Serotonin in Animal Cognition and Behavior

 , ,
, ,  , and
, and 
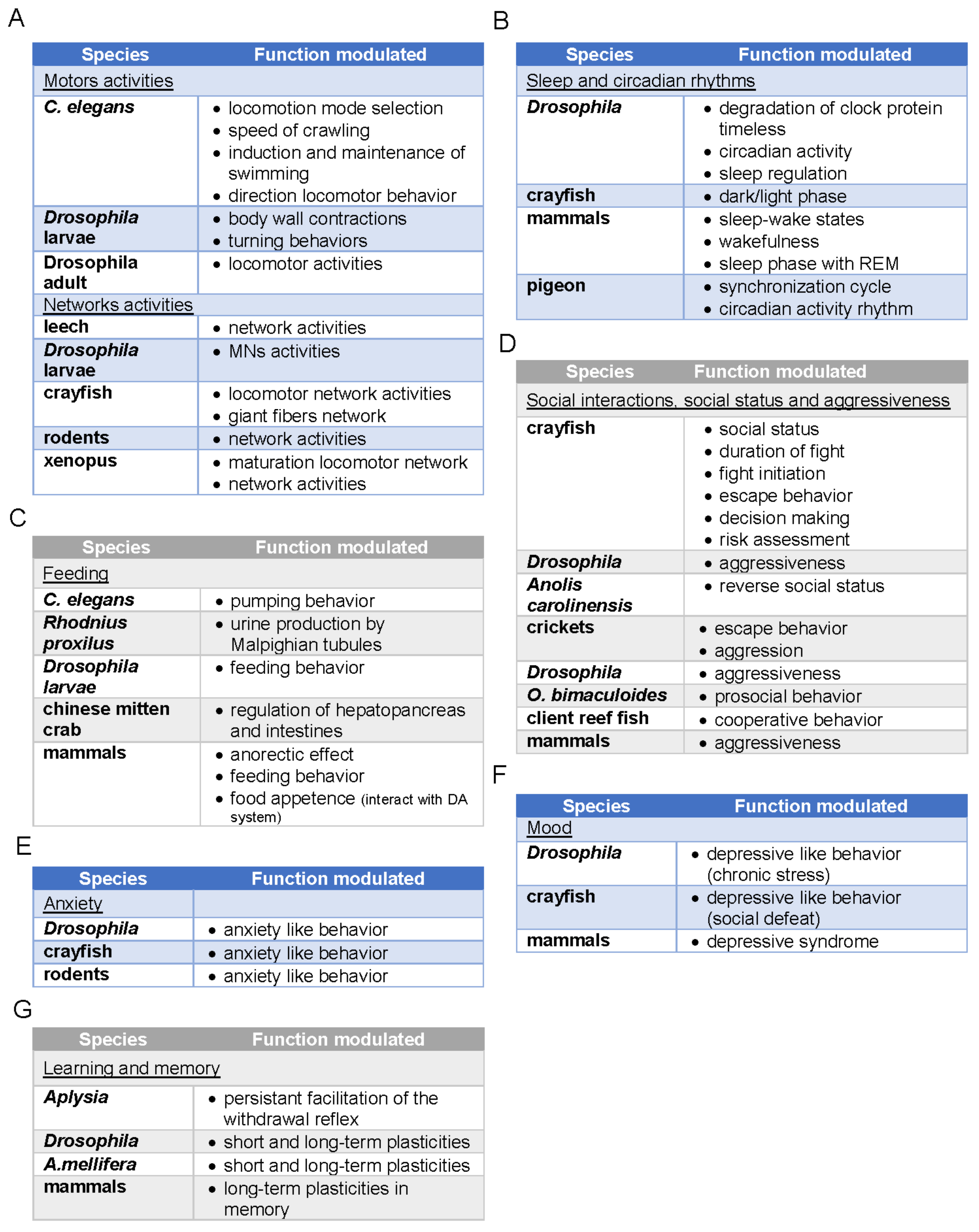{kind=link}
Abstract
:1. Introduction
2. Organization of 5-HT Systems in Animals
3. Motor activity and Locomotion
4. Arousal
5. Sleep and Circadian Rhythm
6. Feeding
7. Social Interactions, Social Status, and/or Aggressiveness
8. Anxiety
9. Mood
10. Learning and Memory
11. Impulsive/Compulsive Dimension and Behavioral Flexibility
12. Discussion
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-HT | Serotonin |
| 5-HTR | Serotonin receptors |
| 5-HTP | 5-hydroxytryptamin |
| CNS | Central nervous system |
| MAO | Monoamine oxidase |
| MBs | Mushroom Bodies |
| SSRI | Selective serotonin reuptake inhibitor |
| SERT | Serotonin transporter |
References
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, P.W.; Bharwani, A.; Lee, K.R.; Fox, M.; Thomson, J.A., Jr. Is serotonin an upper or a downer? The evolution of the serotonergic system and its role in depression and the antidepressant response. Neurosci. Biobehav. Rev. 2015, 51, 164–188. [Google Scholar] [CrossRef] [PubMed]
- Blier, P.; de Montigny, C.; Chaput, Y. A role for the serotonin system in the mechanism of action of antidepressant treatments: Preclinical evidence. J. Clin. Psychiatry 1990, 51, 14–20, discussion 21. [Google Scholar] [PubMed]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Soubrié, P. Reconciling the role of central serotonin neurons in human and animal behavior. Behav. Brain Sci. 1986, 9, 319–335. [Google Scholar] [CrossRef]
- Bari, A.; Robbins, T.W. Inhibition and impulsivity: Behavioral and neural basis of response control. Prog. Neurobiol. 2013, 108, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Lillesaar, C. Probing the diversity of serotonin neurons. Philos. Trans. R.Soc. Lond. Ser. B Biol. Sci. 2012, 367, 2382–2394. [Google Scholar] [CrossRef] [Green Version]
- Alekseyenko, O.V.; Chan, Y.B.; Fernandez, M.P.; Bulow, T.; Pankratz, M.J.; Kravitz, E.A. Single serotonergic neurons that modulate aggression in Drosophila. Curr. Biol. CB 2014, 24, 2700–2707. [Google Scholar] [CrossRef] [Green Version]
- De-Miguel, F.F.; Trueta, C. Synaptic and extrasynaptic secretion of serotonin. Cell. Mol. Neurobiol. 2005, 25, 297–312. [Google Scholar] [CrossRef]
- Dahlström, A.; Fuxe, K. Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand. 1964, 62, 1–55. [Google Scholar]
- Lidov, H.G.; Molliver, M.E. Immunohistochemical study of the development of serotonergic neurons in the rat CNS. Brain Res. Bull. 1982, 9, 559–604. [Google Scholar] [CrossRef]
- Hale, M.W.; Lowry, C.A. Functional topography of midbrain and pontine serotonergic systems: Implications for synaptic regulation of serotonergic circuits. Psychopharmacology 2011, 213, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Steinbusch, H.W. Serotonin-immunoreactive neurons and their projections in the CNS. In Handbook of Chemical Neuroanatomy – Classical Transmitters and Transmitters Receptors in the CNS Part II; Björklund, A.H.T., Kuhar, M.J., Eds.; AAN Publications: Amsterdam, The Netherlands, 1984; pp. 68–125. [Google Scholar]
- Walker, E.P.; Tadi, P. Neuroanatomy, Nucleus Raphe. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2020. [Google Scholar]
- Weissbourd, B.; Ren, J.; DeLoach, K.E.; Guenthner, C.J.; Miyamichi, K.; Luo, L. Presynaptic partners of dorsal raphe serotonergic and GABAergic neurons. Neuron 2014, 83, 645–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, X.; Wang, B.; Zhang, J.; Yan, T.; Yang, W.; Jiao, F.; Liu, J.; Wang, S. Localization of serotonin/tryptophan-hydroxylase-immunoreactive cells in the brain and suboesophageal ganglion of Drosophila melanogaster. Cell Tissue Res. 2010, 340, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wurden, S.; Homberg, U. Immunocytochemical mapping of serotonin and neuropeptides in the accessory medulla of the locust, Schistocerca gregaria. J. Comp. Neurol. 1995, 362, 305–319. [Google Scholar] [CrossRef]
- Antonsen, B.L.; Paul, D.H. Serotonergic and octopaminergic systems in the squat lobster Munida quadrispina (Anomura, Galatheidae). J. Comp. Neurol. 2001, 439, 450–468. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Acevedo, N.; Reyes-Colon, D.; Ruiz-Rodriguez, E.A.; Rivera, N.M.; Rosenthal, J.; Kohn, A.B.; Moroz, L.L.; Sosa, M.A. Cloning and immunoreactivity of the 5-HT 1Mac and 5-HT 2Mac receptors in the central nervous system of the freshwater prawn Macrobrachium rosenbergii. J. Comp. Neurol. 2009, 513, 399–416. [Google Scholar] [CrossRef] [Green Version]
- Real, D.; Czternasty, G. Mapping of serotonin-like immunoreactivity in the ventral nerve cord of crayfish. Brain Res. 1990, 521, 203–212. [Google Scholar] [CrossRef]
- Issa, F.A.; Drummond, J.; Cattaert, D.; Edwards, D.H. Neural circuit reconfiguration by social status. J. Neurosci. 2012, 32, 5638–5645. [Google Scholar] [CrossRef]
- Umbriaco, D.; Anctil, M.; Descarries, L. Serotonin-immunoreactive neurons in the cnidarian Renilla koellikeri. J. Comp. Neurol. 1990, 291, 167–178. [Google Scholar] [CrossRef]
- Daws, L.C. Unfaithful neurotransmitter transporters: Focus on serotonin uptake and implications for antidepressant efficacy. Pharmacol. Ther. 2009, 121, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elekes, K.; Hustert, R. The efferent innervation of the genital chamber by an identified serotonergic neuron in the female cricket Acheta domestica. Cell Tissue Res. 1988, 252, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.J.; Peter, J.U.; Bashammakh, S.; Hortnagl, H.; Voits, M.; Fink, H.; Bader, M. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science 2003, 299, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Beaulieu, J.M.; Sotnikova, T.D.; Gainetdinov, R.R.; Caron, M.G. Tryptophan hydroxylase-2 controls brain serotonin synthesis. Science 2004, 305, 217. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.M.; Neckameyer, W.S. Serotonin synthesis by two distinct enzymes in Drosophila melanogaster. Arch. Insect. Biochem. Physiol. 2005, 59, 12–31. [Google Scholar] [CrossRef] [PubMed]
