Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species

Abstract
:1. Introduction
2. Results
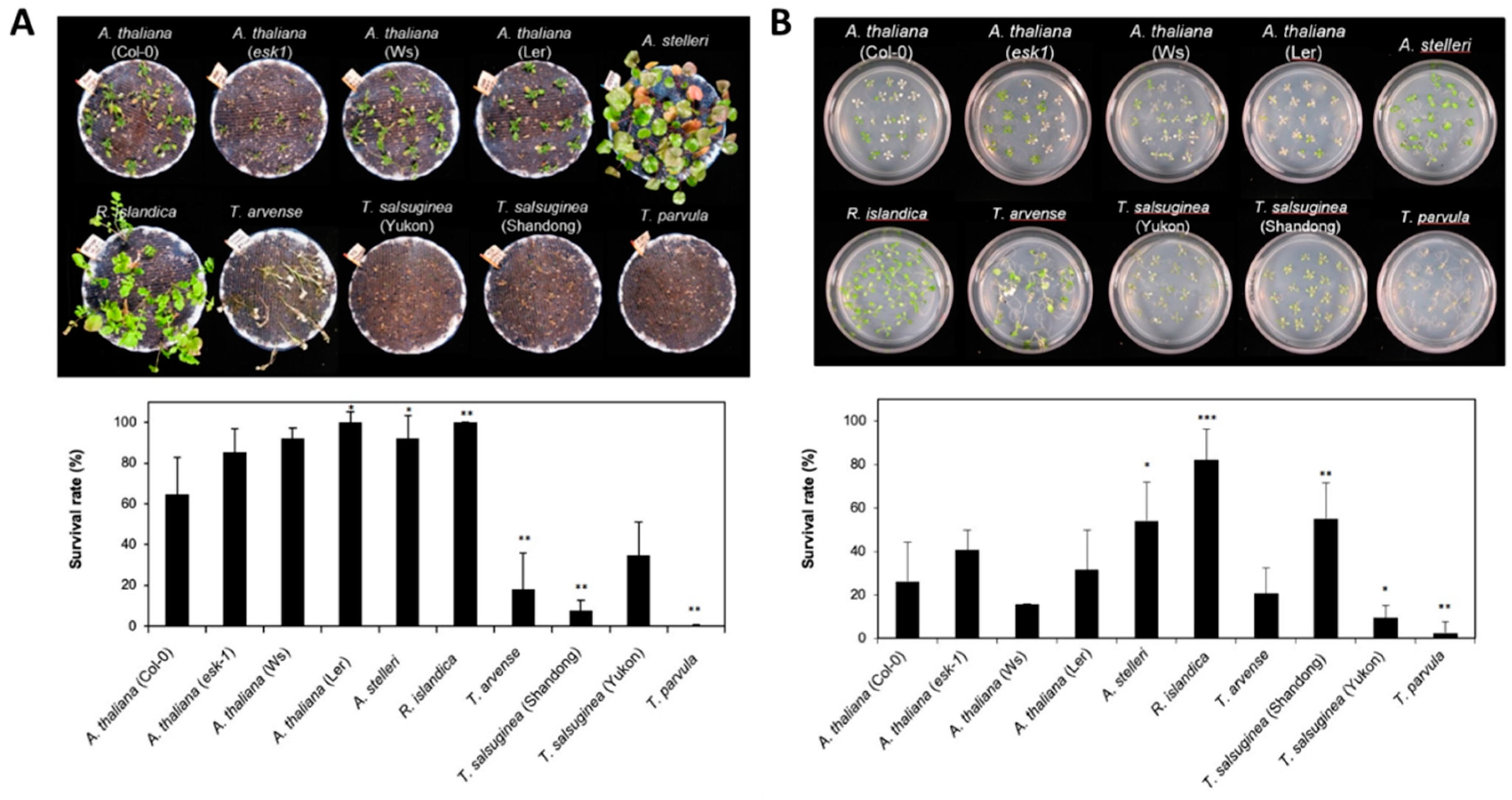
2.1. Physiological Response to Low-Oxygen Stress in Species Closely Related to Arabidopsis
2.2. Expression Profile Analysis of the Low-Oxygen Response in A. thaliana, A. stelleri, R. islandica, and T. salsuginea
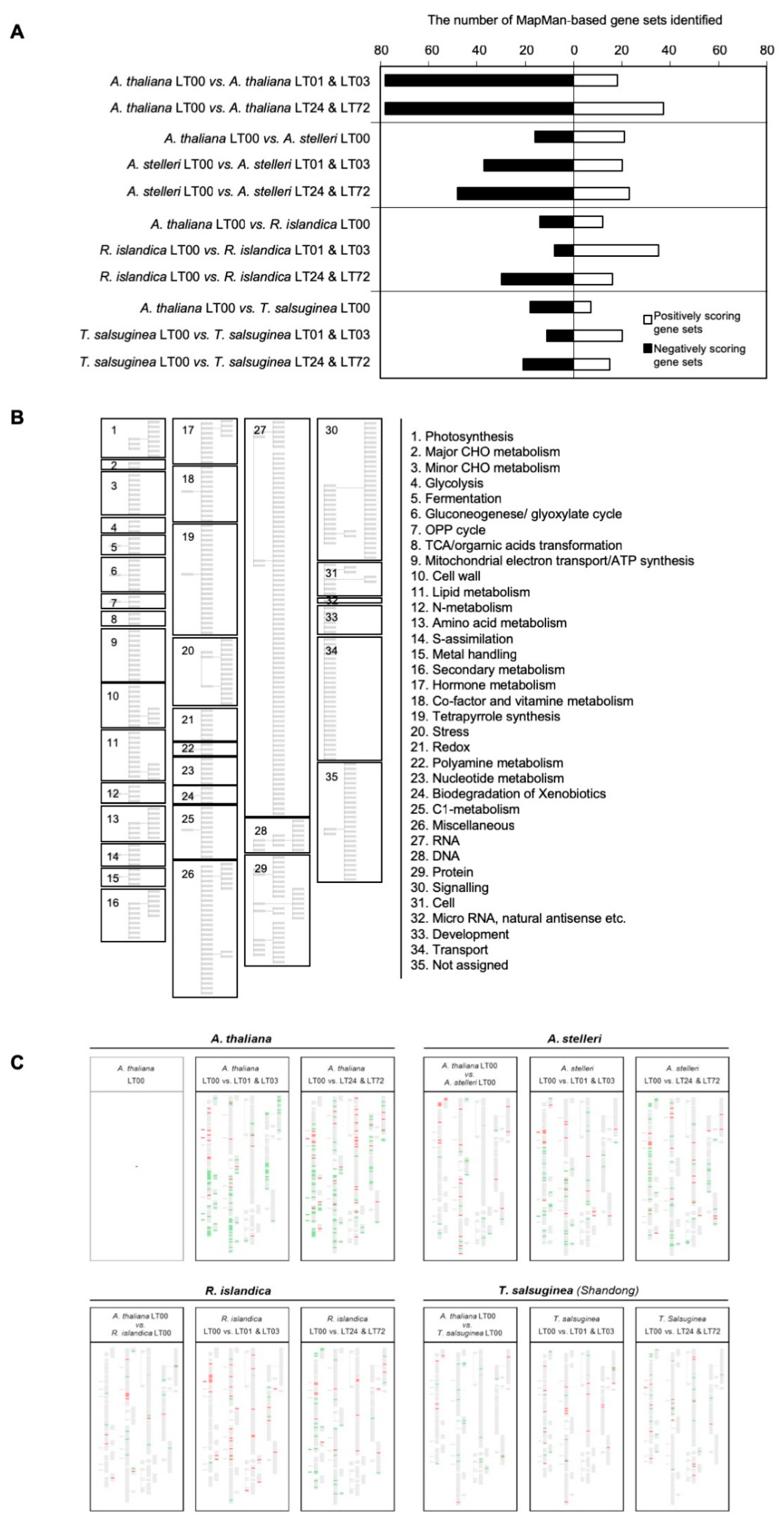
2.3. Pathway Profiles Based on Gene-Set Enrichment Analysis
2.3.1. A. thaliana: Moderately Tolerant Species
2.3.2. A. stelleri: Highly Tolerant Species
2.3.3. R. islandica: Highly Tolerant Species
2.3.4. T. salsuginea Shangdong: Low-Oxygen Sensitive Species
2.4. Interspecies Variation in Reconfiguration of Energy Metabolism under Low-Oxygen Stress
2.5. Confirmation of Changes in Root Temperatures under Low-Oxygen Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth
4.2. Submergence and Low-Oxygen Treatment
4.3. RNA Isolation and Purification
4.4. Microarray Experiment, Image Acquisition, Data Acquisition and Normalization
4.5. GSEA Analysis Using MapMan-Based Gene Set Database
4.6. Infrared Thermography
4.7. ITS PCR Amplification and Analysis
4.8. Semi-Quantitative RT-PCR
4.9. Measurement of Photosynthetic Activity in Root
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Subbaiah, C.C.; Sachs, M.M. Molecular and cellular adaptations of maize to flooding stress. Ann. Bot. 2003, 91, 119–127. [Google Scholar] [CrossRef]
- Liu, F.; Vantoai, T.; Moy, L.P.; Bock, G.; Linford, L.D.; Quackenbush, J. Global transcription profiling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol. 2005, 137, 1115–1129. [Google Scholar] [CrossRef] [Green Version]
- Sachs, M.M.; Freeling, M.; Okimoto, R. The anaerobic proteins of maize. Cell 1980, 20, 761–767. [Google Scholar] [CrossRef]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic Metabolism in Plants. Plant Physiol. 1992, 100, 1. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Hill, R.D. Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J. Exp. Bot. 2004, 55, 2473–2482. [Google Scholar] [CrossRef] [Green Version]
