolism to manage risk factors for colon cancer. J. Biol. Chem. 2018, 293, 18770–18778. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.H.; Guo, L.J.; Huang, Z.Y.; Rao, J.N.; Tang, C.W. Roles of cellular polyamines in mucosal healing in the gastrointestinal tract. J. Physiol. Pharmacol. 2013, 64, 681–693. [Google Scholar]
- Tofalo, R.; Cocchi, S.; Suzzi, G. Polyamines and gut microbiota. Front. Nutr. 2019, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Szefel, J.; Danielak, A.; Kruszewski, W.J. Metabolic pathways of L-arginine and therapeutic consequences in tumors. Adv. Med. Sci. 2019, 64, 104–110. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Chen, S.; Yin, Y. L-arginine and inflammatory bowel diseases (IBD). In L-Arginine in Clinical Nutrition; Patel, V., Preedy, V., Rajendram, R., Eds.; Nutrition and Health. Humana Press: Cham, Switzerland, 2017; pp. 331–342. [Google Scholar] [CrossRef]
- Coburn, L.A.; Singh, K.; Asim, M.; Barry, D.P.; Allaman, M.M.; Al-Greene, N.T.; Hardbower, D.M.; Polosukhina, D.; Williams, C.S.; Delgado, A.G.; et al. Loss of solute carrier family 7 member 2 exacerbates inflammation-associated colon tumorigenesis. Oncogene 2018, 38, 1067–1079. [Google Scholar] [CrossRef]
- Owczarek, D.; Cibor, D.; Mach, T. Asymmetric dimethylarginine (ADMA), symmetric dimethylarginine (SDMA), arginine, and 8-iso-prostaglandin F2alpha (8-iso-PGF2alpha) level in patients with inflammatory bowel diseases. Inflamm. Bowel. Dis. 2010, 16, 52–57. [Google Scholar] [CrossRef]
- Fragkos, K.C.; Forbes, A. Citrulline as a marker of intestinal function and absorption in clinical settings: A systematic review and meta-analysis. United Eur. Gastroenterol. J. 2018, 6, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Curis, E.; Crenn, P.; Cynober, L. Citrulline and the gut. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Coburn, L.A.; Horst, S.N.; Allaman, M.M.; Brown, C.T.; Williams, C.S.; Hodges, M.E.; Druce, J.P.; Beaulieu, D.B.; Schwartz, D.A.; Wilson, K.T. L-arginine availability and metabolism is altered in ulcerative colitis. Inflamm. Bowel. Dis. 2016, 22, 1847–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colón, A.L.; Menchén, L.; Lizasoain, I.; Leza, J.C.; Menchén, P.; González-Lara, V.; Moro, M.A.; Lorenzo, P. Inducible nitric oxide synthase activity is expressed not only in inflamed but also in normal colonic mucosa in patients with ulcerative colitis: A potential prognostic marker. Am. J. Gastroenterol. 2000, 95, 1371–1373. [Google Scholar] [CrossRef]