- De Deurwaerdere, P.; Binda, C.; Corne, R.; Leone, C.; Valeri, A.; Valoti, M.; Ramsay, R.R.; Fall, Y.; Marco-Contelles, J. Comparative Analysis of the Neurochemical Profile and MAO Inhibition Properties of N-(Furan-2-ylmethyl)-N-methylprop-2-yn-1-amine. ACS Chem. Neurosci. 2017, 8, 1026–1035. [Google Scholar] [CrossRef] [Green Version]
- Finberg, J.P. Update on the pharmacology of selective inhibitors of MAO-A and MAO-B: Focus on modulation of CNS monoamine neurotransmitter release. Pharmacol. Ther. 2014, 143, 133–152. [Google Scholar] [CrossRef]
- Di Giovanni, G.; Svob Strac, D.; Sole, M.; Unzeta, M.; Tipton, K.F.; Muck-Seler, D.; Bolea, I.; Della Corte, L.; Nikolac Perkovic, M.; Pivac, N.; et al. Monoaminergic and Histaminergic Strategies and Treatments in Brain Diseases. Front. Neurosci. 2016, 10, 541. [Google Scholar] [CrossRef] [Green Version]
- Fossat, P.; Bacque-Cazenave, J.; De Deurwaerdere, P.; Cattaert, D.; Delbecque, J.P. Serotonin, but not dopamine, controls the stress response and anxiety-like behavior in the crayfish Procambarus clarkii. J. Exp. Biol. 2015, 218, 2745–2752. [Google Scholar] [CrossRef] [Green Version]
- Fossat, P.; Bacque-Cazenave, J.; De Deurwaerdere, P.; Delbecque, J.P.; Cattaert, D. Comparative behavior. Anxiety-like behavior in crayfish is controlled by serotonin. Science 2014, 344, 1293–1297. [Google Scholar] [CrossRef]
- Chagraoui, A.; Whitestone, S.; Baassiri, L.; Manem, J.; Di Giovanni, G.; De Deurwaerdere, P. Neurochemical impact of the 5-HT2C receptor agonist WAY-163909 on monoamine tissue content in the rat brain. Neurochem. Int. 2019, 124, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Fitoussi, A.; Dellu-Hagedorn, F.; De Deurwaerdere, P. Monoamines tissue content analysis reveals restricted and site-specific correlations in brain regions involved in cognition. Neuroscience 2013, 255, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Hyacinthe, C.; De Deurwaerdere, P.; Thiollier, T.; Li, Q.; Bezard, E.; Ghorayeb, I. Blood withdrawal affects iron store dynamics in primates with consequences on monoaminergic system function. Neuroscience 2015, 290, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Pifl, C.; Schingnitz, G.; Hornykiewicz, O. Effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine on the regional distribution of brain monoamines in the rhesus monkey. Neuroscience 1991, 44, 591–605. [Google Scholar] [CrossRef]
- Puginier, E.; Bharatiya, R.; Chagraoui, A.; Manem, J.; Cho, Y.H.; Garret, M.; De Deurwaerdere, P. Early neurochemical modifications of monoaminergic systems in the R6/1 mouse model of Huntington’s disease. Neurochem. Int. 2019, 128, 186–195. [Google Scholar] [CrossRef]
- Kops, M.S.; Kjaer, J.B.; Gunturkun, O.; Westphal, K.G.C.; Korte-Bouws, G.A.H.; Olivier, B.; Korte, S.M.; Bolhuis, J.E. Brain monoamine levels and behaviour of young and adult chickens genetically selected on feather pecking. Behav. Brain Res. 2017, 327, 11–20. [Google Scholar] [CrossRef]
- De Abreu, M.S.; Messias, J.P.M.; Thornqvist, P.O.; Winberg, S.; Soares, M.C. The variable monoaminergic outcomes of cleaner fish brains when facing different social and mutualistic contexts. PeerJ 2018, 6, e4830. [Google Scholar] [CrossRef] [Green Version]
- Klouche, M.S.; De Deurwaerdere, P.; Dellu-Hagedorn, F.; Lakhdar-Ghazal, N.; Benomar, S. Monoamine content during the reproductive cycle of Perna perna depends on site of origin on the Atlantic Coast of Morocco. Sci. Rep. 2015, 5, 13715. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 2002, 71, 533–554. [Google Scholar] [CrossRef]
- Peroutka, S.J.; Howell, T.A. The molecular evolution of G protein-coupled receptors: Focus on 5-hydroxytryptamine receptors. Neuropharmacology 1994, 33, 319–324. [Google Scholar] [CrossRef]
- Saudou, F.; Hen, R. 5-Hydroxytryptamine receptor subtypes in vertebrates and invertebrates. Neurochem. Int. 1994, 25, 503–532. [Google Scholar] [CrossRef]
- Clark, M.C.; Dever, T.E.; Dever, J.J.; Xu, P.; Rehder, V.; Sosa, M.A.; Baro, D.J. Arthropod 5-HT2 receptors: A neurohormonal receptor in decapod crustaceans that displays agonist independent activity resulting from an evolutionary alteration to the DRY motif. J. Neurosci. 2004, 24, 3421–3435. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, N.; Edwards, D.H.; Baro, D.J. Conservation of structure, signaling and pharmacology between two serotonin receptor subtypes from decapod crustaceans, Panulirus interruptus and Procambarus clarkii. J. Exp. Biol. 2008, 211, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Huang, G.; Xu, M.; Zhang, C.; Cheng, Y. Molecular cloning and functional expression of the 5-HT7 receptor in Chinese mitten crab (Eriocheir sinensis). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2018, 226, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Straub, V.A.; Benjamin, P.R. Extrinsic modulation and motor pattern generation in a feeding network: A cellular study. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 1767–1778. [Google Scholar] [CrossRef] [Green Version]
- Yeoman, M.S.; Brierley, M.J.; Benjamin, P.R. Central pattern generator interneurons are targets for the modulatory serotonergic cerebral giant cells in the feeding system of Lymnaea. J. Neurophysiol. 1996, 75, 11–25. [Google Scholar] [CrossRef]
- Lee, Y.S.; Choi, S.L.; Lee, S.H.; Kim, H.; Park, H.; Lee, N.; Lee, S.H.; Chae, Y.S.; Jang, D.J.; Kandel, E.R.; et al. Identification of a serotonin receptor coupled to adenylyl cyclase involved in learning-related heterosynaptic facilitation in Aplysia. Proc. Natl. Acad. Sci. USA 2009, 106, 14634–14639. [Google Scholar] [CrossRef] [Green Version]
- Mapara, S.; Parries, S.; Quarrington, C.; Ahn, K.C.; Gallin, W.J.; Goldberg, J.I. Identification, molecular structure and expression of two cloned serotonin receptors from the pond snail, Helisoma trivolvis. J. Exp. Biol. 2008, 211, 900–910. [Google Scholar] [CrossRef] [Green Version]
- Sosa, M.A.; Spitzer, N.; Edwards, D.H.; Baro, D.J. A crustacean serotonin receptor: Cloning and distribution in the thoracic ganglia of crayfish and freshwater prawn. J. Comp. Neurol. 2004, 473, 526–537. [Google Scholar] [CrossRef]
- Bacque-Cazenave, J.; Issa, F.A.; Edwards, D.H.; Cattaert, D. Spatial segregation of excitatory and inhibitory effects of 5-HT on crayfish motoneurons. J. Neurophysiol. 2013, 109, 2793–2802. [Google Scholar] [CrossRef] [Green Version]
