Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress


Abstract
:
1. Introduction
2. Results
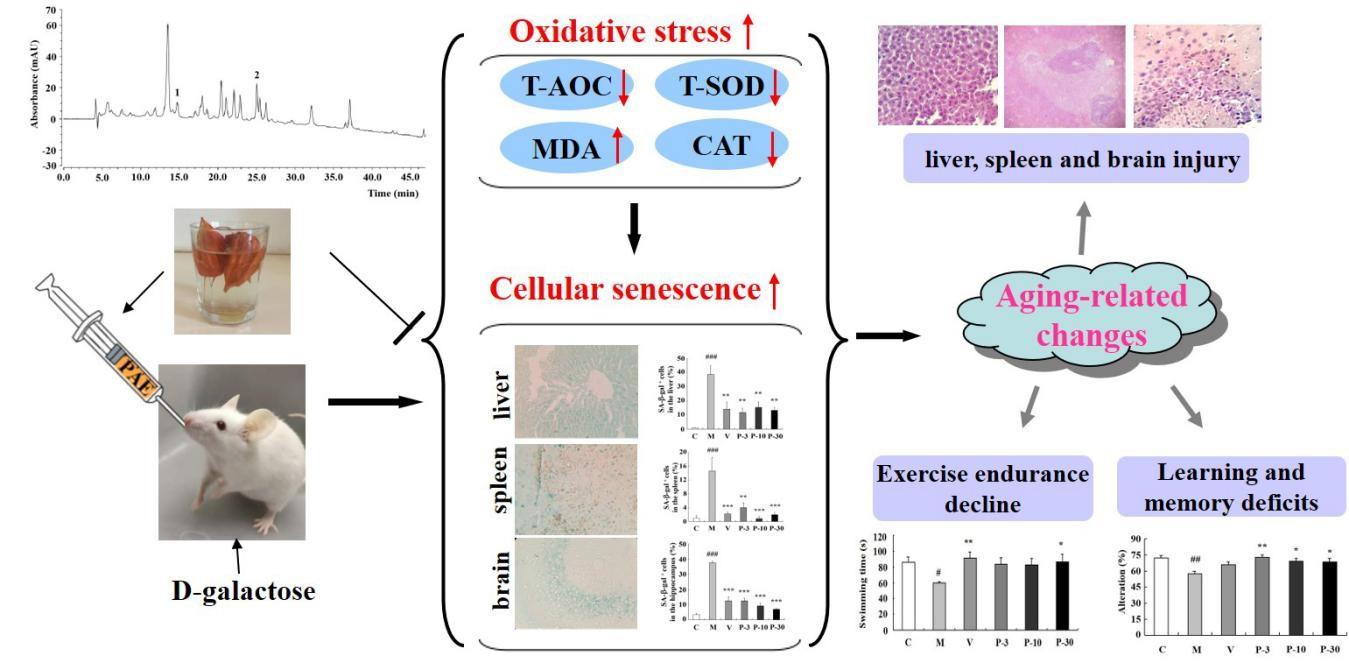
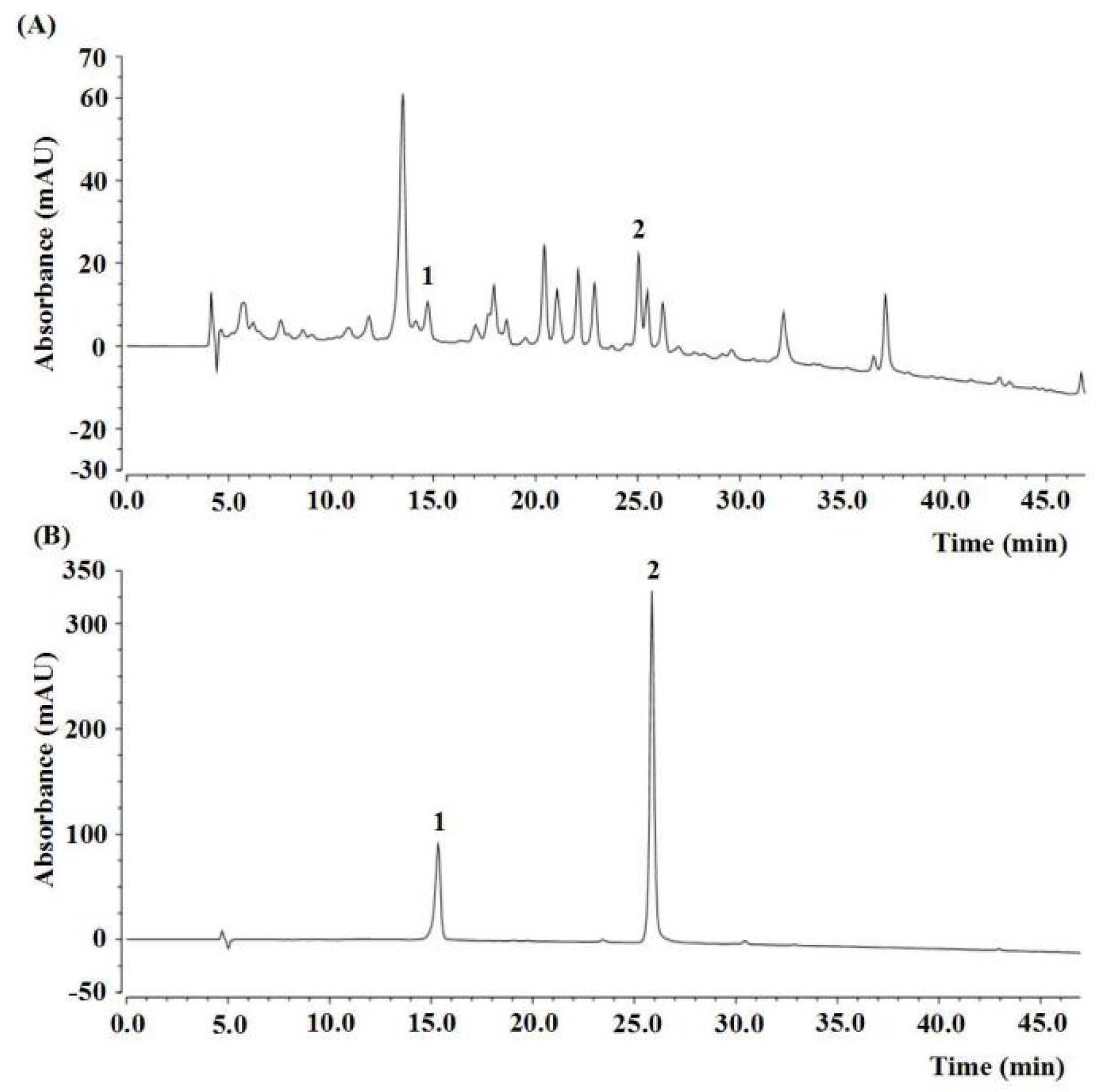
2.1. Component Analysis of PAE
2.2. PAE Improved the d-gal-Induced Recognition Decline
2.3. PAE Ameliorated the d-gal-Induced Working Memory Decline
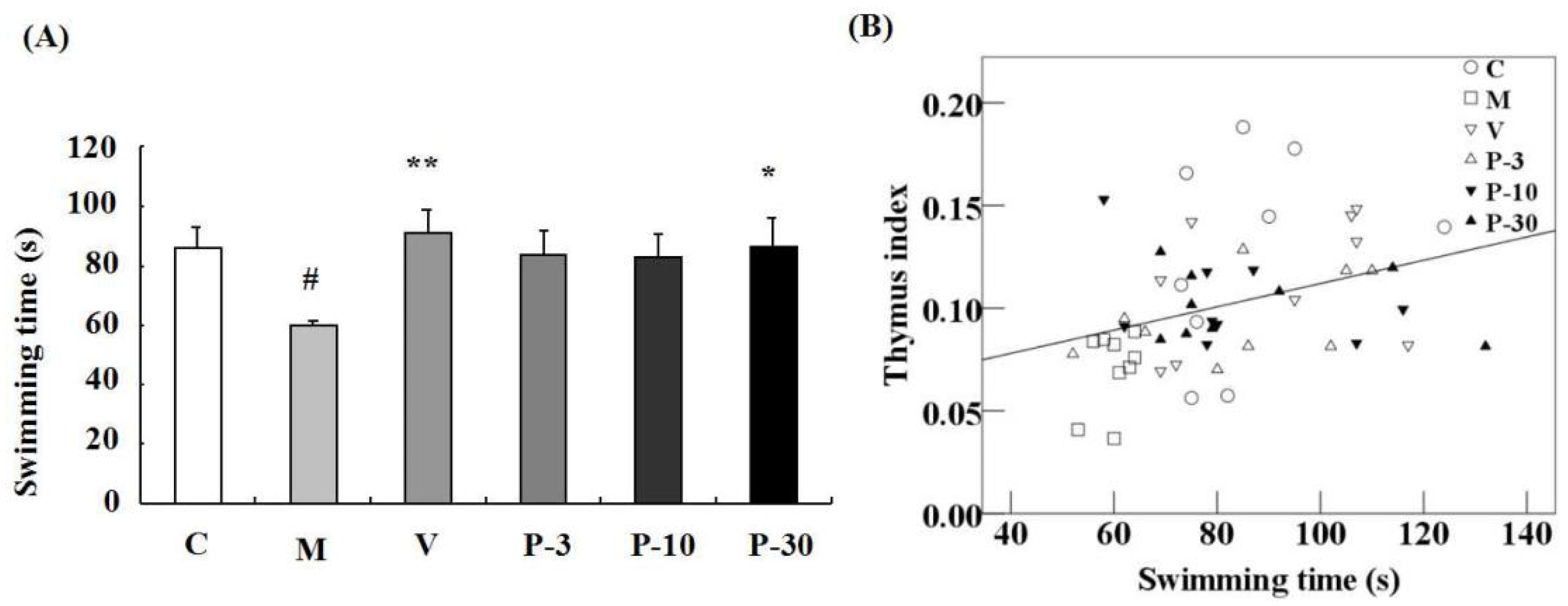
2.4. Effect of PAE on the Weight-Loaded Forced Swimming Test, the Body Weight and Organ Index
2.5. PAE Ameliorated Histopathological Alterations of the Liver, Spleen, and Brain
2.6. Effect of PAE on the Senescence-Associated β-galactosidase Activity of the Liver, Spleen, and Brain
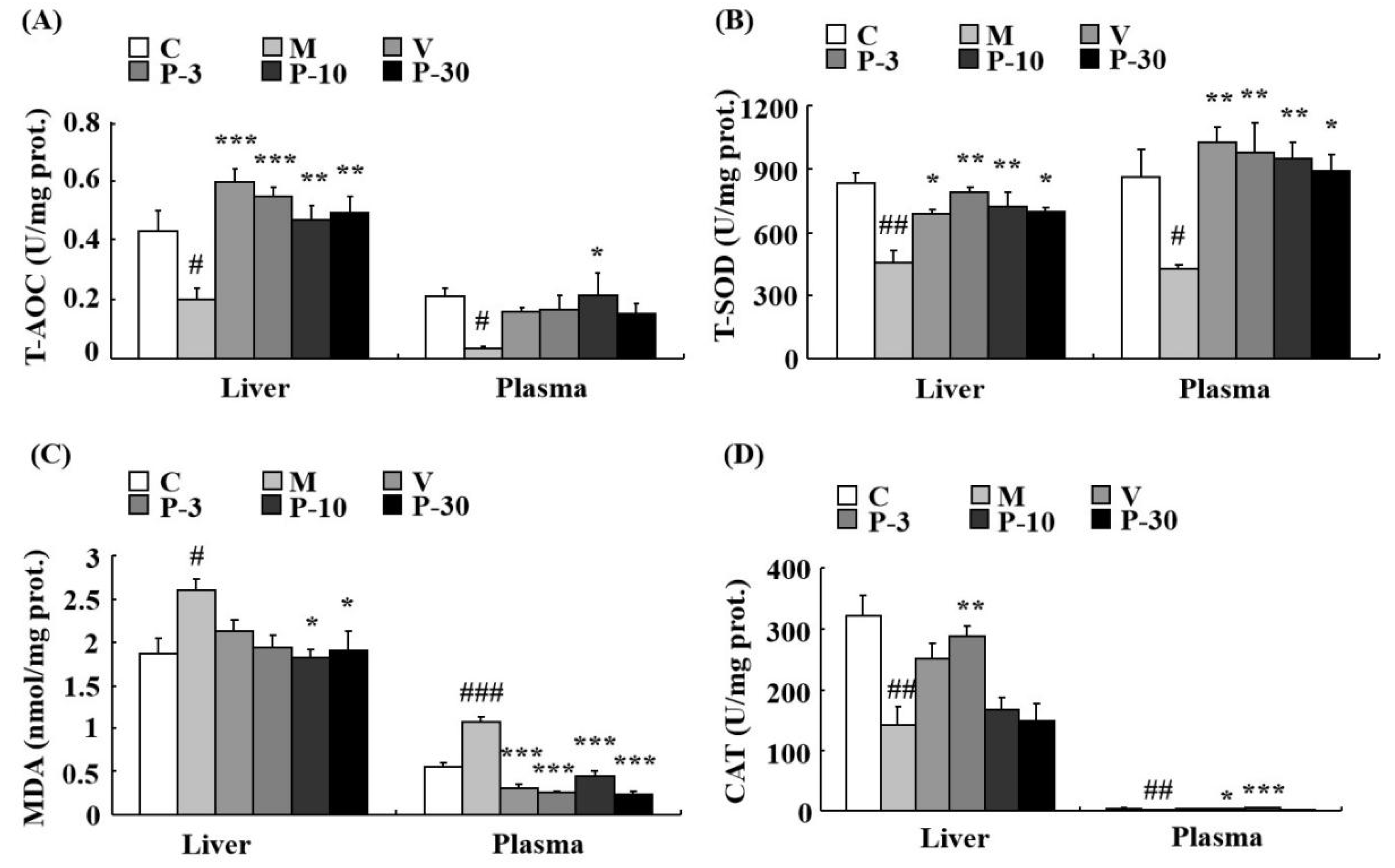
2.7. Anti-Oxidative Stress Effect of PAE on the Liver, Plasma, and Brain of d-gal-Induced Ageing Mice
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Measurement of Total Flavonoid Content
4.3. Determination of Total Phenolic Content
4.4. Determination of Total Saponin Content
4.5. Quantification of Rutin and Luteolin by HPLC
4.6. Chemicals and Kits
4.7. Animals and Drug Administration
4.8. Novel Object Recognition Task
4.9. Spontaneous Alternation Behavior Y-maze Test
4.10. Weight-Loaded Forced Swimming Test
4.11. Bodyweight Measurement and Organ Coefficient Analysis
4.12. Morphological and Histological Analyses
4.13. SA-β-gal Staining
4.14. Biochemical Analysis
4.15. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AChE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| CAT | Catalase |
| d-gal | d-galactose |
| GA | Gallic acid |
| GAEs | Gallic acid equivalents |
| HE | Haematoxylin and eosin |
| HPLC | High-performance liquid chromatography |
| MDA | Malondialdehyde |
| NOR | Novel object recognition |
| OA | Oleanolic acid |
| OAEs | Oleanolic acid equivalents |
| PAE | Ethyl acetate fraction from Physalis Alkekengi |
| REs | Rutin equivalents |
| ROS | Reactive oxygen species |
| SA-β-gal | Senescence-associated β-galactosidase |
| T-AOC | Total anti-oxidant capacity |
| T-SOD | Total superoxide dismutase |
| TFC | Total phenolic content |
| TPC | Total phenolic content |