- Blanc, R.S.; Richard, S. Arginine methylation: The coming of age. Mol. Cell 2017, 65, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Wang, B.; Miyagi, M.; She, Y.; Gopalan, B.; Huang, D.B.; Ghosh, G.; Stark, G.R.; Lu, T. PRMT5 dimethylates R30 of the p65 subunit to activate NF-κB. Proc. Natl. Acad. Sci. USA 2013, 110, 13516–13521. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Huang, L.; Ge, W.; Yang, M.; Ma, Y.; Xie, G.; Wang, W.; Bian, B.; Li, L.; Nie, H.; et al. Protein arginine methyltransferase 5 inhibition upregulates Foxp3+ regulatory T cells frequency and function during the ulcerative colitis. Front. Immunol. 2017, 8, 596. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Harris, D.P.; Adams, G.N.; Lause, G.E.; McHugh, A.; Tillmaand, E.G.; Money, A.; Willard, B.; Fox, P.L.; Dicorleto, P.E. HOXA9 methylation by PRMT5 is essential for endothelial cell expression of leukocyte adhesion molecules. Mol. Cell Biol. 2012, 32, 1202–1213. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.T.; Guo, J.; Prajapati, S.; Park, K.J.; Surabhi, R.M.; Miller, B.; Gehrig, P.; Gaynor, R.B. Methylation of SPT5 regulates its interaction with RNA polymerase II and transcriptional elongation properties. Mol. Cell 2003, 11, 1055–1066. [Google Scholar] [CrossRef]
- Reintjes, A.; Fuchs, J.E.; Kremser, L.; Lindner, H.H.; Liedl, K.R.; Huber, L.A.; Valovka, T. Asymmetric arginine dimethylation of RelA provides a repressive mark to modulate TNFalpha/NF-kappaB response. Proc. Natl. Acad. Sci. USA 2016, 113, 4326–4331. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Li, J.; Li, P.; Ye, Q.; Xu, H.; Wu, X.; Xu, Y. Protein arginine methyltransferase 1 (PRMT1) represses MHC II transcription in macrophages by methylating CIITA. Sci. Rep. 2017, 7, 40531. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.; Bauer, U.-M. Arginine methylation in interferon signaling: New light on an old story. Cell Cycle 2009, 8, 1461–1465. [Google Scholar] [CrossRef] [Green Version]
- Mowen, K.A.; Tang, J.; Zhu, W.; Schurter, B.T.; Shuai, K.; Herschman, H.R.; David, M. Arginine methylation of STAT1 modulates IFNalpha/beta-induced transcription. Cell 2001, 104, 731–741. [Google Scholar] [CrossRef]
- Tan, Z.; Chen, K.; Wu, W.; Zhou, Y.; Zhu, J.; Wu, G.; Cao, L.; Zhang, X.; Guan, H.; Yang, Y.; et al. Overexpression of HOXC10 promotes angiogenesis in human glioma via interaction with PRMT5 and upregulation of VEGFA expression. Theranostics 2018, 8, 5143–5158. [Google Scholar] [CrossRef]
- Bruns, A.F.; Grothe, C.; Claus, P. Fibroblast growth factor 2 (FGF-2) is a novel substrate for arginine methylation by PRMT5. Biol. Chem. 2009, 390, 59–65. [Google Scholar] [CrossRef]
- Lovisa, S.; Genovese, G.; Danese, S. Role of epithelial-to-mesenchymal transition in inflammatory bowel disease. J. Crohns Colitis 2019, 13, 659–668. [Google Scholar] [CrossRef]
- Zhu, Z.D.; Ye, J.M.; Fu, X.M.; Wang, X.C.; Ye, J.Y.; Wu, X.R.; Hua, P.; Liao, Y.Q.; Xuan, W.; Duan, J.L.; et al. DDAH2 alleviates myocardial fibrosis in diabetic cardiomyopathy through activation of the DDAH/ADMA/NOS/NO pathway in rats. Int. J. Mol. Med. 2019, 43, 749–760. [Google Scholar] [CrossRef]