- Tierney, A.J. Invertebrate serotonin receptors: A molecular perspective on classification and pharmacology. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef] [PubMed]
- Vleugels, R.; Lenaerts, C.; Baumann, A.; Vanden Broeck, J.; Verlinden, H. Pharmacological characterization of a 5-HT1-type serotonin receptor in the red flour beetle, Tribolium castaneum. PLoS ONE 2013, 8, e65052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troppmann, B.; Balfanz, S.; Baumann, A.; Blenau, W. Inverse agonist and neutral antagonist actions of synthetic compounds at an insect 5-HT1 receptor. Br. J. Pharmacol. 2010, 159, 1450–1462. [Google Scholar] [CrossRef] [Green Version]
- Di Giovanni, G.C.A.; Bharatiya, R.; De Deurwaerdère, P. Serotonergic control of excitability: From neuron to networks. In Handbook of the Behavioral Neurobiology of Serotonin; Muller, C.C.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 31. [Google Scholar]
- Patocka, N.; Sharma, N.; Rashid, M.; Ribeiro, P. Serotonin signaling in Schistosoma mansoni: A serotonin-activated G protein-coupled receptor controls parasite movement. PLoS Pathog. 2014, 10, e1003878. [Google Scholar] [CrossRef]
- Bielecki, J.; Nachman, G.; Garm, A. Swim pacemaker response to bath applied neurotransmitters in the cubozoan Tripedalia cystophora. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2013, 199, 785–797. [Google Scholar] [CrossRef]
- Ranganathan, R.; Cannon, S.C.; Horvitz, H.R. MOD-1 is a serotonin-gated chloride channel that modulates locomotory behaviour in C. elegans. Nature 2000, 408, 470–475. [Google Scholar] [CrossRef]
- Crisp, K.M.; Mesce, K.A. To swim or not to swim: Regional effects of serotonin, octopamine and amine mixtures in the medicinal leech. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2003, 189, 461–470. [Google Scholar] [CrossRef]
- Veraszto, C.; Ueda, N.; Bezares-Calderon, L.A.; Panzera, A.; Williams, E.A.; Shahidi, R.; Jekely, G. Ciliomotor circuitry underlying whole-body coordination of ciliary activity in the Platynereis larva. eLife 2017, 6. [Google Scholar] [CrossRef]
- Kinney, M.P.; Panting, N.D.; Clark, T.M. Modulation of appetite and feeding behavior of the larval mosquito Aedes aegypti by the serotonin-selective reuptake inhibitor paroxetine: Shifts between distinct feeding modes and the influence of feeding status. J. Exp. Biol. 2014, 217, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Majeed, Z.R.; Abdeljaber, E.; Soveland, R.; Cornwell, K.; Bankemper, A.; Koch, F.; Cooper, R.L. Modulatory Action by the Serotonergic System: Behavior and Neurophysiology in Drosophila melanogaster. Neural Plast. 2016, 2016, 7291438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, S.R.; Guilhermino, L.; Guimaraes, L. Biochemical and locomotor responses of Carcinus maenas exposed to the serotonin reuptake inhibitor fluoxetine. Chemosphere 2011, 85, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Parsons, D.W.; Pinsker, H.M. Swimming in Aplysia brasiliana: Behavioral and cellular effects of serotonin. J. Neurophysiol. 1989, 62, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, J.M.; Katz, P.S. Different functions for homologous serotonergic interneurons and serotonin in species-specific rhythmic behaviours. Proc. Biol. Sci. 2009, 276, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.L.; Lyons, D.E.; Meekins, T.L.; Newcomb, J.M. Serotonin influences locomotion in the nudibranch mollusc Melibe leonina. Biol. Bull. 2011, 220, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Dyakonova, V.; Hernadi, L.; Ito, E.; Dyakonova, T.; Zakharov, I.; Sakharov, D. The activity of isolated snail neurons controlling locomotion is affected by glucose. Biophysics 2015, 11, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Fong, P.P.; Bury, T.B.; Donovan, E.E.; Lambert, O.J.; Palmucci, J.R.; Adamczak, S.K. Exposure to SSRI-type antidepressants increases righting time in the marine snail Ilyanassa obsoleta. Environ. Sci. Pollut. Res. Int. 2017, 24, 725–731. [Google Scholar] [CrossRef]
- Di Poi, C.; Bidel, F.; Dickel, L.; Bellanger, C. Cryptic and biochemical responses of young cuttlefish Sepia officinalis exposed to environmentally relevant concentrations of fluoxetine. Aquat. Toxicol. 2014, 151, 36–45. [Google Scholar] [CrossRef]
- Fong, P.P.; Ford, A.T. The biological effects of antidepressants on the molluscs and crustaceans: A review. Aquat. Toxicol. 2014, 151, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Camicia, F.; Herz, M.; Prada, L.C.; Kamenetzky, L.; Simonetta, S.H.; Cucher, M.A.; Bianchi, J.I.; Fernandez, C.; Brehm, K.; Rosenzvit, M.C. The nervous and prenervous roles of serotonin in Echinococcus spp. Int. J. Parasitol. 2013, 43, 647–659. [Google Scholar] [CrossRef]
- Vidal-Gadea, A.; Topper, S.; Young, L.; Crisp, A.; Kressin, L.; Elbel, E.; Maples, T.; Brauner, M.; Erbguth, K.; Axelrod, A.; et al. Caenorhabditis elegans selects distinct crawling and swimming gaits via dopamine and serotonin. Proc. Natl. Acad. Sci. USA 2011, 108, 17504–17509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, T.; Osada, T.; Shingai, R. Serotonin deficiency shortens the duration of forward movement in Caenorhabditis elegans. Biosci. Biotechnol. Biochem. 2005, 69, 1767–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawin, E.R.; Ranganathan, R.; Horvitz, H.R. C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 2000, 26, 619–631. [Google Scholar] [CrossRef] [Green Version]
- Gurel, G.; Gustafson, M.A.; Pepper, J.S.; Horvitz, H.R.; Koelle, M.R. Receptors and other signaling proteins required for serotonin control of locomotion in Caenorhabditis elegans. Genetics 2012, 192, 1359–1371. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, A.; Takagi, S. An optogenetic application of proton pump ArchT to C. elegans cells. Neurosci. Res. 2013, 75, 29–34. [Google Scholar] [CrossRef]
- Okusawa, S.; Kohsaka, H.; Nose, A. Serotonin and downstream leucokinin neurons modulate larval turning behavior in Drosophila. J. Neurosci. 2014, 34, 2544–2558. [Google Scholar] [CrossRef]
