Natural Products Targeting ER Stress, and the Functional Link to Mitochondria



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
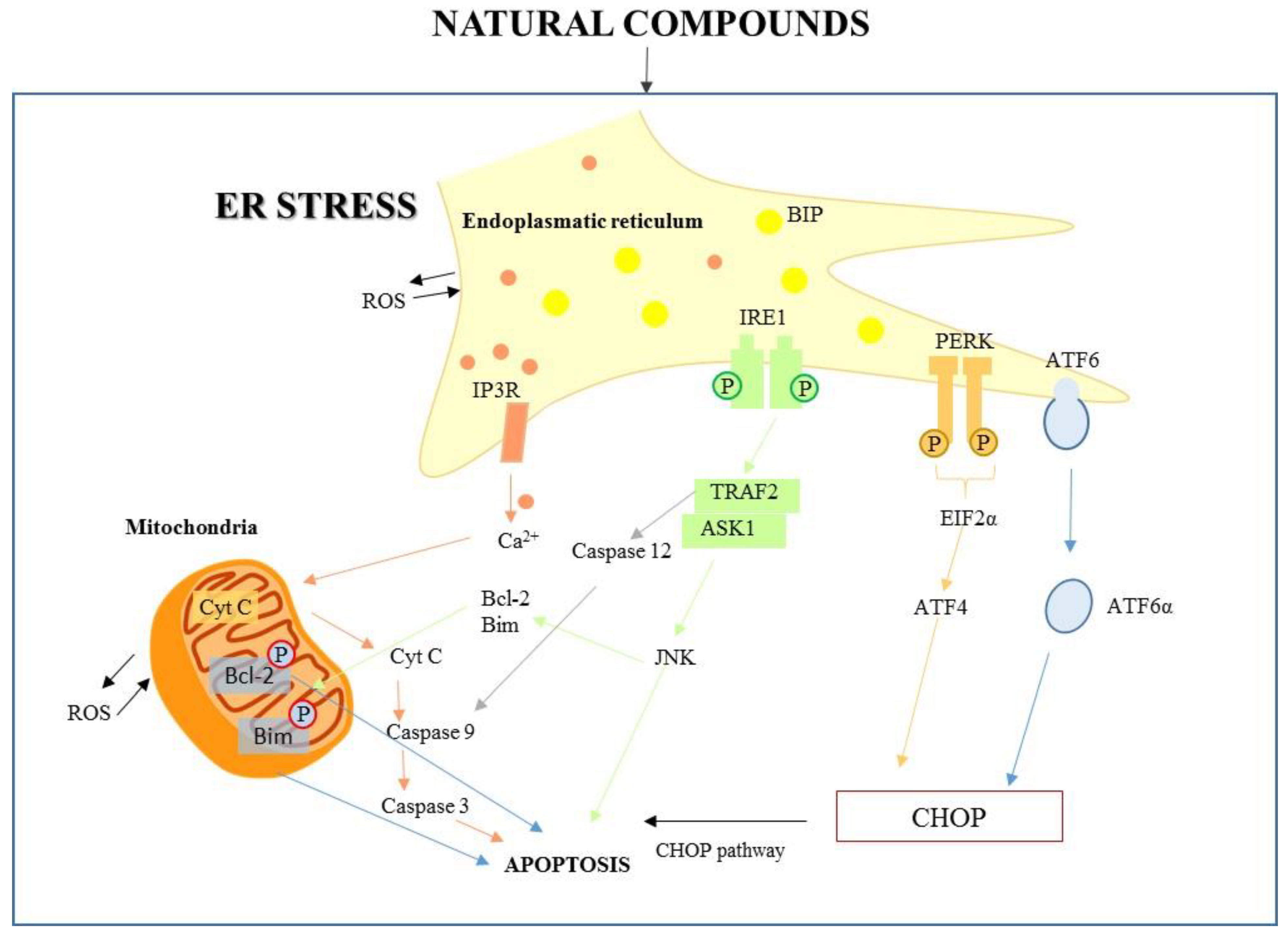
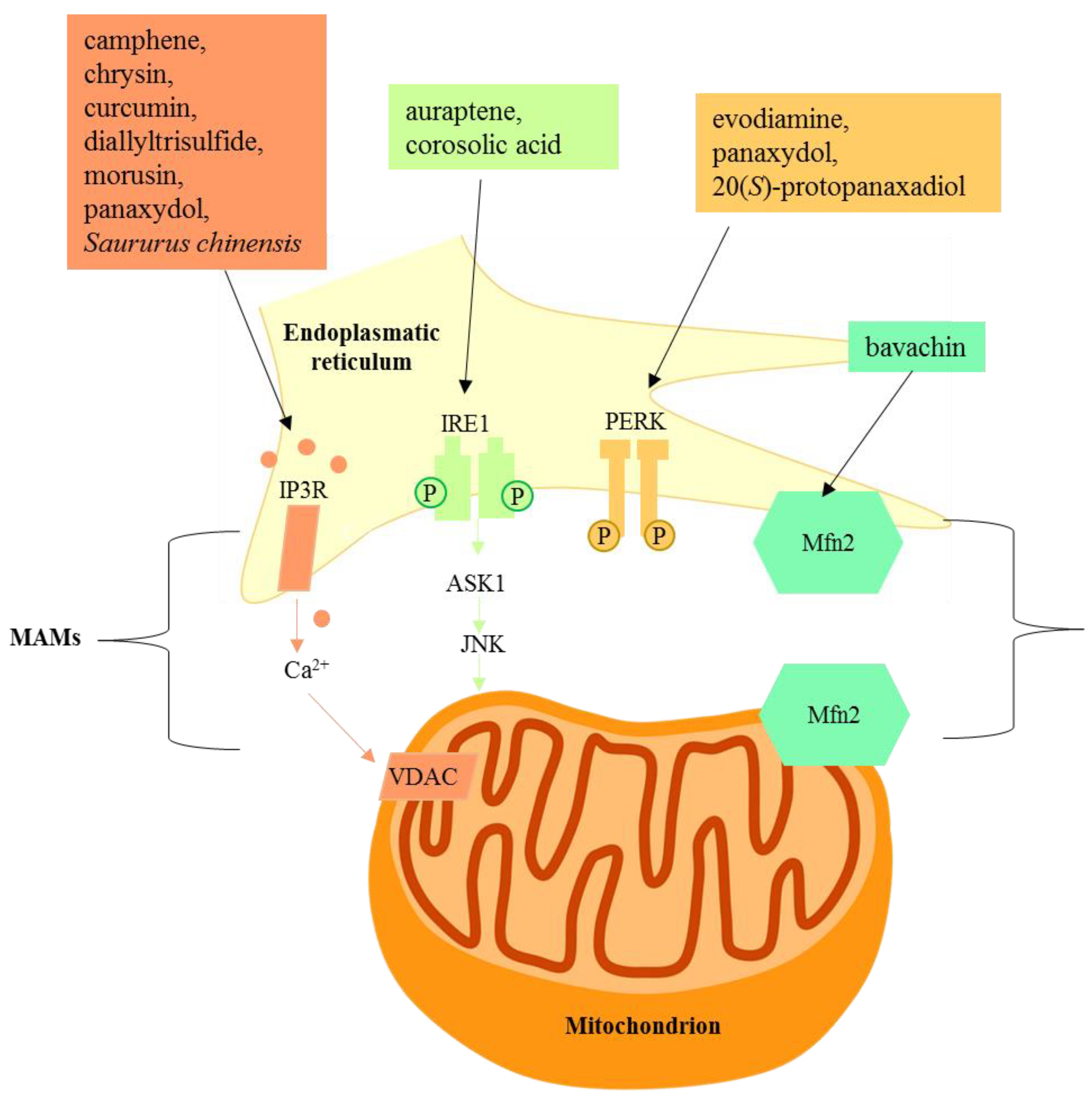
2. ER–Mitochondria Interactions
2.1. ER–Mitochondrial Interaction by Ca2+ Signaling Dependent Mechanisms
2.2. Cross-Talk UPR-Signaling to Mitochondria
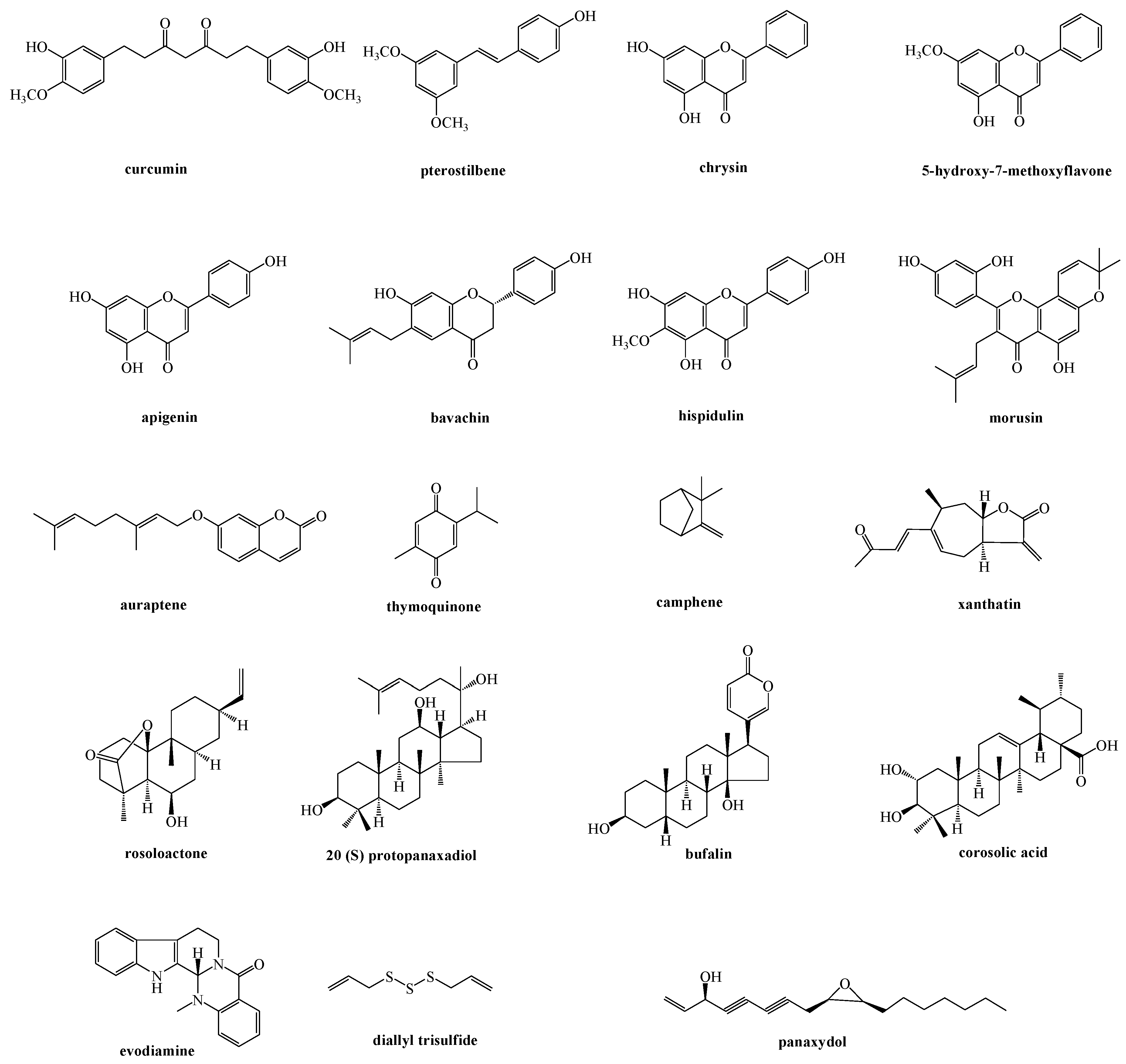
3. Natural Products and the Signaling Communication between ER and Mitochondria
3.1. Flavonoid Derivatives and Phenolic Compounds
3.2. Terpene Derivatives
3.3. Miscellaneous
3.4. Herbal Extracts
4. Natural Compounds Targeting ER Stress-Mediated Apoptosis: “An Update”
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviation
| AMPK | 5′ AMP-activated protein kinase |
| Apaf-1 | Apoptotic protease activating factor-1 |
| ATF4 | Activating transcription factor 4 |
| ATF6 | Activating transcription factor 6 |
| ASK1 | Apoptosis signal-regulating kinase 1 |
| Bak | Bcl-2 homologous antagonist/killer |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| Bcl-xl | B-cell lymphoma-extra large |
| BiP | Binding immunoglobulin protein |
| CHOP | C/EBP homologous protein |
| eIF2 | Eukaryotic initiation factor 2 |
| ER | Endoplasmic reticulum |
| ERAD | ER-associated degradation |
| ERO1 | Endoplasmic reticulum oxireductin 1 |
| GADD34 | Growth arrest and DNA-damage-inducible protein 34 |
| GRP78 | Glucose-regulated protein 78 |
| IRE1 | Inositol requiring enzyme1 |
| JNK | JUN N-terminal kinase |
| MAMs | Mitochondria-associated membranes |
| MCU | Mitochondrial calcium uniporter |
| MFN1 | Mitofusin 1 |
| MFN2 | Mitofusin 2 |
| MMP | Mitochondrial membrane potential |
| mNCX | Mitochondrial Na+/Ca2+ exchanger |
| mTOR | Mammalian target of rapamycin |
| OMM | Outer mitochondrial membrane |
| PERK | Protein kinase R-like ER kinase |
| PUMA | p53 upregulated modulator of apoptosis |
| ROS | Reactive oxygen species |
| RyR | Ryanodine receptors |
| SERCA | Sarcoplasmic/endoplasmic Ca2+ ATPase |
| TNF | Tumor necrosis factor |
| TRAF2 | Tumor necrosis factor receptor-associated factor 2 |
| UPR | Unfolded protein response |
| VDAC | Voltage-dependent anion channel |
| XBP-1 | X-box-binding protein 1 |
References
- Lin, J.H.; Walter, P.; Yen, T.S. Endoplasmic reticulum stress in disease pathogenesis. Annu. Rev. Pathol. 2008, 3, 399–425. [Google Scholar] [CrossRef]