| TSC | Total saponin content |
| VE | Vitamin E |
| WLFST | Weight-loaded forced swimming test |
References
- Lutz, W.; Sanderson, W.; Scherbov, S. The coming acceleration of global population ageing. Nature 2008, 451, 716–719. [Google Scholar] [CrossRef]
- Hoffman, J.D.; Parikh, I.; Green, S.J.; Chlipala, G.; Mohney, R.P.; Keaton, M.; Bauer, B.; Hartz, A.M.S.; Lin, A.L. Age Drives Distortion of Brain Metabolic, Vascular and Cognitive Functions, and the Gut Microbiome. Front. Aging Neurosci. 2017, 9, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, H.; Hale, L.; Friedberg, F. Prevalence and predictors of fatigue among middle-aged and older adults: Evidence from the health and retirement study. J. Am. Geriatr. Soc. 2010, 58, 2033–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.H.; Sharpless, N.E. Senescence in Health and Disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.G.A.; Faragher, R.G.A. Obesity and type-2 diabetes as inducers of premature cellular senescence and ageing. Biogerontology 2018, 19, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Shakeri, H.; Lemmens, K.; Gevaert, A.B.; De Meyer, G.R.Y.; Segers, V.F.M. Cellular senescence links aging and diabetes in cardiovascular disease. Am. J. Physiol Heart Circ. Physiol. 2018, 315, H448–H462. [Google Scholar] [CrossRef]
- Bellantuono, I. Find drugs that delay many diseases of old age. Nature 2018, 554, 293–295. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics improve physical function and increase lifespan in old age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M.; Miwa, S.; Tchkonia, T.; Tiniakos, D.; Wilson, C.L.; Lahat, A.; Day, C.P.; Burt, A.; Palmer, A.; Anstee, Q.M.; et al. Cellular senescence drives age-dependent hepatic steatosis. Nat. Commun. 2017, 8, 15691. [Google Scholar] [CrossRef]
- Jeon, O.H.; Kim, C.; Laberge, R.M.; Demaria, M.; Rathod, S.; Vasserot, A.P.; Chung, J.W.; Kim, D.H.; Poon, Y.; David, N.; et al. Local clearance of senescent cells attenuates the development of post-traumatic osteoarthritis and creates a pro-regenerative environment. Nat. Med. 2017, 23, 775–781. [Google Scholar] [CrossRef]