- Wu, W.H.; Cooper, R.L. Serotonin and synaptic transmission at invertebrate neuromuscular junctions. Exp. Neurobiol. 2012, 21, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Silva, B.; Goles, N.I.; Varas, R.; Campusano, J.M. Serotonin receptors expressed in Drosophila mushroom bodies differentially modulate larval locomotion. PLoS ONE 2014, 9, e89641. [Google Scholar] [CrossRef] [Green Version]
- Harris-Warrick, R.M.; Cohen, A.H. Serotonin modulates the central pattern generator for locomotion in the isolated lamprey spinal cord. J. Exp. Biol. 1985, 116, 27–46. [Google Scholar]
- Brustein, E.; Chong, M.; Holmqvist, B.; Drapeau, P. Serotonin patterns locomotor network activity in the developing zebrafish by modulating quiescent periods. J. Neurobiol. 2003, 57, 303–322. [Google Scholar] [CrossRef]
- Gabriel, J.P.; Mahmood, R.; Kyriakatos, A.; Soll, I.; Hauptmann, G.; Calabrese, R.L.; El Manira, A. Serotonergic modulation of locomotion in zebrafish: Endogenous release and synaptic mechanisms. J. Neurosci. 2009, 29, 10387–10395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, J.E.; Wahlstrom-Helgren, S.; Wiggin, T.D.; Corwin, B.M.; Lillesaar, C.; Masino, M.A. Intraspinal serotonergic signaling suppresses locomotor activity in larval zebrafish. Dev. Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sillar, K.T.; Reith, C.A.; McDearmid, J.R. Development and aminergic neuromodulation of a spinal locomotor network controlling swimming in Xenopus larvae. Ann. N. Y. Acad. Sci. 1998, 860, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Rauscent, A.; Einum, J.; Le Ray, D.; Simmers, J.; Combes, D. Opposing aminergic modulation of distinct spinal locomotor circuits and their functional coupling during amphibian metamorphosis. J. Neurosci. 2009, 29, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Viala, D.; Buser, P. Effects of a decarboxylase inhibitor on the Dopa and 5-HTP induced changes in the locomotor-like discharge pattern of rabbit hind limb nerves. Psychopharmacologia 1974, 40, 225–233. [Google Scholar] [CrossRef]
- Cazalets, J.R.; Sqalli-Houssaini, Y.; Clarac, F. Activation of the central pattern generators for locomotion by serotonin and excitatory amino acids in neonatal rat. J. Physiol. 1992, 455, 187–204. [Google Scholar] [CrossRef]
- Nishimaru, H.; Takizawa, H.; Kudo, N. 5-Hydroxytryptamine-induced locomotor rhythm in the neonatal mouse spinal cord in vitro. Neurosci. Lett. 2000, 280, 187–190. [Google Scholar] [CrossRef]
- Antri, M.; Barthe, J.Y.; Mouffle, C.; Orsal, D. Long-lasting recovery of locomotor function in chronic spinal rat following chronic combined pharmacological stimulation of serotonergic receptors with 8-OHDPAT and quipazine. Neurosci. Lett. 2005, 384, 162–167. [Google Scholar] [CrossRef]
- Gimenez y Ribotta, M.; Orsal, D.; Feraboli-Lohnherr, D.; Privat, A.; Provencher, J.; Rossignol, S. Kinematic analysis of recovered locomotor movements of the hindlimbs in paraplegic rats transplanted with monoaminergic embryonic neurons. Ann. N. Y. Acad. Sci. 1998, 860, 521–523. [Google Scholar] [CrossRef]
- Slawinska, U.; Miazga, K.; Jordan, L.M. The role of serotonin in the control of locomotor movements and strategies for restoring locomotion after spinal cord injury. Acta Neurobiol. Exp. 2014, 74, 172–187. [Google Scholar]
- Ballion, B.; Morin, D.; Viala, D. Forelimb locomotor generators and quadrupedal locomotion in the neonatal rat. Eur. J. Neurosci. 2001, 14, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Norreel, J.C.; Pflieger, J.F.; Pearlstein, E.; Simeoni-Alias, J.; Clarac, F.; Vinay, L. Reversible disorganization of the locomotor pattern after neonatal spinal cord transection in the rat. J. Neurosci. 2003, 23, 1924–1932. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jordan, L.M. Stimulation of the parapyramidal region of the neonatal rat brain stem produces locomotor-like activity involving spinal 5-HT7 and 5-HT2A receptors. J. Neurophysiol. 2005, 94, 1392–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beliez, L.; Barriere, G.; Bertrand, S.S.; Cazalets, J.R. Origin of thoracic spinal network activity during locomotor-like activity in the neonatal rat. J. Neurosci. 2015, 35, 6117–6130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oueghlani, Z.; Juvin, L.; Lambert, F.M.; Cardoit, L.; Courtand, G.; Masmejean, F.; Cazalets, J.R.; Barriere, G. Serotonergic modulation of sacral dorsal root stimulation-induced locomotor output in newborn rat. Neuropharmacology 2019. [Google Scholar] [CrossRef]
- Perrier, J.F.; Cotel, F. Serotonergic modulation of spinal motor control. Curr. Opin. Neurobiol. 2015, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Harvey, P.J.; Li, X.; Li, Y.; Bennett, D.J. 5-HT2 receptor activation facilitates a persistent sodium current and repetitive firing in spinal motoneurons of rats with and without chronic spinal cord injury. J. Neurophysiol. 2006, 96, 1158–1170. [Google Scholar] [CrossRef] [Green Version]
- Dasari, S.; Cooper, R.L. Modulation of sensory-CNS-motor circuits by serotonin, octopamine, and dopamine in semi-intact Drosophila larva. Neurosci. Res. 2004, 48, 221–227. [Google Scholar] [CrossRef]
- Angstadt, J.D.; Grassmann, J.L.; Theriault, K.M.; Levasseur, S.M. Mechanisms of postinhibitory rebound and its modulation by serotonin in excitatory swim motor neurons of the medicinal leech. J. Comp. Physiol. A Neuroetho.l Sens. Neural Behav. Physiol. 2005, 191, 715–732. [Google Scholar] [CrossRef]
- Bacque-Cazenave, J.; Fossat, P.; Issa, F.A.; Edwards, D.H.; Delbecque, J.P.; Cattaert, D. Duality of 5-HT Effects on Crayfish Motoneurons. Front. Physiol. 2019, 10, 1280. [Google Scholar] [CrossRef]
- Lee, S.H.; Taylor, K.; Krasne, F.B. Reciprocal stimulation of decay between serotonergic facilitation and depression of synaptic transmission. J. Neurophysiol. 2008, 100, 1113–1126. [Google Scholar] [CrossRef] [Green Version]
- Komuniecki, R.W.; Hobson, R.J.; Rex, E.B.; Hapiak, V.M.; Komuniecki, P.R. Biogenic amine receptors in parasitic nematodes: What can be learned from Caenorhabditis elegans? Mol. Biochem. Parasitol. 2004, 137, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fischer, L.; Florey, E. Modulation of synaptic transmission and excitation-contraction coupling in the opener muscle of the crayfish, Astacus leptodactylus, by 5-hydroxytryptamine and octopamine. J. Exp. Biol. 1983, 102, 187–198. [Google Scholar]