- Ma, Y.; Hendershot, L.M. The role of the unfolded protein response in tumour development: Friend or foe? Nat. Rev. Cancer 2004, 4, 966–977. [Google Scholar] [CrossRef]
- Kincaid, M.M.; Cooper, A.A. Misfolded proteins traffic from the endoplasmic reticulum (ER) due to ER export signals. Mol. Biol. Cell 2007, 18, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Xu, W.J.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef]
- Amodio, G.; Moltedo, O.; Fasano, D.; Zerillo, L.; Oliveti, M.; Di Pietro, P.; Faraonio, R.; Barone, P.; Pellecchia, M.T.; De Rose, A.; et al. PERK-Mediated Unfolded Protein Response Activation and Oxidative Stress in PARK20 Fibroblasts. Front. Neurosci-Switz. 2019, 13. [Google Scholar] [CrossRef] [Green Version]
- Gardner, B.M.; Walter, P. Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science 2011, 333, 1891–1894. [Google Scholar] [CrossRef] [Green Version]
- Asada, R.; Kanemoto, S.; Kondo, S.; Saito, A.; Imaizumi, K. The signalling from endoplasmic reticulum-resident bZIP transcription factors involved in diverse cellular physiology. J. Biochem. 2011, 149, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Martino, M.B.; Jones, L.; Brighton, B.; Ehre, C.; Abdulah, L.; Davis, C.W.; Ron, D.; O’Neal, W.K.; Ribeiro, C.M.P. The ER stress transducer IRE1 beta is required for airway epithelial mucin production. Mucosal Immunol. 2013, 6, 639–654. [Google Scholar] [CrossRef] [Green Version]
- Urra, H.; Dufey, E.; Lisbona, F.; Rojas-Rivera, D.; Hetz, C. When ER stress reaches a dead end. Biochim. Biophys. Acta 2013, 1833, 3507–3517. [Google Scholar] [CrossRef] [Green Version]
- Puthalakath, H.; O’Reilly, L.A.; Gunn, P.; Lee, L.; Kelly, P.N.; Huntington, N.D.; Hughes, P.D.; Michalak, E.M.; McKimm-Breschkin, J.; Motoyama, N.; et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007, 129, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Reimertz, C.; Kogel, D.; Rami, A.; Chittenden, T.; Prehn, J.H. Gene expression during ER stress-induced apoptosis in neurons: Induction of the BH3-only protein Bbc3/PUMA and activation of the mitochondrial apoptosis pathway. J. Cell Biol. 2003, 162, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Malhi, H.; Kaufman, R.J. Endoplasmic reticulum stress in liver disease. J. Hepatol. 2011, 54, 795–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limonta, P.; Moretti, R.M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Marelli, M.M. Role of endoplasmic reticulum stress in the anticancer activity of natural compounds. Int. J. Mol. Sci. 2019, 20, 961. [Google Scholar] [CrossRef] [Green Version]