- Pirola, L.; Ciesielski, O.; Balcerczyk, A. The methylation status of the epigenome: Its emerging role in the regulation of tumor angiogenesis and tumor growth, and potential for drug targeting. Cancers (Basel) 2018, 10, 268. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, P.; Vaites, L.P.; Kim, J.K.; Mellert, H.; Gurung, B.; Nakagawa, H.; Herlyn, M.; Hua, X.; Rustgi, A.K.; McMahon, S.B.; et al. Nuclear cyclin D1/CDK4 kinase regulates CUL4 expression and triggers neoplastic growth via activation of the PRMT5 methyltransferase. Cancer Cell 2010, 18, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.Y.; Juan, C.C.; Hisa, J.Y.; Su, L.J.; Lee, Y.C.; Chou, H.Y.; Chen, J.M.; Wu, Y.C.; Chiu, S.C.; Hsu, C.P.; et al. Protein arginine methyltransferase 5 is a potential oncoprotein that upregulates G1 cyclins/cyclin-dependent kinases and the phosphoinositide 3-kinase/AKT signaling cascade. Cancer Sci. 2012, 103, 1640–1650. [Google Scholar] [CrossRef]
- Karkhanis, V.; Hu, Y.J.; Baiocchi, R.A.; Imbalzano, A.N.; Sif, S. Versatility of PRMT5-induced methylation in growth control and development. Trends Biochem. Sci. 2011, 36, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielli, P.; Pagliarini, V.; Pieraccioli, M.; Caggiano, C.; Sette, C. Splicing dysregulation as oncogenic driver and passenger factor in brain tumors. Cells 2019, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.; Cao, R.X.; Liu, J.H.; Liu, Y.B.; Wang, J.; Liu, L.P.; Chen, Y.J.; Yang, J.; Zhang, Q.H.; Wu, Y.; et al. Nuclear loss of protein arginine N-methyltransferase 2 in breast carcinoma is associated with tumor grade and overexpression of cyclin D1 protein. Oncogene 2014, 33, 5546–5558. [Google Scholar] [CrossRef] [Green Version]
- Scoumanne, A.; Zhang, J.; Chen, X. PRMT5 is required for cell-cycle progression and p53 tumor suppressor function. Nucleic Acids Res. 2009, 37, 4965–4976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulin, J.-A.; Gubareva, E.A.; Jarzebska, N.; Rodionov, R.N.; Mangoni, A.A.; Tommasi, S. Inhibition of dimethylarginine dimethylaminohydrolase (DDAH) enzymes as an emerging therapeutic strategy to target angiogenesis and vasculogenic mimicry in cancer. Front. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Toxic dimethylarginines: Asymmetric dimethylarginine (ADMA) and symmetric dimethylarginine (SDMA). Toxins (Basel) 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buijs, N.; Oosterink, J.E.; Jessup, M.; Schierbeek, H.; Stolz, D.B.; Houdijk, A.P.; Geller, D.A.; van Leeuwen, P.A. A new key player in VEGF-dependent angiogenesis in human hepatocellular carcinoma: Dimethylarginine dimethylaminohydrolase 1. Angiogenesis 2017, 20, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Kostourou, V.; Robinson, S.P.; Cartwright, J.E.; St. J. Whitley, G. Dimethylarginine dimethylaminohydrolase I promotes tumour growth and angiogenesis. Br. J. Cancer 2002, 87, 673–680. [Google Scholar] [CrossRef]