- Strawn, J.R.; Neckameyer, W.S.; Cooper, R.L. The effects of 5-HT on sensory, central and motor neurons driving the abdominal superficial flexor muscles in the crayfish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 127, 533–550. [Google Scholar] [CrossRef]
- Fox, L.E.; Lloyd, P.E. Mechanisms involved in persistent facilitation of neuromuscular synapses in aplysia. J. Neurophysiol. 2002, 87, 2018–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotshaw, D.P.; Lloyd, P.E. Peptidergic and serotonergic facilitation of a neuromuscular synapse in Aplysia. Brain Res. 1990, 526, 81–94. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Fornal, C.A. Activity of serotonergic neurons in behaving animals. Neuropsychopharmacology 1999, 21, 9S–15S. [Google Scholar] [CrossRef]
- Severson, C.A.; Wang, W.; Pieribone, V.A.; Dohle, C.I.; Richerson, G.B. Midbrain serotonergic neurons are central pH chemoreceptors. Nat. Neurosci. 2003, 6, 1139–1140. [Google Scholar] [CrossRef]
- Jing, J.; Gillette, R.; Weiss, K.R. Evolving concepts of arousal: Insights from simple model systems. Rev. Neurosci. 2009, 20, 405–427. [Google Scholar] [CrossRef]
- Nall, A.; Sehgal, A. Monoamines and sleep in Drosophila. Behav. Neurosci. 2014, 128, 264–272. [Google Scholar] [CrossRef]
- Monti, J.M. Serotonin control of sleep-wake behavior. Sleep Med. Rev. 2011, 15, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Garau, C.; Aparicio, S.; Rial, R.V.; Nicolau, M.C.; Esteban, S. Age related changes in the activity-rest circadian rhythms and c-fos expression of ring doves with aging. Effects of tryptophan intake. Exp. Gerontol. 2006, 41, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.D.; Terron, M.P.; Cubero, J.; Valero, V.; Barriga, C.; Reiter, R.J.; Rodriguez, A.B. Comparative study of the activity/rest rhythms in young and old ringdove (Streptopelia risoria): Correlation with serum levels of melatonin and serotonin. Chronobiol. Int. 2006, 23, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Lin, F.; Zheng, X.; Sehgal, A. Serotonin modulates circadian entrainment in Drosophila. Neuron 2005, 47, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.D. Drosophila melanogaster neurobiology, neuropharmacology, and how the fly can inform central nervous system drug discovery. Pharmacol. Ther. 2006, 112, 677–700. [Google Scholar] [CrossRef]
- Artiushin, G.; Sehgal, A. The Drosophila circuitry of sleep-wake regulation. Curr. Opin. Neurobiol. 2017, 44, 243–250. [Google Scholar] [CrossRef]
- Yuan, Q.; Joiner, W.J.; Sehgal, A. A sleep-promoting role for the Drosophila serotonin receptor 1A. Curr. Biol. CB 2006, 16, 1051–1062. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.; Dircksen, H. Circadian clocks in crustaceans: Identified neuronal and cellular systems. Front. Biosci. Landmark Ed. 2010, 15, 1040–1074. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Chimal, E.G.; Van Herp, F.; Fanjul-Moles, M.L. Daily variations in crustacean hyperglycaemic hormone and serotonin immunoreactivity during the development of crayfish. J. Exp. Biol. 2001, 204, 1073–1081. [Google Scholar]
- Calderon-Rosete, G.; Flores, G.; Rodriguez-Sosa, L. Diurnal rhythm in the levels of the serotonin 5-HT1A receptors in the crayfish eyestalk. Synapse 2006, 59, 368–373. [Google Scholar] [CrossRef]
- Voigt, J.P.; Fink, H. Serotonin controlling feeding and satiety. Behav. Brain Res. 2015, 277, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Higgins, G.A.; Fletcher, P.J.; Shanahan, W.R. Lorcaserin: A review of its preclinical and clinical pharmacology and therapeutic potential. Pharmacol. Ther. 2019, 205, 107417. [Google Scholar] [CrossRef] [PubMed]
- Vickers, S.P.; Dourish, C.T. Serotonin receptor ligands and the treatment of obesity. Curr. Opin. Investig. Drugs 2004, 5, 377–388. [Google Scholar] [PubMed]
- D’Agostino, G.; Lyons, D.; Cristiano, C.; Lettieri, M.; Olarte-Sanchez, C.; Burke, L.K.; Greenwald-Yarnell, M.; Cansell, C.; Doslikova, B.; Georgescu, T.; et al. Nucleus of the Solitary Tract Serotonin 5-HT2C Receptors Modulate Food Intake. Cell Metab. 2018, 28, 619–630. [Google Scholar] [CrossRef] [Green Version]
- Valencia-Torres, L.; Olarte-Sánchez, C.M.; Lyons, D.J.; Georgescu, T.; Greenwald-Yarnell, M.; Myers, M.G., Jr.; Bradshaw, C.M.; Heisler, L.K. Activation of Ventral Tegmental Area 5-HT(2C) Receptors Reduces Incentive Motivation. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2017, 42, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Anastasio, N.C.; Stutz, S.J.; Fox, R.G.; Sears, R.M.; Emeson, R.B.; DiLeone, R.J.; O’Neil, R.T.; Fink, L.H.; Li, D.; Green, T.A.; et al. Functional status of the serotonin 5-HT2C receptor (5-HT2CR) drives interlocked phenotypes that precipitate relapse-like behaviors in cocaine dependence. Neuropsychopharmacology 2014, 39, 370–382. [Google Scholar] [CrossRef] [Green Version]
- De Deurwaerdere, P.; Navailles, S.; Berg, K.A.; Clarke, W.P.; Spampinato, U. Constitutive activity of the serotonin2C receptor inhibits in vivo dopamine release in the rat striatum and nucleus accumbens. J. Neurosci. 2004, 24, 3235–3241. [Google Scholar] [CrossRef]
- De Deurwaerdere, P.; Ramos, M.; Bharatiya, R.; Puginier, E.; Chagraoui, A.; Manem, J.; Cuboni, E.; Pierucci, M.; Deidda, G.; Casarrubea, M.; et al. Lorcaserin bidirectionally regulates dopaminergic function site-dependently and disrupts dopamine brain area correlations in rats. Neuropharmacology 2019, 166, 107915. [Google Scholar] [CrossRef]
- Higgins, G.A.; Fletcher, P.J. Therapeutic Potential of 5-HT2C Receptor Agonists for Addictive Disorders. ACS Chem. Neurosci. 2015, 6, 1071–1088. [Google Scholar] [CrossRef]
- Higgins, G.A.; Silenieks, L.B.; Altherr, E.B.; MacMillan, C.; Fletcher, P.J.; Pratt, W.E. Lorcaserin and CP-809101 reduce motor impulsivity and reinstatement of food seeking behavior in male rats: Implications for understanding the anti-obesity property of 5-HT2C receptor agonists. Psychopharmacology 2016, 233, 2841–2856. [Google Scholar] [CrossRef]