- Beukes, N.; Levendal, R.-A.; Frost, C.L. Selected terpenoids from medicinal plants modulate endoplasmic reticulum stress in metabolic disorders. J. Pharm. Pharmacol. 2014, 66, 1505–1525. [Google Scholar] [CrossRef] [PubMed]
- Choy, K.W.; Murugan, D.; Mustafa, M.R. Natural products targeting ER stress pathway for the treatment of cardiovascular diseases. Pharmacol. Res. 2018, 132, 119–129. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentao, P.; Correia-da-Silva, G.; Teixeira, N.; Andrade, P.B. Translating endoplasmic reticulum biology into the clinic: A role for ER-targeted natural products? Nat. Prod. Rep. 2015, 32, 705–722. [Google Scholar] [CrossRef]
- Lepretti, M.; Martucciello, S.; Aceves, M.A.B.; Putti, R.; Lionetti, L. Omega-3 Fatty Acids and Insulin Resistance: Focus on the Regulation of Mitochondria and Endoplasmic Reticulum Stress. Nutrients 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Nishitoh, H. Stress responses from the endoplasmic reticulum in cancer. Front. Oncol. 2015, 5, 93. [Google Scholar] [CrossRef] [Green Version]
- Moltedo, O.; Remondelli, P.; Amodio, G. The Mitochondria-Endoplasmic Reticulum Contacts and Their Critical Role in Aging and Age-Associated Diseases. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, X.; Xu, S.; Bao, J.; Yu, H. Curcumin induces endoplasmic reticulum stress-associated apoptosis in human papillary thyroid carcinoma BCPAP cells via disruption of intracellular calcium homeostasis. Medicine 2018, 97, e11095. [Google Scholar] [CrossRef]
- Rizzuto, R.; Pinton, P.; Carrington, W.; Fay, F.S.; Fogarty, K.E.; Lifshitz, L.M.; Tuft, R.A.; Pozzan, T. Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science 1998, 280, 1763–1766. [Google Scholar] [CrossRef]
- Varghese, E.; Samuel, S.M.; Sadiq, Z.; Kubatka, P.; Liskova, A.; Benacka, J.; Pazinka, P.; Kruzliak, P.; Busselberg, D. Anti-Cancer Agents in Proliferation and Cell Death: The Calcium Connection. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Kim, W.H.; Yeo, J.; Jung, M.H. ER stress is implicated in mitochondrial dysfunction-induced apoptosis of pancreatic beta cells. Mol. Cells 2010, 30, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Gilady, S.Y.; Bui, M.; Lynes, E.M.; Benson, M.D.; Watts, R.; Vance, J.E.; Simmen, T. Ero1alpha requires oxidizing and normoxic conditions to localize to the Mitochondria-Associated Membrane (MAM). Cell Stress Chaperones 2010, 15, 619–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, J.P.; Ivanova, S.; Sanchez-Wandelmer, J.; Martinez-Cristobal, P.; Noguera, E.; Sancho, A.; Diaz-Ramos, A.; Hernandez-Alvarez, M.I.; Sebastian, D.; Mauvezin, C.; et al. Mfn2 modulates the UPR and mitochondrial function via repression of PERK. EMBO J. 2013, 32, 2348–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masullo, M.; Montoro, P.; Mari, A.; Pizza, C.; Piacente, S. Medicinal plants in the treatment of women’s disorders: Analytical strategies to assure quality, safety and efficacy. J. Pharm. Biomed. Anal. 2015, 113, 189–211. [Google Scholar] [CrossRef]
- Benavides, A.; Montoro, P.; Bassarello, C.; Piacente, S.; Pizza, C. Catechin derivatives in Jatropha macrantha stems: Characterisation and LC/ESI/MS/MS quali-quantitative analysis. J. Pharm. Biomed. 2006, 40, 639–647. [Google Scholar] [CrossRef]
- Cerulli, A.; Lauro, G.; Masullo, M.; Cantone, V.; Olas, B.; Kontek, B.; Nazzaro, F.; Bifulco, G.; Piacente, S. Cyclic Diarylheptanoids from Corylus avellana Green Leafy Covers: Determination of Their Absolute Configurations and Evaluation of Their Antioxidant and Antimicrobial Activities. J. Nat. Prod. 2017, 80, 1703–1713. [Google Scholar] [CrossRef]
- Masullo, M.; Mari, A.; Cerulli, A.; Bottone, A.; Kontek, B.; Olas, B.; Pizza, C.; Piacente, S. Quali-quantitative analysis of the phenolic fraction of the flowers of Corylus avellana, source of the Italian PGI product “Nocciola di Giffoni”: Isolation of antioxidant diarylheptanoids. Phytochemistry 2016, 130, 273–281. [Google Scholar] [CrossRef]
- Xue, J.; Li, R.; Zhao, X.; Ma, C.; Lv, X.; Liu, L.; Liu, P. Morusin induces paraptosis-like cell death through mitochondrial calcium overload and dysfunction in epithelial ovarian cancer. Chem. Biol. Interact. 2018, 283, 59–74. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, X.; Haa, F.; Ma, Z.; Wang, Y.; Wang, L.; Gao, Y. Bavachin induces apoptosis through mitochondria! regulated ER stress pathway in HepG2 cells. Biol. Pharm. Bull. 2018, 41, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, S.; Lim, W.; Song, G.; Lim, W.; Bazer, F.W. Chrysin induces death of prostate cancer cells by inducing ROS and ER stress. J. Cell. Physiol. 