- Shiozawa, T.; Iyama, S.; Toshima, S.; Sakata, A.; Usui, S.; Minami, Y.; Sato, Y.; Hizawa, N.; Noguchi, M. Dimethylarginine dimethylaminohydrolase 2 promotes tumor angiogenesis in lung adenocarcinoma. Virchows Arch. 2016, 468, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Lawrance, I.C.; Rogler, G.; Bamias, G.; Breynaert, C.; Florholmen, J.; Pellino, G.; Reif, S.; Speca, S.; Latella, G. Cellular and molecular mediators of intestinal fibrosis. J. Crohns Colitis 2017, 11, 491–1503. [Google Scholar] [CrossRef] [Green Version]
- Dolivo, D.M.; Larson, S.A.; Dominko, T. Fibroblast growth factor 2 as an antifibrotic: Antagonism of myofibroblast differentiation and suppression of pro-fibrotic gene expression. Cytokine Growth Factor Rev. 2017, 38, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Xu, J.; Li, Y.; Huang, Q.; Huang, J.; Wang, J.; Zhong, W.; Lin, X.; Chen, W.; Lin, X. DDAH1 mediates gastric cancer cell invasion and metastasis via Wnt/β-catenin signaling pathway. Mol. Oncol. 2017, 11, 1208–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.R.K.; Dasari, C.; Duscharla, D.; Supriya, B.; Ram, N.S.; Surekha, M.V.; Kumar, J.M.; Ummanni, R. Dimethylarginine dimethylaminohydrolase-1 (DDAH1) is frequently upregulated in prostate cancer, and its overexpression conveys tumor growth and angiogenesis by metabolizing asymmetric dimethylarginine (ADMA). Angiogenesis 2018, 21, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Avdagić, N.; Zaćiragić, A.; Babić, N.; Hukić, M.; Seremet, M.; Lepara, O.; Nakaš-Ićindić, E. Nitric oxide as a potential biomarker in inflammatory bowel disease. Bosn. J. Basic Med. Sci. 2013, 13, 5–9. [Google Scholar]
- Baranipour, S.; Amini Kadijani, A.; Qujeq, D.; Shahrokh, S.; Haghazali, M.; Mirzaei, A.; Asadzadeh-Aghdaei, H. Inducible nitric oxide synthase as a potential blood-based biomarker in inflammatory bowel diseases. Gastroenterol. Hepatol. Bed Bench 2018, 11, S124–S128. [Google Scholar]
- Singh, K.; Gobert, A.P.; Coburn, L.A.; Barry, D.P.; Allaman, M.; Asim, M.; Luis, P.B.; Schneider, C.; Milne, G.L.; Boone, H.H.; et al. Dietary arginine regulates severity of experimental colitis and affects the colonic microbiome. Front. Cell Infect. Microbiol. 2019, 9, 66. [Google Scholar] [CrossRef]
- Kolios, G.; Rooney, N.; Murphy, C.T.; Robertson, D.A.; Westwick, J. Expression of inducible nitric oxide synthase activity in human colon epithelial cells: Modulation by T lymphocyte derived cytokines. Gut 1998, 43, 56–63. [Google Scholar] [CrossRef]
- Latour, Y.L.; Gobert, A.P.; Wilson, K.T. The role of polyamines in the regulation of macrophage polarization and function. Amino Acids 2019. [Google Scholar] [CrossRef]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A multifaceted enzyme important in health and disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef] [Green Version]
- Setty, B.A.; Jin, Y.; Houghton, P.J.; Yeager, N.D.; Gross, T.G.; Nelin, L.D. Hypoxic proliferation of osteosarcoma cells depends on arginase II. Cell Physiol. Biochem. 2016, 39, 802–813. [Google Scholar] [CrossRef]