- Chinta, S.J.; Woods, G.; Demaria, M.; Rane, A.; Zou, Y.; Mc Quade, A.; Rajagopalan, S.; Limbad, C.; Madden, D.T.; Campisi, J.; et al. Cellular senescence is induced by the environmental neurotoxin paraquat and contributes to neuropathy linked to Parkinson’s disease. Cell Rep. 2018, 22, 930–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussian, T.J.; Aziz, A.; Meyer, C.F.; Swenson, B.L.; van Deursen, J.M.; Baker, D.J. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature 2018, 562, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Gluscevic, M.; Baker, D.J.; Laberge, R.M.; Marquess, D.; Dananberg, J.; van Deursen, J.M. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov. 2017, 16, 718–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueira, I.; Fernandes, A.; Mladenovic Djordjevic, A.; Lopez-Contreras, A.; Henriques, C.M.; Selman, C.; Ferreiro, E.; Gonos, E.S.; Trejo, J.L.; Misra, J.; et al. Interventions for age-related diseases: Shifting the paradigm. Mech. Ageing Dev. 2016, 160, 69–92. [Google Scholar] [CrossRef]
- Zhu, Y.; Tchkonia, T.; Pirtskhalava, T.; Gower, A.C.; Ding, H.; Giorgadze, N.; Palmer, A.K.; Ikeno, Y.; Hubbard, G.B.; Lenburg, M.; et al. The Achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell 2015, 14, 644–658. [Google Scholar] [CrossRef]
- Farr, J.N.; Xu, M.; Weivoda, M.M.; Monroe, D.G.; Fraser, D.G.; Onken, J.L.; Negley, B.A.; Sfeir, J.G.; Ogrodnik, M.B.; Hachfeld, C.M.; et al. Targeting cellular senescence prevents age-related bone loss in mice. Nat. Med. 2017, 23, 1072–1079. [Google Scholar] [CrossRef]
- Soefje, S.A.; Karnad, A.; Brenner, A.J. Common toxicities of mammalian target of rapamycin inhibitors. Target Oncol. 2011, 6, 125–129. [Google Scholar] [CrossRef]
- Wang, G.S.; Hoyte, C. Review of Biguanide (Metformin) Toxicity. J. Intensiv. Care Med. 2018. [CrossRef]
- Hong, J.M.; Kwon, O.K.; Shin, I.S.; Song, H.H.; Shin, N.R.; Jeon, C.M.; Oh, S.R.; Han, S.B.; Ahn, K.S. Anti-inflammatory activities of Physalis alkekengi var. franchetii extract through the inhibition of MMP-9 and AP-1 activation. Immunobiology 2015, 220, 1–9. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef]