2017, 232, 3786–3797. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.Y.; Kim, J.S.; Park, H.S.; Han, C.R.; Fang, Z.; Woo, M.H.; Rhee, I.K.; Kim, Y.H. Apoptogenic activity of auraptene of Zanthoxylum schinifolium toward human acute leukemia Jurkat T cells is associated with ER stress-mediated caspase-8 activation that stimulates mitochondria-dependent or -independent caspase cascade. Carcinogenesis 2007, 28, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Iranshahi, M.; Chini, M.G.; Masullo, M.; Sahebkar, A.; Javidnia, A.; Yazdi, M.C.; Pergola, C.; Koeberle, A.; Werz, O.; Pizza, C.; et al. Can Small Chemical Modifications of Natural Pan-inhibitors Modulate the Biological Selectivity? The Case of Curcumin Prenylated Derivatives Acting as HDAC or mPGES-1 Inhibitors. J. Nat. Prod. 2015, 78, 2867–2879. [Google Scholar] [CrossRef]
- Szebeni, G.J.; Balazs, A.; Madarasz, I.; Pocz, G.; Ayaydin, F.; Kanizsai, I.; Fajka-Boja, R.; Alfoldi, R.; Hackler, L.; Puskas, L.G. Achiral Mannich-Base Curcumin Analogs Induce Unfolded Protein Response and Mitochondrial Membrane Depolarization in PANC-1 Cells. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Masullo, M.; Pizza, C.; Piacente, S. Oleanane derivatives for pharmaceutical use: A patent review (2000–2016). Expert Opin. Ther. Pat. 2017, 27, 237–255. [Google Scholar] [CrossRef]
- Girola, N.; Figueiredo, C.R.; Farias, C.F.; Azevedo, R.A.; Ferreira, A.K.; Teixeira, S.F.; Capello, T.M.; Martins, E.G.A.; Matsuo, A.L.; Travassos, L.R.; et al. Camphene isolated from essential oil of Piper cernuum (Piperaceae) induces intrinsic apoptosis in melanoma cells and displays antitumor activity in vivo. Biochem. Biophys. Res. Commun. 2015, 467, 928–934. [Google Scholar] [CrossRef]
- Zhou, L.; Qin, J.; Ma, L.; Li, H.; Li, L.; Ning, C.; Gao, W.; Yu, H.; Han, L. Rosoloactone: A natural diterpenoid inducing apoptosis in human cervical cancer cells through endoplasmic reticulum stress and mitochondrial damage. Biomed. Pharmacother. 2017, 95, 355–362. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, H.; Wang, Y.; Zhao, A.; Zhu, Z.; Bao, X.; Sun, Y.; Zhang, Q.; Zhang, H.; Li, L. Corosolic acid, a natural triterpenoid, induces ER stress-dependent apoptosis in human castration resistant prostate cancer cells via activation of IRE-1/JNK, PERK/CHOP and TRIB3. J. Exp. Clin. Cancer Res. 2018, 37, 210. [Google Scholar] [CrossRef]