- Loreti, E.; Poggi, A.; Novi, G.; Alpi, A.; Perata, P. A Genome-Wide Analysis of the Effects of Sucrose on Gene Expression in Arabidopsis Seedlings under Anoxia. Plant Physiol. 2005, 137, 1130. [Google Scholar] [CrossRef] [Green Version]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding Stress: Acclimations and Genetic Diversity. Ann. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Klok, E.J.; Wilson, I.W.; Wilson, D.; Chapman, S.C.; Ewing, R.M.; Somerville, S.C.; Peacock, W.J.; Dolferus, R.; Dennis, E.S. Expression profile analysis of the low-oxygen response in Arabidopsis root cultures. Plant Cell 2002, 14, 2481–2494. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.M.; Vriezen, W.H.; van der Straeten, D. Molecular and Physiological Mechanisms of Flooding Avoidance and Tolerance in Rice. Russ. J. Plant Physiol. 2003, 50, 743–751. [Google Scholar] [CrossRef]
- Branco-Price, C.; Kawaguchi, R.; Ferreira, R.B.; Bailey-Serres, J. Genome-wide analysis of transcript abundance and translation in Arabidopsis seedlings subjected to oxygen deprivation. Ann. Bot. 2005, 96, 647–660. [Google Scholar] [CrossRef] [Green Version]
- Lasanthi-Kudahettige, R.; Magneschi, L.; Loreti, E.; Gonzali, S.; Licausi, F.; Novi, G.; Beretta, O.; Vitulli, F.; Alpi, A.; Perata, P. Transcript Profiling of the Anoxic Rice Coleoptile. Plant Physiol. 2007, 144, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.H.; Lee, M.O.; Choy, Y.-H.; Ha-Lee, Y.-M.; Hong, C.B.; Lee, D.-H. Expression Profile Analysis of Hypoxia Responses in Arabidopsis Roots and Shoots. J. Plant Biol. 2011, 54, 373. [Google Scholar] [CrossRef]
- Sasidharan, R.; Mustroph, A. Plant Oxygen Sensing Is Mediated by the N-End Rule Pathway: A Milestone in Plant Anaerobiosis. Plant Cell 2011, 23, 4173. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Mustroph, A.; Sasidharan, R.; Vashisht, D.; Pedersen, O.; Oosumi, T.; Voesenek, L.A.C.J.; Bailey-Serres, J. Molecular characterization of the submergence response of the Arabidopsis thaliana ecotype Columbia. New Phytol. 2011, 190, 457–471. [Google Scholar] [CrossRef]
- Sasidharan, R.; Mustroph, A.; Boonman, A.; Akman, M.; Ammerlaan, A.M.H.; Breit, T.; Schranz, M.E.; Voesenek, L.A.C.J.; van Tienderen, P.H. Root Transcript Profiling of Two Rorippa Species Reveals Gene Clusters Associated with Extreme Submergence Tolerance. Plant Physiol. 2013, 163, 1277. [Google Scholar] [CrossRef] [Green Version]