- Li, X.; Zhao, J.P.; Yang, M.; Liu, Y.L.; Li, Z.C.; Li, R.Y.; Li, X.R.; Li, N.; Xu, Q.M.; Khan, I.A.; et al. Physalins and withanolides from the fruits of Physalis alkekengi L. var. franchetii (Mast.) Makino and the inhibitory activities against human tumor cells. Phytochem. Lett. 2014, 10, 95–100. [Google Scholar] [CrossRef]
- Li, A.L.; Chen, B.J.; Li, G.H.; Zhou, M.X.; Li, Y.R.; Ren, D.M.; Lou, H.X.; Wang, X.N.; Shen, T. Physalis alkekengi L. var. franchetii (Mast.) Makino: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2018, 210, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Zhang, Q.; Zhang, P.P.; Sun, L.J.; Liang, J.C.; Morris-Natschke, S.L.; Chen, Y.; Lee, K.H. Evaluation of in vitro/in vivo anti-diabetic effects and identification of compounds from Physalis alkekengi. Fitoterapia 2018, 127, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Bose, S.; Kim, Y.M.; Chin, Y.W.; Cho, J. The ethyl acetate fraction from physalis alkekengi inhibits LPS-induced pro-inflammatory mediators in BV2 cells and inflammatory pain in mice. J. Ethnopharmacol. 2016, 181, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Chin, Y.W.; Cho, J. HO-1 dependent antioxidant effects of ethyl acetate fraction from Physalis alkekengi fruit ameliorates scopolamine-induced cognitive impairments. Cell Stress Chaperones 2018, 23, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.Y.; Yang, P.Y.; Zhao, R.; Bai, Y.T.; Guo, Z.J. Matrine Attenuates D-Galactose-Induced Aging-Related Behavior in Mice via Inhibition of Cellular Senescence and Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 7108604. [Google Scholar] [CrossRef] [Green Version]
- Tu, D.G.; Chang, Y.L.; Chou, C.H.; Lin, Y.L.; Chiang, C.C.; Chang, Y.Y.; Chen, Y.C. Preventive effects of taurine against D-galactose-induced cognitive dysfunction and brain damage. Food Funct. 2018, 9, 124–133. [Google Scholar] [CrossRef]