- Zhu, G.-Y.; Li, Y.-W.; Tse, A.K.-W.; Hau, D.K.-P.; Leung, C.-H.; Yu, Z.-L.; Fong, W.-F. 20(S)-Protopanaxadiol, a metabolite of ginsenosides, induced cell apoptosis through endoplasmic reticulum stress in human hepatocarcinoma HepG2 cells. Eur. J. Pharmacol. 2011, 668, 88–98. [Google Scholar] [CrossRef]
- Chen, T.-C.; Chien, C.-C.; Wu, M.-S.; Chen, Y.-C. Evodiamine from Evodia rutaecarpa induces apoptosis via activation of JNK and PERK in human ovarian cancer cells. Phytomedicine 2016, 23, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lim, J.M.; Kim, J.Y.; Kim, Y.; Park, S.; Sohn, J. Panaxydol, a component of Panax ginseng, induces apoptosis in cancer cells through EGFR activation and ER stress and inhibits tumor growth in mouse models. Int. J. Cancer 2016, 138, 1432–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-C.; Hsieh, S.-C.; Yang, J.-H.; Lin, S.-Y.; Sheen, L.-Y. Diallyl Trisulfide Induces Apoptosis of Human Basal Cell Carcinoma Cells via Endoplasmic Reticulum Stress and the Mitochondrial Pathway. Nutr. Cancer 2012, 64, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Han, Y.-A.; Kim, J.-E.; Hong, S.-H.; Park, E.-J.; Cho, M.-H. Saururus chinensis Baill induces apoptosis through endoplasmic reticulum stress in HepG2 hepatocellular carcinoma cells. Food Chem. Toxicol. 2015, 83, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Nishihara, H.; Hara, H.; Adachi, T. Ethanol extract of Brazilian red propolis induces apoptosis in human breast cancer MCF-7 cells through endoplasmic reticulum stress. J. Agric. Food Chem. 2012, 60, 11065–11070. [Google Scholar] [CrossRef]
- Fu, K.; Chen, L.; Zhang, W.; Bai, Y.; Miao, L.; Guo, Y. Grape Seed Proanthocyanidins Protect N2a Cells against Ischemic Injury via Endoplasmic Reticulum Stress and Mitochondrial-associated Pathways. CNS Neurol. Disord. Drug Targets 2019, 18, 334–341. [Google Scholar] [CrossRef]
- Park, S.; Song, G.; Park, S.; Song, G.; Lim, W.; Bazer, F.W. Apigenin induces ROS-dependent apoptosis and ER stress in human endometriosis cells. J. Cell. Physiol. 2018, 233, 3055–3065. [Google Scholar] [CrossRef]