- Bockaert, J.; Claeysen, S.; Compan, V.; Dumuis, A. 5-HT(4) receptors, a place in the sun: Act two. Curr. Opin. Pharmacol. 2011, 11, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Jean, A.; Laurent, L.; Bockaert, J.; Charnay, Y.; Dusticier, N.; Nieoullon, A.; Barrot, M.; Neve, R.; Compan, V. The nucleus accumbens 5-HTR(4)-CART pathway ties anorexia to hyperactivity. Transl. Psychiatry 2012, 2, e203. [Google Scholar] [CrossRef] [PubMed]
- Haahr, M.E.; Rasmussen, P.M.; Madsen, K.; Marner, L.; Ratner, C.; Gillings, N.; Baare, W.F.; Knudsen, G.M. Obesity is associated with high serotonin 4 receptor availability in the brain reward circuitry. Neuroimage 2012, 61, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Rebholz, H.; Friedman, E.; Castello, J. Alterations of Expression of the Serotonin 5-HT4 Receptor in Brain Disorders. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luedtke, S.; O’Connor, V.; Holden-Dye, L.; Walker, R.J. The regulation of feeding and metabolism in response to food deprivation in Caenorhabditis elegans. Invertebr. Neurosci. IN 2010, 10, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Iwanir, S.; Kopito, R.B.; Scholz, M.; Calarco, J.A.; Biron, D.; Levine, E. Serotonin-dependent kinetics of feeding bursts underlie a graded response to food availability in C. elegans. Nat. Commun. 2017, 8, 14221. [Google Scholar] [CrossRef] [PubMed]
- Orchard, I. Peptides and serotonin control feeding-related events in Rhodnius prolixus. Front. Biosci. Elite Ed. 2009, 1, 250–262. [Google Scholar]
- Edwards, D.H.; Spitzer, N. Social dominance and serotonin receptor genes in crayfish. Curr. Top. Dev. Biol. 2006, 74, 177–199. [Google Scholar] [CrossRef]
- Stevenson, P.A.; Rillich, J. The decision to fight or flee - insights into underlying mechanism in crickets. Front. Neurosci. 2012, 6, 118. [Google Scholar] [CrossRef] [Green Version]
- Miczek, K.A.; Fish, E.W.; De Bold, J.F.; De Almeida, R.M. Social and neural determinants of aggressive behavior: Pharmacotherapeutic targets at serotonin, dopamine and gamma-aminobutyric acid systems. Psychopharmacology 2002, 163, 434–458. [Google Scholar] [CrossRef]
- Yeh, S.R.; Fricke, R.A.; Edwards, D.H. The effect of social experience on serotonergic modulation of the escape circuit of crayfish. Science 1996, 271, 366–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, S.R.; Musolf, B.E.; Edwards, D.H. Neuronal adaptations to changes in the social dominance status of crayfish. J. Neurosci. 1997, 17, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Cattaert, D.; Delbecque, J.P.; Edwards, D.H.; Issa, F.A. Social interactions determine postural network sensitivity to 5-HT. J. Neurosci. 2010, 30, 5603–5616. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Smith, K.; Delago, A.; Isaksson, K.; Kravitz, E.A. Serotonin and aggressive motivation in crustaceans: Altering the decision to retreat. Proc. Natl. Acad. Sci. USA 1997, 94, 5939–5942. [Google Scholar] [CrossRef] [Green Version]
- Bacque-Cazenave, J.; Cattaert, D.; Delbecque, J.P.; Fossat, P. Alteration of size perception: Serotonin has opposite effects on the aggressiveness of crayfish confronting either a smaller or a larger rival. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Fachinelli, C.; Sargo, S.; Bataller, R.; Rodriguez Echandia, E.L. Effect of 5-HTP and ketanserine on the aggressive reaction induced by food competition in dominant and submissive pigeons (Columba livia). Behav. Brain Res. 1989, 35, 265–270. [Google Scholar] [CrossRef]
- Larson, E.T.; Summers, C.H. Serotonin reverses dominant social status. Behav. Brain Res. 2001, 121, 95–102. [Google Scholar] [CrossRef]
- Murakami, S.; Itoh, M.T. Effects of aggression and wing removal on brain serotonin levels in male crickets, Gryllus bimaculatus. J. Insect Physiol. 2001, 47, 1309–1312. [Google Scholar] [CrossRef]
- Dierick, H.A.; Greenspan, R.J. Serotonin and neuropeptide F have opposite modulatory effects on fly aggression. Nat. Genet. 2007, 39, 678–682. [Google Scholar] [CrossRef]
- Alekseyenko, O.V.; Lee, C.; Kravitz, E.A. Targeted manipulation of serotonergic neurotransmission affects the escalation of aggression in adult male Drosophila melanogaster. PLoS ONE 2010, 5, e10806. [Google Scholar] [CrossRef] [Green Version]
- Alekseyenko, O.V.; Chan, Y.B.; Okaty, B.W.; Chang, Y.; Dymecki, S.M.; Kravitz, E.A. Serotonergic Modulation of Aggression in Drosophila Involves GABAergic and Cholinergic Opposing Pathways. Curr. Biol. 2019, 29, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Bubak, A.N.; Watt, M.J.; Yaeger, J.D.W.; Renner, K.J.; Swallow, J.G. The stalk-eyed fly as a model for aggression - is there a conserved role for 5-HT between vertebrates and invertebrates? J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef]
- Edsinger, E.; Dolen, G. A Conserved Role for Serotonergic Neurotransmission in Mediating Social Behavior in Octopus. Curr. Biol. CB 2018, 28, 3136–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, M.C. The Neurobiology of Mutualistic Behavior: The Cleanerfish Swims into the Spotlight. Front. Behav. Neurosci. 2017, 11, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, M.S.; Messias, J.P.M.; Thornqvist, P.O.; Winberg, S.; Soares, M.C. Monoaminergic levels at the forebrain and diencephalon signal for the occurrence of mutualistic and conspecific engagement in client reef fish. Sci. Rep. 2018, 8, 7346. [Google Scholar] [CrossRef]
- Miczek, K.A.; Maxson, S.C.; Fish, E.W.; Faccidomo, S. Aggressive behavioral phenotypes in mice. Behav. Brain Res. 2001, 125, 167–181. [Google Scholar] [CrossRef]
- Olivier, B.; van Oorschot, R. 5-HT1B receptors and aggression: A review. Eur. J. Pharmacol. 2005, 526, 207–217. [Google Scholar] [CrossRef]
- Howell, S.; Westergaard, G.; Hoos, B.; Chavanne, T.J.; Shoaf, S.E.; Cleveland, A.; Snoy, P.J.; Suomi, S.J.; Dee Higley, J. Serotonergic influences on life-history outcomes in free-ranging male rhesus macaques. Am. J. Primatol. 2007, 69, 851–865. [Google Scholar] [CrossRef]
- Moeller, F.G.; Dougherty, D.M.; Swann, A.C.; Collins, D.; Davis, C.M.; Cherek, D.R. Tryptophan depletion and aggressive responding in healthy males. Psychopharmacology 1996, 126, 97–103. [Google Scholar] [CrossRef]