- Lasch, M.; Caballero-Martinez, A.; Troidl, K.; Schloegl, I.; Lautz, T.; Deindl, E. Arginase inhibition attenuates arteriogenesis and interferes with M2 macrophage accumulation. Lab. Invest. 2016, 96, 830–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molderings, G.J.; Haenisch, B. Agmatine (decarboxylated l-arginine): Physiological role and therapeutic potential. Pharmacol. Ther. 2012, 133, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Seiler, N.; Raul, F. Polyamines and apoptosis. J. Cell Mol. Med. 2005, 9, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Regulation of ornithine decarboxylase. J. Biol. Chem. 2006, 281, 14529–14532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Coburn, L.A.; Asim, M.; Barry, D.P.; Allaman, M.M.; Shi, C.; Washington, M.K.; Luis, P.B.; Schneider, C.; Delgado, A.G.; et al. Ornithine decarboxylase in macrophages exacerbates colitis and promotes colitis-associated colon carcinogenesis by impairing M1 immune responses. Cancer Res. 2018, 78, 4303–4315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, W.H. Autoimmune diseases and polyamines. Clinic Rev. Allerg. Immunol. 2012, 42, 58–70. [Google Scholar] [CrossRef]
- Fleszar, M.G.; Wiśniewski, J.; Krzystek-Korpacka, M.; Misiak, B.; Frydecka, D.; Piechowicz, J.; Lorenc-Kukuła, K.; Gamian, A. Quantitative analysis of L-arginine, dimethylated arginine derivatives, L-citrulline, and dimethylamine in human serum using liquid chromatography–mass spectrometric method. Chromatographia 2018, 81, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Chachaj, A.; Wiśniewski, J.; Rybka, J.; Butrym, A.; Biedroń, M.; Krzystek-Korpacka, M.; Fleszar, M.G.; Karczewski, M.; Wróbel, T.; Mazur, G.; et al. Asymmetric and symmetric dimethylarginines and mortality in patients with hematological malignancies-A prospective study. PLoS ONE 2018, 13, e0197148. [Google Scholar] [CrossRef] [Green Version]
- Fleszar, M.G.; Wiśniewski, J.; Zboch, M.; Diakowska, D.; Gamian, A.; Krzystek-Korpacka, M. Targeted metabolomic analysis of nitric oxide/L-arginine pathway metabolites in dementia: Association with pathology, severity, and structural brain changes. Sci. Rep. 2019, 9, 13764. [Google Scholar] [CrossRef]
- Krzystek-Korpacka, M.; Wiśniewski, J.; Fleszar, M.G.; Bednarz-Misa, I.; Bronowicka-Szydełko, A.; Gacka, M.; Masłowski, L.; Kędzior, K.; Witkiewicz, W.; Gamian, A. Metabolites of the nitric oxide (NO) pathway are altered and indicative of reduced NO and arginine bioavailability in patients with cardiometabolic diseases complicated with chronic wounds of lower extremities: Targeted metabolomics approach (LC-MS/MS). Oxid. Med. Cell Longev. 2019, 2019, 5965721. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [Green Version]