- Han, M.; Gao, H.; Gao, M.-Q.; Liu, K.-L.; Xie, J.; Chen, X.-H.; Han, Y.-T.; Yuan, Y.-P.; Yuan, Y.-P.; Yuan, Q.; et al. Hispidulin induces ER stress-mediated apoptosis in human hepatocellular carcinoma cells in vitro and in vivo by activating AMPK signaling pathway. Acta Pharmacol. Sin. 2019, 40, 666–676. [Google Scholar] [CrossRef]
- Quan, Z.; Gu, J.; Dong, P.; Lu, J.; Wu, X.; Wu, W.; Fei, X.; Li, S.; Wang, Y.; Wang, J.; et al. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to cirsimaritin-induced apoptosis in human gallbladder carcinoma GBC-SD cells. Cancer Lett. (Shannon, Irel.) 2010, 295, 252–259. [Google Scholar] [CrossRef]
- Shi, T.-L.; Zhang, L.; Feng, X.-J.; Cheng, Q.-Y.; Liu, J.; Shen, Y.-J.; Yu, J.-S.; Shen, Y.-X. Xanthatin induces apoptosis by activating endoplasmic reticulum stress in hepatoma cells. Eur. J. Pharmacol. 2019, 843, 1–11. [Google Scholar] [CrossRef]
- Bouhlel, A.; Mosbah, B.; Abdallah, H.; Ribault, C.; Viel, R.; Mannai, S.; Corlu, A.; Abdennebi, B. Thymoquinone prevents endoplasmic reticulum stress and mitochondria-induced apoptosis in a rat model of partial hepatic warm ischemia reperfusion. Biomed. Pharmacother. 2017, 94, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, F.; Lv, Z.M.; Shi, W.Q.; Zhang, L.Y.; Yan, M. Triptolide disrupts the actin-based Sertoli-germ cells adherens junctions by inhibiting Rho GTPases expression. Toxicol. Appl. Pharmacol. 2016, 310, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Martucciello, S.; Lavric, M.; Toth, B.; Korponay-Szabo, I.; Nadalutti, C.; Myrsky, E.; Rauhavirta, T.; Esposito, C.; Sulic, A.M.; Sblattero, D.; et al. RhoB is associated with the anti-angiogenic effects of celiac patient transglutaminase 2-targeted autoantibodies. J. Mol. Med. 2012, 90, 817–826. [Google Scholar] [CrossRef]
- Bailly, C.; Waring, M.J. Pharmacological effectors of GRP78 chaperone in cancers. Biochem. Pharmacol. 2019, 163, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Mujumdar, N.; Banerjee, S.; Chen, Z.; Sangwan, V.; Chugh, R.; Dudeja, V.; Yamamoto, M.; Vickers, S.M.; Saluja, A.K. Triptolide activates unfolded protein response leading to chronic ER stress in pancreatic cancer cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G1011–G1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piacente, S.; Masullo, M.; De Neve, N.; Dewelle, J.; Hamed, A.; Kiss, R.; Mijatovic, T. Cardenolides from Pergularia tomentosa display cytotoxic activity resulting from their potent inhibition of Na+/K+-ATPase. J. Nat. Prod. 2009, 72, 1087–1091. [Google Scholar] [CrossRef]