- Van Veen, H.; Akman, M.; Jamar, D.C.L.; Vreugdenhil, D.; Kooiker, M.; van Tienderen, P.; Voesenek, L.A.C.J.; Schranz, M.E.; Sasidharan, R. Group VII Ethylene Response Factor diversification and regulation in four species from flood-prone environments. Plant Cell Environ. 2014, 37, 2421–2432. [Google Scholar] [CrossRef]
- Jung, Y.; Park, J.; Choi, Y.; Yang, J.-G.; Kim, D.; Kim, B.-G.; Roh, K.; Lee, D.-H.; Auh, C.-K.; Lee, S. Expression Analysis of Proline Metabolism-related Genes From Halophyte Arabis stelleri under Osmotic Stress Conditions. J. Integr. Plant Biol. 2010, 52, 891–903. [Google Scholar] [CrossRef]
- Sharma, N.; Cram, D.; Huebert, T.; Zhou, N.; Parkin, I.A.P. Exploiting the wild crucifer Thlaspi arvense to identify conserved and novel genes expressed during a plant’s response to cold stress. Plant Mol. Biol. 2007, 63, 171–184. [Google Scholar] [CrossRef]
- Inan, G.; Zhang, Q.; Li, P.; Wang, Z.; Cao, Z.; Zhang, H.; Zhang, C.; Quist, T.M.; Goodwin, S.M.; Zhu, J.; et al. Salt cress. A halophyte and cryophyte Arabidopsis relative model system and its applicability to molecular genetic analyses of growth and development of extremophiles. Plant Physiol. 2004, 135, 1718–1737. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.E.; Li, Y.; Whitty, B.R.; Díaz-Camino, C.; Akhter, S.R.; Brandle, J.E.; Golding, G.B.; Weretilnyk, E.A.; Moffatt, B.A.; Griffith, M. Expressed sequence tags from the Yukon ecotype of Thellungiellareveal that gene expression in response to cold, drought and salinity shows little overlap. Plant Mol. Biol. 2005, 58, 561–574. [Google Scholar] [CrossRef]
- Wong, C.E.; Li, Y.; Labbe, A.; Guevara, D.; Nuin, P.; Whitty, B.; Diaz, C.; Golding, G.B.; Gray, G.R.; Weretilnyk, E.A.; et al. Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Arabidopsis. Plant Physiol. 2006, 140, 1437–1450. [Google Scholar] [CrossRef] [Green Version]
- Griffith, M.; Timonin, M.; Wong, A.C.E.; Gray, G.R.; Akhter, S.R.; Saldanha, M.; Rogers, M.A.; Weretilnyk, E.A.; Moffatt, B. Thellungiella: An Arabidopsis-related model plant adapted to cold temperatures. Plant Cell Environ. 2007, 30, 529–538. [Google Scholar] [CrossRef]
- Orsini, F.; D’Urzo, M.P.; Inan, G.; Serra, S.; Oh, D.-H.; Mickelbart, M.V.; Consiglio, F.; Li, X.; Jeong, J.C.; Yun, D.-J.; et al. A comparative study of salt tolerance parameters in 11 wild relatives of Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 3787–3798. [Google Scholar] [CrossRef]