- Uchida, S.; Kitamoto, A.; Umeeda, H.; Nakagawa, N.; Masushige, S.; Kida, S. Chronic reduction in dietary tryptophan leads to changes in the emotional response to stress in mice. J. Nutr. Sci. Vitaminol. 2005, 51, 175–181. [Google Scholar] [CrossRef]
- De Boer, S.F.; Koolhaas, J.M. 5-HT1A and 5-HT1B receptor agonists and aggression: A pharmacological challenge of the serotonin deficiency hypothesis. Eur. J. Pharmacol. 2005, 526, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Weerts, E.; Haney, M.; Tidey, J. Neurobiological mechanisms controlling aggression: Preclinical developments for pharmacotherapeutic interventions. Neurosci. Biobehav. Rev. 1994, 18, 97–110. [Google Scholar] [CrossRef]
- Saudou, F.; Amara, D.A.; Dierich, A.; LeMeur, M.; Ramboz, S.; Segu, L.; Buhot, M.C.; Hen, R. Enhanced aggressive behavior in mice lacking 5-HT1B receptor. Science 1994, 265, 1875–1878. [Google Scholar] [CrossRef] [PubMed]
- Johnson, O.; Becnel, J.; Nichols, C.D. Serotonin 5-HT(2) and 5-HT(1A)-like receptors differentially modulate aggressive behaviors in Drosophila melanogaster. Neuroscience 2009, 158, 1292–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rillich, J.; Rillich, B.; Stevenson, P.A. Differential modulation of courtship behavior and subsequent aggression by octopamine, dopamine and serotonin in male crickets. Horm. Behav. 2019, 114, 104542. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J. The neurobiology and control of anxious states. Prog. Neurobiol. 2003, 70, 83–244. [Google Scholar] [CrossRef]
- Bacque-Cazenave, J.; Cattaert, D.; Delbecque, J.P.; Fossat, P. Social harassment induces anxiety-like behaviour in crayfish. Sci. Rep. 2017, 7, 39935. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, F.; Aryal, S.; Ho, J.; Stewart, J.C.; Norman, N.A.; Tan, T.L.; Eisaka, A.; Claridge-Chang, A. Ancient Anxiety Pathways Influence Drosophila Defense Behaviors. Curr. Biol. CB 2016, 26, 981–986. [Google Scholar] [CrossRef] [Green Version]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Di Giovanni, G.; De Deurwaerdere, P. New therapeutic opportunities for 5-HT2C receptor ligands in neuropsychiatric disorders. Pharmacol. Ther. 2016, 157, 125–162. [Google Scholar] [CrossRef]
- Millan, M.J. Serotonin 5-HT2C receptors as a target for the treatment of depressive and anxious states: Focus on novel therapeutic strategies. Therapie 2005, 60, 441–460. [Google Scholar] [CrossRef] [PubMed]
- Ries, A.S.; Hermanns, T.; Poeck, B.; Strauss, R. Serotonin modulates a depression-like state in Drosophila responsive to lithium treatment. Nat. Commun. 2017, 8, 15738. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasper, S.; Hamon, M. Beyond the monoaminergic hypothesis: Agomelatine, a new antidepressant with an innovative mechanism of action. World J. Biol. Psychiatry Off. J. World Fed. Soc. Biol. Psychiatry 2009, 10, 117–126. [Google Scholar] [CrossRef]
- Lopez-Munoz, F.; Alamo, C. Monoaminergic neurotransmission: The history of the discovery of antidepressants from 1950s until today. Curr. Pharm. Des. 2009, 15, 1563–1586. [Google Scholar] [CrossRef] [Green Version]
- De Deurwaerdere, P.; Di Giovanni, G. Serotonergic modulation of the activity of mesencephalic dopaminergic systems: Therapeutic implications. Prog. Neurobiol. 2017, 151, 175–236. [Google Scholar] [CrossRef]
- Artigas, F.; Nutt, D.J.; Shelton, R. Mechanism of action of antidepressants. Psychopharmacol. Bull. 2002, 36 (Suppl. 2), 123–132. [Google Scholar]
- Blenau, W.; Thamm, M. Distribution of serotonin (5-HT) and its receptors in the insect brain with focus on the mushroom bodies: Lessons from Drosophila melanogaster and Apis mellifera. Arthropod Struct. Dev. 2011, 40, 381–394. [Google Scholar] [CrossRef]
- Pittenger, C.; Kandel, E.R. In search of general mechanisms for long-lasting plasticity: Aplysia and the hippocampus. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Buhot, M.C. Serotonin receptors in cognitive behaviors. Curr. Opin. Neurobiol. 1997, 7, 243–254. [Google Scholar] [CrossRef]
- Svob Strac, D.; Pivac, N.; Muck-Seler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, I.E.M.; Mork, A. Antagonism of the 5-HT6 receptor—Preclinical rationale for the treatment of Alzheimer’s disease. Neuropharmacology 2017, 125, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Khoury, R.; Grysman, N.; Gold, J.; Patel, K.; Grossberg, G.T. The role of 5 HT6-receptor antagonists in Alzheimer’s disease: An update. Expert Opin. Investig. Drugs 2018, 27, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Meneses, A. Serotonin, neural markers, and memory. Front. Pharmacol. 2015, 6, 143. [Google Scholar] [CrossRef]
- Meneses, A. Could the 5-HT1B receptor inverse agonism affect learning consolidation? Neurosci. Biobehav. Rev. 2001, 25, 193–201. [Google Scholar] [CrossRef]
- Harvey, J.A.; Welsh, S.E.; Hood, H.; Romano, A.G. Effect of 5-HT2 receptor antagonists on a cranial nerve reflex in the rabbit: Evidence for inverse agonism. Psychopharmacology 1999, 141, 162–168. [Google Scholar] [CrossRef]
- De Deurwaerdère, P.; Drutel, G.; Di Giovanni, G. Pharmacological analysis in favour of a physiological role for the constitutive activity of 5-HT2A receptors in learning. In 5-HT2A Receptors in the Central Nervous System; Guiard, B., Di Giovanni, G., Eds.; Humana Press: Passaic, NJ, USA, 2018; pp. 3–29. [Google Scholar]
- Harvey, J.A. Role of the serotonin 5-HT(2A) receptor in learning. Learn. Mem. Cold Spring Harb. N. Y. 2003, 10, 355–362. [Google Scholar] [CrossRef] [Green Version]
- De Deurwaerdere, P.; Bharatiya, R.; Chagraoui, A.; Di Giovanni, G. Constitutive activity of 5-HT receptors: Factual analysis. Neuropharmacology 2020, 17, 107967. [Google Scholar] [CrossRef]
- Dunn, B.D.; Dalgleish, T.; Lawrence, A.D. The somatic marker hypothesis: A critical evaluation. Neurosci. Biobehav. Rev. 2006, 30, 239–271. [Google Scholar] [CrossRef]
- Daruna, J.; Barnes, P. The impulsive client: Theory, research and treatment. In A Neurodevelopmental View of Impulsivityand Its Relationship to the Superfactors of Personality; McCown, W., Johnson, J., Shure, M., Eds.; American Psychological Association: Washington, DC, USA, 1993. [Google Scholar]