- Krzystek-Korpacka, M.; Diakowska, D.; Bania, J.; Gamian, A. Expression stability of common housekeeping genes is differently affected by bowel inflammation and cancer: Implications for finding suitable normalizers for inflammatory bowel disease studies. Inflamm. Bowel. Dis. 2014, 20, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, K.; Bednarz-Misa, I.; Walecka-Zacharska, E.; Wierzbicki, J.; Agrawal, A.; Gamian, A.; Krzystek-Korpacka, M. Oversecretion and overexpression of nicotinamide phosphoribosyltransferase/pre-B colony-enhancing factor/visfatin in inflammatory bowel disease reflects the disease activity, severity of inflammatory response and hypoxia. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzystek-Korpacka, M.; Gorska, S.; Diakowska, D.; Kapturkiewicz, B.; Podkowik, M.; Gamian, A.; Bednarz-Misa, I. Midkine is up-regulated in both cancerous and inflamed bowel, reflecting lymph node metastasis in colorectal cancer and clinical activity of ulcerative colitis. Cytokine 2017, 89, 68–75. [Google Scholar] [CrossRef] [PubMed]








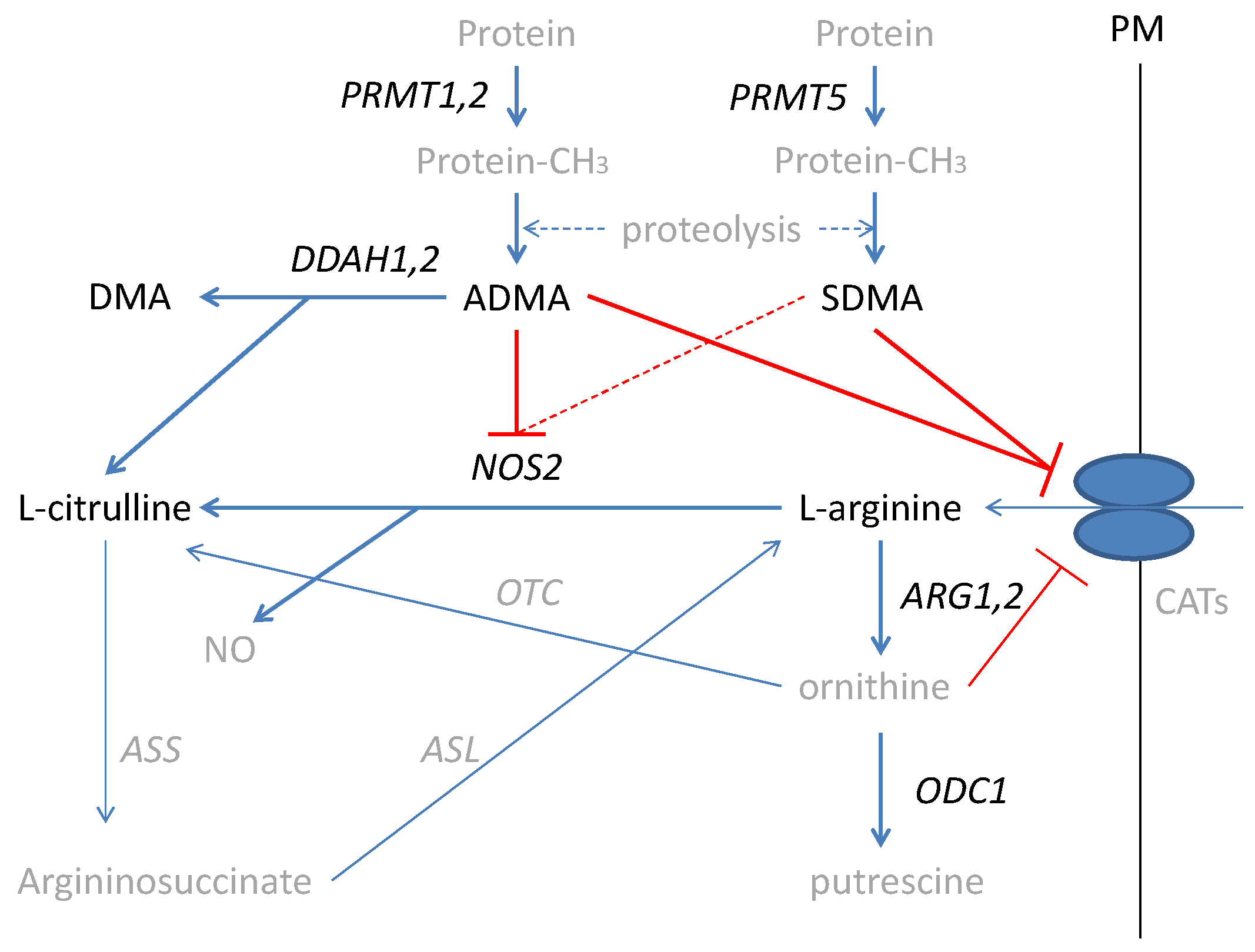{kind=link}
{kind=link}
{kind=link}
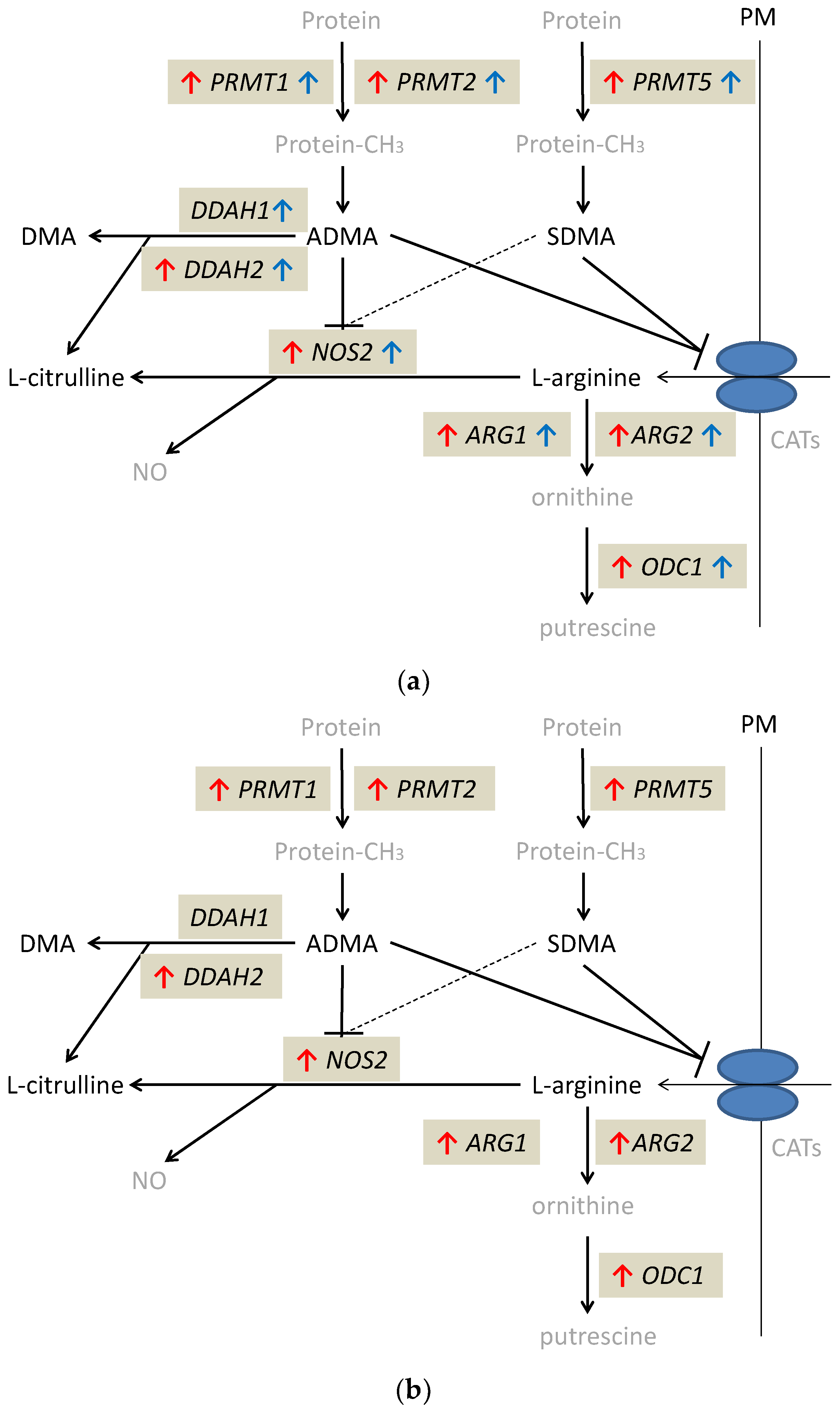{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Controls (n = 40) | IBS (n = 18) | IBD (n = 100) | p-value |
|---|---|---|---|---|
| L-arginine [µM] | 157.1 (144–177) | 142.6 (122–173) | 147.7 (137–160) | 0.084 |
| L-citrulline [µM] | 42.7 (37.9–49.8) a,b | 113.7 (65.1–153.7) b,c | 74.2 (58.1–85.4) a,c | <0.00001 |
| ADMA [µM] | 0.603 (0.57–0.64) | 0.532 (0.50–0.63) | 0.583 (0.55–0.61) | 0.280 |
| SDMA [µM] | 0.494 (0.47–0.54) a | 0.429 (0.39–0.45) b,c | 0.468 (0.45–0.49) a | 0.016 |
| DMA [µM] | 50.37 (45–53.6) a,b | 71.06 (62.5–87) c | 69.14 (63.1–74)c | <0.00001 |
| Metabolite | Controls (n = 40) | IBS (n = 18) | IBD (n = 100) | p-value | |||
|---|---|---|---|---|---|---|---|
| Active CD (n = 40) | Inactive CD (n = 12) | Active UC (n = 33) | Inactive UC (n = 15) | ||||
| L-arginine [µM] | 157.1 (144–177) a | 142.6 (122–173) | 128.5 (107–148) b,c | 167.9 (113–179) | 163.9 (143–184) a | 142.5 (129–160) | 0.006 |
| L-citrulline [µM] | 42.7 (37.9–49.8) a,c–f | 113.7 (65.1–153.7) a–c | 68.7 (54.8–90.3) b,e | 90.9 (23.5–155) b | 65.9 (34.2–86.4) b,e,f | 80.5 (56.7–148) b,c | <0.0001 |
| ADMA [µM] | 0.603 (0.57–0.64) | 0.532 (0.50–0.63) | 0.565 (0.53–0.60) | 0.600 (0.54–0.69) | 0.591 (0.55–0.63) | 0.584 (0.46–0.68) | 0.555 |
| SDMA [µM] | 0.494 (0.47–0.54) a,e | 0.429 (0.39–0.45) b–d | 0.437 (0.41–0.47) b,c | 0.485 (0.44–0.59) e | 0.520 (0.47–0.60) a,e | 0.457 (0.42–0.50) | 0.003 |
| DMA [µM] | 50.37 (45–53.6) a,c–f | 71.06 (62.5–87) b,d | 70.9 (59.7–77.9) b,d | 60.5 (49.4–72.1) a–c,e | 73.9 (65.2–77.5) b,d | 64.8 (47.1–90.3) b | <0.00001 |
| Mayo Endoscopic Score (N) | Arginine | p-value |
|---|---|---|
| 0 (10) | 139.1 (118–161) a | 0.040 |
| 1 (8) | 159.7 (132–188) | |
| 2 (9) | 163.5 (140–187) | |
| 3 (7) | 190.8 (150–232) b |
| Gene | CD_i (n = 16) | CD_q (n = 18) | UC_n (n = 17) | p-value |
|---|---|---|---|---|