- Xu, L.Q.; Xie, Y.L.; Gui, S.H.; Zhang, X.; Mo, Z.Z.; Sun, C.Y.; Li, C.L.; Luo, D.D.; Zhang, Z.B.; Su, Z.R.; et al. Polydatin attenuates D-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 7, 4545–4555. [Google Scholar] [CrossRef]
- Sha, J.Y.; Zhou, Y.D.; Yang, J.Y.; Leng, J.; Li, J.H.; Hu, J.N.; Liu, W.; Jiang, S.; Wang, Y.P.; Chen, C.; et al. Maltol (3-hydroxy-2-methyl-4-pyrone) slows D-galactose-induced brain aging process by damping the Nrf2/HO-1-mediated oxidative stress in mice. J. Agric. Food Chem. 2019, 67, 10342–10351. [Google Scholar] [CrossRef]
- Ji, M.; Su, X.; Liu, J.; Zhao, Y.; Li, Z.; Xu, X.; Li, H.; Nashun, B. Comparison of naturally aging and D-galactose induced aging model in beagle dogs. Exp. Ther. Med. 2017, 14, 5881–5888. [Google Scholar] [CrossRef] [Green Version]
- Kudryavtsev, B.N.; Kudryavtseva, M.V.; Sakuta, G.A.; Stein, G.I. Human hepatocyte polyploidization kinetics in the course of life cycle. Virchows Archiv. B 1993, 64, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Li, W.J.; Kong, S.Z.; Li, S.D.; Guo, J.Q.; Guo, M.H.; Cai, T.T.; Li, N.; Chen, R.Z.; Luo, R.Q.; et al. Protective effects of collagen polypeptide from tilapia skin against injuries to the liver and kidneys of mice induced by D-galactose. Biomed. Pharmacother. 2019, 117, 109204. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, L.; Zhang, J.; Ran, R.; Shao, Y.; Li, J.; Jia, D.; Zhang, Y.; Zhang, M.; Wang, L.; et al. Protective effects of ginsenoside Rg1 on splenocytes and thymocytes in an aging rat model induced by d-galactose. Int. Immunopharmacol 2018, 58, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Chen, W.; Nong, Z.; Huang, J.; Chen, C. Protective effect of hyperbaric oxygen on cognitive impairment induced by D-galactose in mice. Neurochem. Res. 2016, 41, 3032–3041. [Google Scholar] [CrossRef]
- Ittai, B.P.; Robert, A.W. The signals and pathways activating cellular senescence. Int. J. Biochem. Cell Biol. 2004, 37, 961–976. [Google Scholar]
- Araujo, J.A.; Studzinski, C.M.; Milgram, N.W. Further evidence for the cholinergic hypothesis of aging and dementia from the canine model of aging. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2005, 29, 411–422. [Google Scholar] [CrossRef]