- Feldman, L.J. Regulation of Root Development. Ann. Rev. Plant Physiol. 1984, 35, 223–242. [Google Scholar] [CrossRef]
- Tomsovic, M. “Binge” and continuous drinkers: Characteristics and treatment follow-up. Q. J. Stud. Alcohol 1974, 35, 558–564. [Google Scholar]
- Warwick, S.I.; Francis, A.; Susko, D.J. The biology of Canadian weeds. 9. Thlaspi arvense L. Can. J. Plant Sci. 2002, 82, 803–823. [Google Scholar] [CrossRef] [Green Version]
- Bressan, R.A.; Zhang, C.; Zhang, H.; Hasegawa, P.M.; Bohnert, H.J.; Zhu, J.K. Learning from the Arabidopsis experience. The next gene search paradigm. Plant Physiol. 2001, 127, 1354–1360. [Google Scholar] [CrossRef]
- Warwick, S.I.; Francis, A.; Susko, D.J. Brassicaceae of Canada; Contribution No. 981317.1225; Agriculture and Agri-Food Canada, Eastern Cereal and Oilseed Research Centre: Ottawa, ON, Canada, 2004. [Google Scholar]
- Oh, D.-H.; Dassanayake, M.; Haas, J.S.; Kropornika, A.; Wright, C.; d’Urzo, M.P.; Hong, H.; Ali, S.; Hernandez, A.; Lambert, G.M.; et al. Genome structures and halophyte-specific gene expression of the extremophile Thellungiella parvula in comparison with Thellungiella salsuginea (Thellungiella halophila) and Arabidopsis. Plant Physiol. 2010, 154, 1040–1052. [Google Scholar] [CrossRef] [Green Version]
- Usami, T.; Mochizuki, N.; Kondo, M.; Nishimura, M.; Nagatani, A. Cryptochromes and Phytochromes Synergistically Regulate Arabidopsis Root Greening under Blue Light. Plant Cell Physiol. 2004, 45, 1798–1808. [Google Scholar] [CrossRef]
- Greenway, H.; Gibbs, J. Review: Mechanisms of anoxia tolerance in plants. II. Energy requirements for maintenance and energy distribution to essential processes. Funct. Plant Biol. 2003, 30, 999–1036. [Google Scholar] [CrossRef]
- Williams, L.A. Heat Release in Alcoholic Fermentation: A Critical Reappraisal. Am. J. Enol. Viticult. 1982, 33, 149. [Google Scholar]
- Borevitz, J.O.; Ecker, J.R. Plant Genomics: The Third Wave. Ann. Rev. Genom. Human Gen. 2004, 5, 443–477. [Google Scholar] [CrossRef] [Green Version]
- Irish, V.F.; Benfey, P.N. Beyond Arabidopsis. Translational Biology Meets Evolutionary Developmental Biology. Plant Physiol. 2004, 135, 611. [Google Scholar] [CrossRef] [Green Version]