- Dalley, J.W.; Everitt, B.J.; Robbins, T.W. Impulsivity, compulsivity, and top-down cognitive control. Neuron 2011, 69, 680–694. [Google Scholar] [CrossRef] [Green Version]
- Dalley, J.W.; Mar, A.C.; Economidou, D.; Robbins, T.W. Neurobehavioral mechanisms of impulsivity: Fronto-striatal systems and functional neurochemistry. Pharmacol. Biochem. Behav. 2008, 90, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Sieling, F.; Bedecarrats, A.; Simmers, J.; Prinz, A.A.; Nargeot, R. Differential roles of nonsynaptic and synaptic plasticity in operant reward learning-induced compulsive behavior. Curr. Biol. CB 2014, 24, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, T.G.; Persaud, N. The adaptiveness of self-control: Simulation of foraging mice. J. Comp. Psychol. 2008, 122, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pu, Y.; Shen, P. Neuropeptide-gated perception of appetitive olfactory inputs in Drosophila larvae. Cell Rep. 2013, 3, 820–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellu-Hagedorn, F. Relationship between impulsivity, hyperactivity and working memory: A differential analysis in the rat. Behav. Brain Funct. BBF 2006, 2, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivalan, M.; Valton, V.; Series, P.; Marchand, A.R.; Dellu-Hagedorn, F. Elucidating poor decision-making in a rat gambling task. PLoS ONE 2013, 8, e82052. [Google Scholar] [CrossRef] [PubMed]
- De Haas, E.N.; van der Eijk, J.A.J. Where in the serotonergic system does it go wrong? Unravelling the route by which the serotonergic system affects feather pecking in chickens. Neurosci. Biobehav. Rev. 2018, 95, 170–188. [Google Scholar] [CrossRef]
- Grant, J.E.; Kim, S.W. Brain circuitry of compulsivity and impulsivity. CNS Spectr. 2014, 19, 21–27. [Google Scholar] [CrossRef]
- Clarke, H.F.; Dalley, J.W.; Crofts, H.S.; Robbins, T.W.; Roberts, A.C. Cognitive inflexibility after prefrontal serotonin depletion. Science 2004, 304, 878–880. [Google Scholar] [CrossRef] [Green Version]
- Clarke, H.F.; Walker, S.C.; Dalley, J.W.; Robbins, T.W.; Roberts, A.C. Cognitive inflexibility after prefrontal serotonin depletion is behaviorally and neurochemically specific. Cereb. Cortex 2007, 17, 18–27. [Google Scholar] [CrossRef]
- Flaisher-Grinberg, S.; Klavir, O.; Joel, D. The role of 5-HT2A and 5-HT2C receptors in the signal attenuation rat model of obsessive-compulsive disorder. Int. J. Neuropsychopharmacol. 2008, 11, 811–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsio, J.; Nilsson, S.R.; Gastambide, F.; Wang, R.A.; Dam, S.A.; Mar, A.C.; Tricklebank, M.; Robbins, T.W. The role of 5-HT2C receptors in touchscreen visual reversal learning in the rat: A cross-site study. Psychopharmacology 2015, 232, 4017–4031. [Google Scholar] [CrossRef] [PubMed]
- Kreiss, D.S.; De Deurwaerdere, P. Purposeless oral activity induced by meta-chlorophenylpiperazine (m-CPP): Undefined tic-like behaviors? J. Neurosci. Methods 2017, 292, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S.V.; Gutierrez-Ferre, V.; Flores, P.; Moreno, M. Activation of serotonin 5-HT2A receptors inhibits high compulsive drinking on schedule-induced polydipsia. Psychopharmacology 2015, 232, 683–697. [Google Scholar] [CrossRef]
- Anastasio, N.C.; Stutz, S.J.; Fink, L.H.; Swinford-Jackson, S.E.; Sears, R.M.; DiLeone, R.J.; Rice, K.C.; Moeller, F.G.; Cunningham, K.A. Serotonin (5-HT) 5-HT2A Receptor (5-HT2AR):5-HT2CR Imbalance in Medial Prefrontal Cortex Associates with Motor Impulsivity. ACS Chem. Neurosci. 2015, 6, 1248–1258. [Google Scholar] [CrossRef] [Green Version]
- Winstanley, C.A.; Theobald, D.E.; Dalley, J.W.; Robbins, T.W. Interactions between serotonin and dopamine in the control of impulsive choice in rats: Therapeutic implications for impulse control disorders. Neuropsychopharmacology 2005, 30, 669–682. [Google Scholar] [CrossRef]
- Dellu-Hagedorn, F.; Rivalan, M.; Fitoussi, A.; De Deurwaerdere, P. Inter-individual differences in the impulsive/compulsive dimension: Deciphering related dopaminergic and serotonergic metabolisms at rest. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- De Gregorio, D.; McLaughlin, R.J.; Posa, L.; Ochoa-Sanchez, R.; Enns, J.; Lopez-Canul, M.; Aboud, M.; Maione, S.; Comai, S.; Gobbi, G. Cannabidiol modulates serotonergic transmission and reverses both allodynia and anxiety-like behavior in a model of neuropathic pain. Pain 2019, 160, 136–150. [Google Scholar] [CrossRef]
- Millan, M.J. The induction of pain: An integrative review. Prog. Neurobiol. 1999, 57, 1–164. [Google Scholar] [CrossRef]
- Pineda-Farias, J.B.; Barragan-Iglesias, P.; Valdivieso-Sanchez, A.; Rodriguez-Silverio, J.; Flores-Murrieta, F.J.; Granados-Soto, V.; Rocha-Gonzalez, H.I. Spinal 5-HT4 and 5-HT6 receptors contribute to the maintenance of neuropathic pain in rats. Pharmacol. Rep. PR 2017, 69, 916–923. [Google Scholar] [CrossRef]
- Perrin, F.E.; Noristani, H.N. Serotonergic mechanisms in spinal cord injury. Exp. Neurol. 2019, 318, 174–191. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacqué-Cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J.-P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdère, P. Serotonin in Animal Cognition and Behavior. Int. J. Mol. Sci. 2020, 21, 1649. https://doi.org/10.3390/ijms21051649
Bacqué-Cazenave J, Bharatiya R, Barrière G, Delbecque J-P, Bouguiyoud N, Di Giovanni G, Cattaert D, De Deurwaerdère P. Serotonin in Animal Cognition and Behavior. International Journal of Molecular Sciences. 2020; 21(5):1649. https://doi.org/10.3390/ijms21051649
Chicago/Turabian StyleBacqué-Cazenave, Julien, Rahul Bharatiya, Grégory Barrière, Jean-Paul Delbecque, Nouhaila Bouguiyoud, Giuseppe Di Giovanni, Daniel Cattaert, and Philippe De Deurwaerdère. 2020. "Serotonin in Animal Cognition and Behavior" International Journal of Molecular Sciences 21, no. 5: 1649. https://doi.org/10.3390/ijms21051649
APA StyleBacqué-Cazenave, J., Bharatiya, R., Barrière, G., Delbecque, J. -P., Bouguiyoud, N., Di Giovanni, G., Cattaert, D., & De Deurwaerdère, P. (2020). Serotonin in Animal Cognition and Behavior. International Journal of Molecular Sciences, 21(5), 1649. https://doi.org/10.3390/ijms21051649