- Huang, T.T.; Zou, Y.; Corniola, R. Oxidative stress and adult neurogenesis-effects of radiation and superoxide dismutase deficiency. Semin. Cell Dev. Biol. 2012, 23, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.T.; Wu, M.F.; Lee, M.C.; Wu, J.H.; Huang, C.C.; Huang, W.C. Antifatigue Activity and Exercise Performance of Phenolic-Rich Extracts from Calendula officinalis, Ribes nigrum, and Vaccinium myrtillus. Nutrients 2019, 11, 1715. [Google Scholar] [CrossRef] [Green Version]
- Duan, F.F.; Guo, Y.; Li, J.W.; Yuan, K. Antifatigue Effect of Luteolin-6-C-Neohesperidoside on Oxidative Stress Injury Induced by Forced Swimming of Rats through Modulation of Nrf2/ARE Signaling Pathways. Oxid. Med. Cell. Longev. 2017, 2017, 3159358. [Google Scholar] [CrossRef]
- Norheim, K.B.; Jonsson, G.; Omdal, R. Biological mechanisms of chronic fatigue. Rheumatology 2011, 50, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, G.; Surana, U.; Clément, M.V. Replication stress-induced endogenous DNA damage drives cellular senescence induced by a sub-lethal oxidative stress. Nucleic Acids Res. 2017, 45, 10564–10582. [Google Scholar] [CrossRef] [PubMed]
- Vendelbo, M.H.; Nair, K.S. Mitochondrial longevity pathways. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 634–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.N.; Guo, Y.; Xi, M.M.; Yang, P.; Zhou, X.Y.; Yin, S.; Hai, C.X.; Li, J.G.; Qin, X.J. Saponins from Aralia taibaiensis Attenuate D-Gal-Induced Aging in Rats by Activating FOXO3a and Nrf2 Pathways. Oxid. Med. Cell. Longev. 2014, 2014, 320513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of malondialdehyde (MDA) by high-performance liquid chromatography in serum and liver as a biomarker for oxidative stress. Application to a rat model for hypercholesterolemia and evaluation of the effect of diets rich in phenolic antioxidants from fruits. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 827, 76–82. [Google Scholar]