- Paterson, A.H.; Bowers, J.E.; Chapman, B.A.; Peterson, D.G.; Rong, J.; Wicker, T.M. Comparative genome analysis of monocots and dicots, toward characterization of angiosperm diversity. Curr. Opin. Biotechnol. 2004, 15, 120–125. [Google Scholar] [CrossRef]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.-K.; Shinozaki, K. Comparative Genomics in Salt Tolerance between Arabidopsis and Arabidopsis-Related Halophyte Salt Cress Using Arabidopsis Microarray. Plant Physiol. 2004, 135, 1697. [Google Scholar] [CrossRef] [Green Version]
- Girke, T.; Todd, J.; Ruuska, S.; White, J.; Benning, C.; Ohlrogge, J. Microarray Analysis of Developing Arabidopsis Seeds. Plant Physiol. 2000, 124, 1570. [Google Scholar] [CrossRef] [Green Version]
- Gong, Q.; Li, P.; Ma, S.; Indu Rupassara, S.; Bohnert, H.J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 2005, 44, 826–839. [Google Scholar] [CrossRef]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545. [Google Scholar] [CrossRef] [Green Version]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. Mapman: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Daugherty, C.J.; Musgrave, M.E. Characterization of populations of rapid-cycling Brassica rapa L. selected for differential waterlogging tolerance. J. Exp. Bot. 1994, 45, 385–392. [Google Scholar] [CrossRef]
- Zou, X.-L.; Zeng, L.; Lu, G.-Y.; Cheng, Y.; Xu, J.-S.; Zhang, X.-K. Comparison of transcriptomes undergoing waterlogging at the seedling stage between tolerant and sensitive varieties of Brassica napus L. J. Integr. Agric. 2015, 14, 1723–1734. [Google Scholar] [CrossRef]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and Acclimation Under Hypoxia and Anoxia. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Hill, R.D. Plant mitochondrial function during anaerobiosis. Ann. Bot. 2009, 103, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Munir, E.; Yoon, J.J.; Tokimatsu, T.; Hattori, T.; Shimada, M. A physiological role for oxalic acid biosynthesis in the wood-rotting basidiomycete Fomitopsis palustris. Proc. Natl. Acad. Sci. USA 2001, 98, 11126–11130. [Google Scholar] [CrossRef] [Green Version]
- Berney, M.; Cook, G.M. Unique flexibility in energy metabolism allows mycobacteria to combat starvation and hypoxia. PLoS ONE 2010, 5, e8614. [Google Scholar] [CrossRef] [Green Version]