| ARG1 [NRQ] | 0.60 (0.25–1.42) | 0.81 (0.36–1.82) | 0.83 (0.38–1.83) | 0.804 |
| ARG2 [NRQ] | 0.67 (0.42–1.09) a,b | 2.83 (1.84–4.34) c | 1.60 (0.83–3.09) c | 0.001 |
| DDAH1 [NRQ] | 0.92 (0.52–1.64) | 1.90 (1.21–2.99) b | 0.65 (0.30–1.42) a | 0.032 |
| DDAH2 [NRQ] | 1.05 (0.89–1.24) b | 1.08 (0.85–1.37) b | 0.70 (0.61–0.81) a,c | 0.002 |
| NOS2 [NRQ] | 1.05 (0.42–2.66) | 1.32 (0.47–3.79) | 1.50 (0.50–4.48) | 0.866 |
| ODC1 [NRQ] | 1.0 (0.82–1.22) | 1.03 (0.88–1.21) | 0.89 (0.72–1.1) | 0.484 |
| PRMT1 [NRQ] | 1.0 (0.84–1.2) | 1.21 (0.92–1.61) b | 0.81 (0.66–0.99) a | 0.036 |
| PRMT2 [NRQ] | 1.16 (0.98–1.39) b | 1.22 (0.89–1.68) b | 0.71 (0.55–0.92) a,c | 0.005 |
| PRMT5 [NRQ] | 1.01 (0.87–1.16) | 1.1 (0.94–1.3) b | 0.81 (0.66–0.98) a | 0.024 |
| Gene | CD_i (n = 9) | CD_q (n = 16) | UC_i (n = 15) | Normal (n = 20) | p-value |
|---|---|---|---|---|---|
| ARG1 [NRQ] | 0.79 (0.5–28.7) a | 1.18 (0.5–7.2) a | 8.86 (1.3–15.4) a | 0 (0–0) b–d | <0.0001 |
| ARG2 [NRQ] | 0.96 (0.6–6.4) a | 1.4 (0.8–3.0) a | 1.45 (0.7–2.4) a | 0 (0–0) b–d | <0.0001 |
| DDAH1 [NRQ] | 0.48 (0.1–2.9) c | 2.75 (0.9–4.7) a,b,d | 0.72 (0.2–2.5) c | 0.3 (0.2–0.6) c | 0.002 |
| DDAH2 [NRQ] | 1.57 (1.2–3.1) a,d | 1.54 (1.2–1.9) a,d | 0.93 (0.8–1.3) a–c | 0.31 (0.1–0.4) b–d | <0.0001 |
| NOS2 [NRQ] | 0.21 (0.05–1.2) a,d | 0.74 (0.2–1.5) a,d | 7.96 (1–24.2) a–c | 0.07 (0.04–0.1) b–d | <0.0001 |
| ODC1 [NRQ] | 1.27 (0.9–2.1) a,c | 2 (1.6–2.3) a,b | 1.72 (1.4–2.3) a | 0.02 (0.01–0.03) b–d | <0.0001 |
| PRMT1 [NRQ] | 1.68 (0.8–3.7) a | 1.76 (1.3–2) a | 1.33 (1.2–1.8) a | 0.08 (0.05–0.2) b–d | <0.0001 |
| PRMT2 [NRQ] | 1.55 (1–5.4) a | 1.7 (1.4–3.1) a,d | 1.42 (0.9–1.7) a,c | 0 (0–0) b–d | <0.0001 |
| PRMT5 [NRQ] | 1.09 (0.9–3) a | 1.7 (1.5–2) a | 1.54 (1.4–1.8) a | 0.07 (0–0.2) b–d | <0.0001 |
| Parameter | Controls | IBS | IBD | p-value 1 | IBD | p-value 2 | |||
|---|---|---|---|---|---|---|---|---|---|
| Active CD | Inactive CD | Active UC | Inactive UC | ||||||
| N | 40 | 18 | 100 | 40 | 12 | 34 | 14 | ||
| Sex (F/M) | 20/20 | 10/8 | 41/59 | 0.394 3 | 19/21 | 4/8 | 11/22 | 7/8 | 0.577 3 |
| Age [yrs.], mean ± SD | 35.9 ± 11.5 | 41.1 ± 14.5 | 39.7 ± 14.4 | 0.251 4 | 36.4 ± 13.7 | 42.9 ± 14.3 | 42.4 ± 15.3 | 40.2 ± 13.4 | 0.215 4 |
| Patients with active disease [%] | 77 | 71 | 0.504 5 | ||||||
| Activity, mean ± SD | CDAI: 259 ± 89 | RI: 10.4 ± 5 | |||||||
| Parameter | Small Bowel | p-value | ||
| UC_n | CD_q | CD_i | ||
| N | 17 | 18 | 16 | |
| Sex (F/M) | 9/8 | 7/11 | 6/10 | 0.605 1 |
| Age [yrs.], mean ± SD | 36.7 ± 11.9 | 32.9 ± 6.1 | 31.9 ± 6 | 0.232 2 |
| Large Bowel | ||||
| CD_q | CD_i | UC_i | ||
| N | 16 | 9 | 15 | |
| Sex (F/M) | 5/11 | 3/6 | 8/7 | 0.409 1 |
| Age [yrs.], mean ± SD | 31.9 ± 6.4 | 31.3 ± 9.3 | 37.3 ± 12 | 0.204 2 |
| Symbol | Encoded Protein | Accession No. | Primer Sequence 5′→3′ | Amp. Size [bp] |
|---|---|---|---|---|
| PPIA 1 | Peptidylprolyl isomerase A | NM_021130.3 | F: ggcaaatgctggacccaacaca R: tgctggtcttgccattcctgga | 161 |