- Zhang, Z.; Yang, J.; Liu, C.; Xie, J.; Qiu, S.; Yang, X.; Wu, C. Pseudoginsenoside-F11 alleviates cognitive deficits and Alzheimer’s disease-type pathologies in SAMP8 mice. Int. Immunopharmacol. 2019, 67, 78–86. [Google Scholar] [CrossRef]
- Burton, M.D.; Rytych, J.L.; Amin, R.; Johnson, R.W. Dietary Luteolin Reduces Proinflammatory Microglia in the Brain of Senescent Mice. Rejuvenation Res. 2016, 19, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain Res. Bull. 2015, 119, 1–11. [Google Scholar] [CrossRef]
- Li, T.; Chen, S.; Feng, T.; Dong, J.; Li, Y.; Li, H. Rutin protects against aging-related metabolic dysfunction. Food Funct. 2016, 7, 1147–1154. [Google Scholar] [CrossRef]
- Shen, C.Y.; Jiang, J.G.; Yang, L.; Wang, D.W.; Zhu, W. Anti-ageing active ingredients from herbs and nutraceuticals used in traditional Chinese medicine: Pharmacological mechanisms and implications for drug discovery. Br. J. Pharmacol. 2017, 174, 1395–1425. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, K.; Liu, Q.; Quiles, J.L.; Filosa, R.; Kamal, M.A.; Wang, F.; Kai, G.; Zou, X.; Teng, H.; et al. Protective effects of raspberry on the oxidative damage in HepG2 cells through Keap1/Nrf2-dependent signaling pathway. Food Chem. Toxicol. 2019, 133, 110781. [Google Scholar] [CrossRef]
- Quan, W.; Tao, Y.D.; Qie, X.J.; Zeng, M.M.; Qina, F.; Chen, J.; He, Z.Y. Effects of high-pressure homogenization, thermal processing, and milk matrix on the in vitro bioaccessibility of phenolic compounds in pomelo and kiwi juices. J. Funct. Foods 2020, 64, 103633. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, C.F.; Tian, G.G.; Qian, L.R.; Yu, L.J. Characterization of active constituents in Pyracantha fortuneana fruit extract and their effects on hyperlipidaemia, obesity, and oxidative stress in rodents. J. Funct. Foods 2016, 22, 278–290. [Google Scholar] [CrossRef]