- Mommer, L.; Visser, E.J.W. Underwater photosynthesis in flooded terrestrial plants: A matter of leaf plasticity. Ann. Bot. 2005, 96, 581–589. [Google Scholar] [CrossRef]
- Kogawara, S.; Yamanoshita, T.; Norisada, M.; Masumori, M.; Kojima, K. Photosynthesis and photoassimilate transport during root hypoxia in Melaleuca cajuputi, a flood-tolerant species, and in Eucalyptus camaldulensis, a moderately flood-tolerant species. Tree Physiol. 2006, 26, 1413–1423. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/ (accessed on 23 May 2019).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- White; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Physiological Characters Affecting Tolerance | Species with Different Accessions | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. thaliana (Col-0) | A. thaliana (esk-1) | A. thaliana (Ws) | A. thaliana (Ler) | A. stelleri | R. islandica | T. arvense | T. salsuginea (Shandong) | T. salsuginea (Yukon) | T. parvula | ||
| Submergence treatment | Growth | ++ | + | ++ | ++ | ++ | +++ | + | + | + | + |
| Survival | ++ | +++ | +++ | +++ | +++ | +++ | + | + | ++ | + | |
| Low-oxygen treatment | Growth | ++ | ++ | + | + | +++ | +++ | ++ | + | ++ | + |
| Survival | + | ++ | + | + | ++ | +++ | + | ++ | + | + | |
| Tolerance range to low-oxygen stress | Moderate | High | Low | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.-H.; Yu, S.-i.; Lee, B.-h.; Lee, D.-H. Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species. Int. J. Mol. Sci. 2020, 21, 1787. https://doi.org/10.3390/ijms21051787
Hwang J-H, Yu S-i, Lee B-h, Lee D-H. Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species. International Journal of Molecular Sciences. 2020; 21(5):1787. https://doi.org/10.3390/ijms21051787
Chicago/Turabian StyleHwang, Ji-Hye, Si-in Yu, Byeong-ha Lee, and Dong-Hee Lee. 2020. "Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species" International Journal of Molecular Sciences 21, no. 5: 1787. https://doi.org/10.3390/ijms21051787
APA StyleHwang, J. -H., Yu, S. -i., Lee, B. -h., & Lee, D. -H. (2020). Modulation of Energy Metabolism Is Important for Low-Oxygen Stress Adaptation in Brassicaceae Species. International Journal of Molecular Sciences, 21(5), 1787. https://doi.org/10.3390/ijms21051787