| RPLP0 1 | Ribosomal protein, large, P0 | NM_001002.3 | F: tcacaacaagcataccaagaagc R: gtatccgatgtccacaatgtcaag | 263 |
| ARG1 1 | Arginase-1 | NM_001244438.2 | F: tcatctgggtggatgctcacac R: gagaatcctggcacatcgggaa | 130 |
| ARG2 1 | Arginase-2 | NM_001172.4 | F: ctggcttgatgaaaaggctctcc R: tgagcgtggattcactatcaggt | 119 |
| NOS2 1 | Inducible nitric oxide synthase | NM_000625.4 | F: gctctacacctccaatgtgacc R: ctgccgagatttgagcctcatg | 136 |
| PRMT1 1 | Arginine N-methyltransferase-1 | NM_001536.5 | F: tgcggtgaagatcgtcaaagcc R: ggactcgtagaagaggcagtag | 142 |
| PRMT2 1 | Arginine N-methyltransferase-2 | NM_206962.4 | F: gcagttggacatgagaaccgtg R: aggctctggaagtggacgctaa | 129 |
| PRMT5 1 | Arginine N-methyltransferase-5 | NM_006109.5 | F: ctagaccgagtaccagaagagg R: cagcatacagctttatccgccg | 136 |
| DDAH1 1 | Dimethylarginine dimethylaminohydrolase-1 | NM_012137.4 | F: atgcagtctccacagtgccagt R: ttgtcgtagcggtggtcactca | 151 |
| DDAH2 1 | Dimethylarginine dimethylaminohydrolase-2 | NM_001303007.2 | F: ctttcttcgtcctgggttgcct R: ctccagttctgagcaggacaca | 136 |
| ODC1 1 | Ornithine decarboxylase-1 | NM_002539.3 | F: ccaaagcagtctgtcgtctcag R: cagagattgcctgcacgaaggt | 162 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzystek-Korpacka, M.; G. Fleszar, M.; Bednarz-Misa, I.; Lewandowski, Ł.; Szczuka, I.; Kempiński, R.; Neubauer, K. Transcriptional and Metabolomic Analysis of L-Arginine/Nitric Oxide Pathway in Inflammatory Bowel Disease and Its Association with Local Inflammatory and Angiogenic Response: Preliminary Findings. Int. J. Mol. Sci. 2020, 21, 1641. https://doi.org/10.3390/ijms21051641
Krzystek-Korpacka M, G. Fleszar M, Bednarz-Misa I, Lewandowski Ł, Szczuka I, Kempiński R, Neubauer K. Transcriptional and Metabolomic Analysis of L-Arginine/Nitric Oxide Pathway in Inflammatory Bowel Disease and Its Association with Local Inflammatory and Angiogenic Response: Preliminary Findings. International Journal of Molecular Sciences. 2020; 21(5):1641. https://doi.org/10.3390/ijms21051641
Chicago/Turabian StyleKrzystek-Korpacka, Małgorzata, Mariusz G. Fleszar, Iwona Bednarz-Misa, Łukasz Lewandowski, Izabela Szczuka, Radosław Kempiński, and Katarzyna Neubauer. 2020. "Transcriptional and Metabolomic Analysis of L-Arginine/Nitric Oxide Pathway in Inflammatory Bowel Disease and Its Association with Local Inflammatory and Angiogenic Response: Preliminary Findings" International Journal of Molecular Sciences 21, no. 5: 1641. https://doi.org/10.3390/ijms21051641
APA StyleKrzystek-Korpacka, M., G. Fleszar, M., Bednarz-Misa, I., Lewandowski, Ł., Szczuka, I., Kempiński, R., & Neubauer, K. (2020). Transcriptional and Metabolomic Analysis of L-Arginine/Nitric Oxide Pathway in Inflammatory Bowel Disease and Its Association with Local Inflammatory and Angiogenic Response: Preliminary Findings. International Journal of Molecular Sciences, 21(5), 1641. https://doi.org/10.3390/ijms21051641