- Hosseini, S.H.; Masullo, M.; Cerulli, A.; Martucciello, S.; Ayyari, M.; Pizza, C.; Piacente, S. Antiproliferative Cardenolides from the Aerial Parts of Pergularia tomentosa. J. Nat. Prod. 2019, 82, 74–79. [Google Scholar] [CrossRef]
- Hamed, A.I.; Plaza, A.; Balestrieri, M.L.; Mahalel, U.A.; Springuel, I.V.; Oleszek, W.; Pizza, C.; Piacente, S. Cardenolide glycosides from Pergularia tomentosa and their proapoptotic activity in Kaposi’s sarcoma cells. J. Nat. Prod. 2006, 69, 1319–1322. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, Y.; Wang, Z.; Gong, X.; Liu, R. Bufalin induces the interplay between apoptosis and autophagy in glioma cells through endoplasmic reticulum stress. Int. J. Biol. Sci. 2014, 10, 212–224. [Google Scholar] [CrossRef]
- Martucciello, S.; Paolella, G.; Muzashvili, T.; Skhirtladze, A.; Pizza, C.; Caputo, I.; Piacente, S. Steroids from Helleborus caucasicus reduce cancer cell viability inducing apoptosis and GRP78 down-regulation. Chem. Biol. Interact. 2018, 279, 43–50. [Google Scholar] [CrossRef]
- Li, J.; Xia, X.; Ke, Y.; Nie, H.; Smith, M.A.; Zhu, X. Trichosanthin induced apoptosis in HL-60 cells via mitochondrial and endoplasmic reticulum stress signaling pathways. Biochim. Biophys. Acta 2007, 1770, 1169–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martucciello, S.; Masullo, M.; Cerulli, A.; Piacente, S. Natural Products Targeting ER Stress, and the Functional Link to Mitochondria. Int. J. Mol. Sci. 2020, 21, 1905. https://doi.org/10.3390/ijms21061905
Martucciello S, Masullo M, Cerulli A, Piacente S. Natural Products Targeting ER Stress, and the Functional Link to Mitochondria. International Journal of Molecular Sciences. 2020; 21(6):1905. https://doi.org/10.3390/ijms21061905
Chicago/Turabian StyleMartucciello, Stefania, Milena Masullo, Antonietta Cerulli, and Sonia Piacente. 2020. "Natural Products Targeting ER Stress, and the Functional Link to Mitochondria" International Journal of Molecular Sciences 21, no. 6: 1905. https://doi.org/10.3390/ijms21061905
APA StyleMartucciello, S., Masullo, M., Cerulli, A., & Piacente, S. (2020). Natural Products Targeting ER Stress, and the Functional Link to Mitochondria. International Journal of Molecular Sciences, 21(6), 1905. https://doi.org/10.3390/ijms21061905