- Sun, K.; Bai, Y.; Zhao, R.; Guo, Z.; Su, X.; Li, P.; Yang, P. Neuroprotective effects of matrine on scopolamine-induced amnesia via inhibition of AChE/BuChE and oxidative stress. Metab. Brain Dis. 2019, 34, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Y.; Cai, G.; Kong, F.; Wang, X.; Liu, Y.; Yang, C.; Wang, D.; Teng, L. Antifatigue activity of liquid cultured tricholoma matsutake mycelium partially via regulation of antioxidant pathway in mouse. BioMed Res. Int. 2015, 2015, 562345. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Components | Content |
|---|---|
| TFC (mg REs/g PAE) | 71.72 ± 2.99 |
| TPC (mg GAEs/g PAE) | 40.19 ± 0.47 |
| TSC (mg OAEs / g PAE) | 128.13 ± 1.04 |
| Rutin (mg/ g PAE) | 1.67 ± 0.07 |
| Luteolin (mg/g PAE) | 1.62 ± 0.01 |
| Weight (g) | Organ Index (g/g %) | |||
|---|---|---|---|---|
| Group | Initial | Final | Thymus | Spleen |
| Control | 34.8 ± 2.1 | 40.3 ± 2.0 | 0.126 ± 0.016 | 0.383 ± 0.018 |
| d-gal | 33.3 ± 1.3 | 38.7 ± 0.7 | 0.070 ± 0.006 ## | 0.401 ± 0.019 |
| VE-100 | 35.2 ± 2.0 | 38.6 ± 1.4 | 0.112 ± 0.011 ** | 0.422 ± 0.057 |
| PAE-3 | 34.8 ± 1.6 | 40.0 ± 0.8 | 0.095 ± 0.007 | 0.407 ± 0.030 |
| PAE-10 | 34.7 ± 1.9 | 40.2 ± 1.4 | 0.103 ± 0.007 * | 0.410 ± 0.014 |
| PAE-30 | 35.4 ± 1.7 | 38.1 ± 1.0 | 0.102 ± 0.006 * | 0.417 ± 0.018 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, K.; Sun, Y.; Li, H.; Han, D.; Bai, Y.; Zhao, R.; Guo, Z. Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 1836. https://doi.org/10.3390/ijms21051836
Sun K, Sun Y, Li H, Han D, Bai Y, Zhao R, Guo Z. Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress. International Journal of Molecular Sciences. 2020; 21(5):1836. https://doi.org/10.3390/ijms21051836
Chicago/Turabian StyleSun, Kaiyue, Yingting Sun, Heyang Li, Dongyao Han, Yuting Bai, Rong Zhao, and Zijiao Guo. 2020. "Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress" International Journal of Molecular Sciences 21, no. 5: 1836. https://doi.org/10.3390/ijms21051836
APA StyleSun, K., Sun, Y., Li, H., Han, D., Bai, Y., Zhao, R., & Guo, Z. (2020). Anti-Ageing Effect of Physalis alkekengi Ethyl Acetate Layer on a d-galactose-Induced Mouse Model through the Reduction of Cellular Senescence and Oxidative Stress. International Journal of Molecular Sciences, 21(5), 1836. https://doi.org/10.3390/ijms21051836