Overexpression of Barley Transcription Factor HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
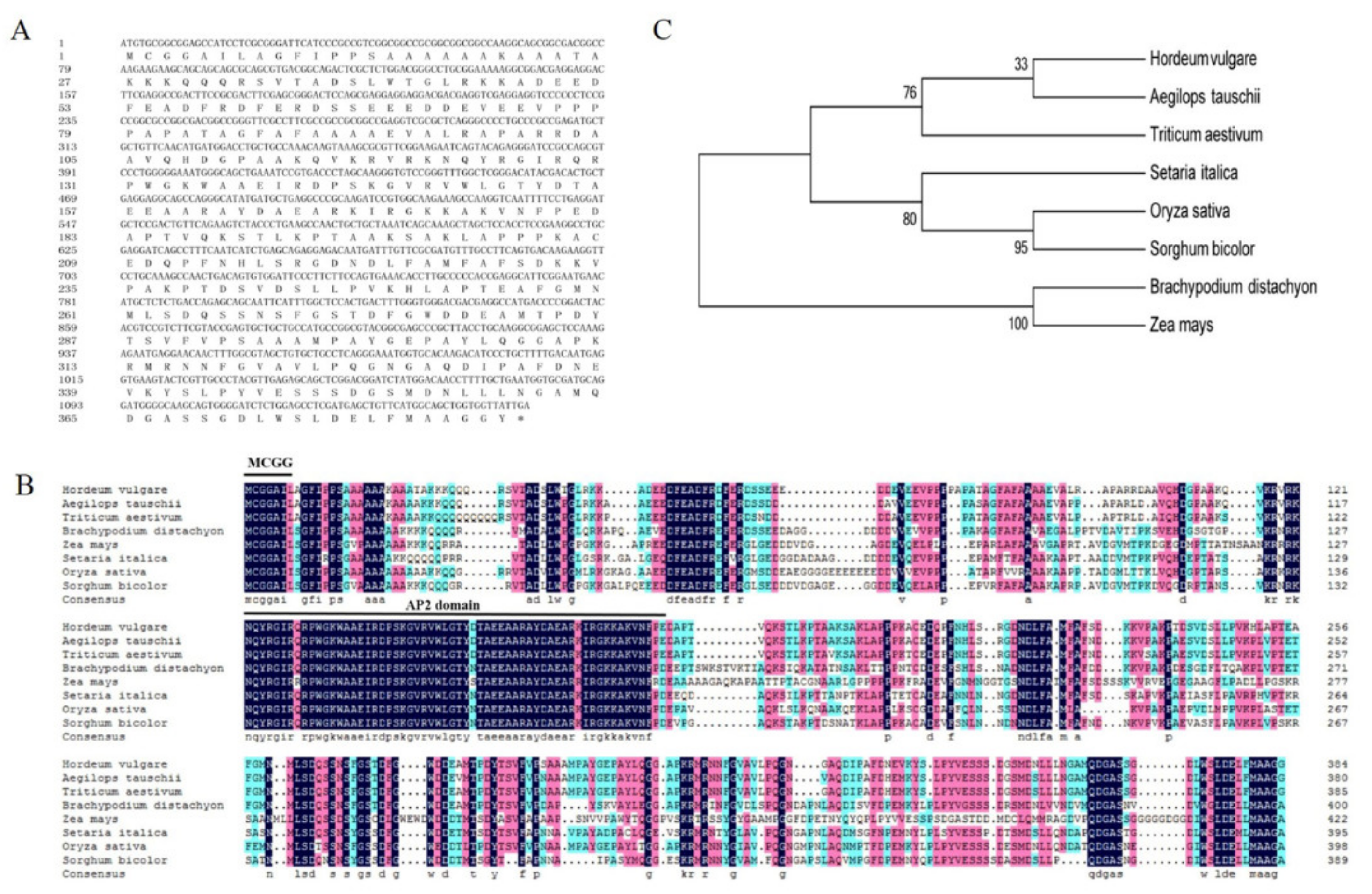
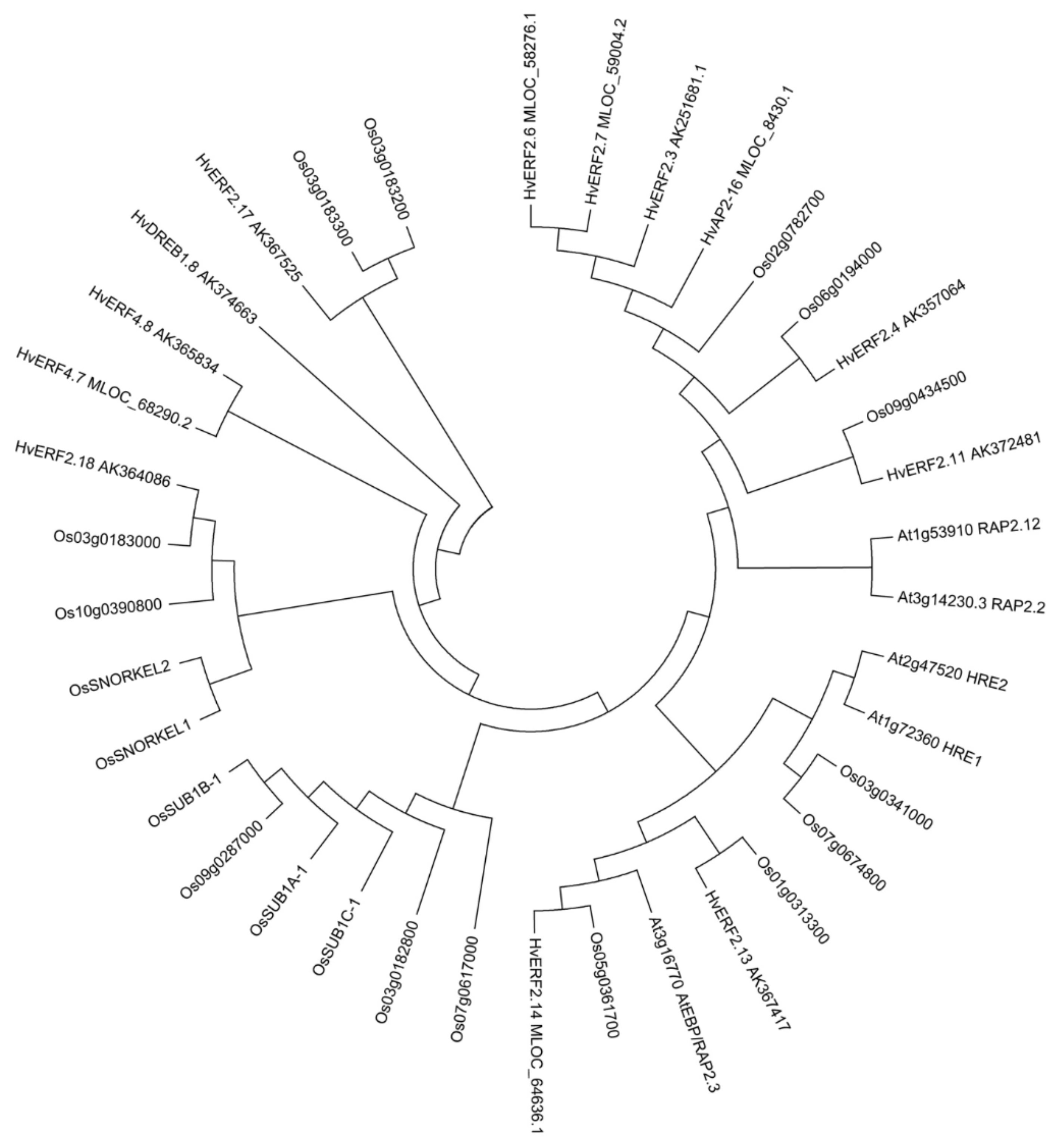
2.1. Sequence Feature of HvERF2.11
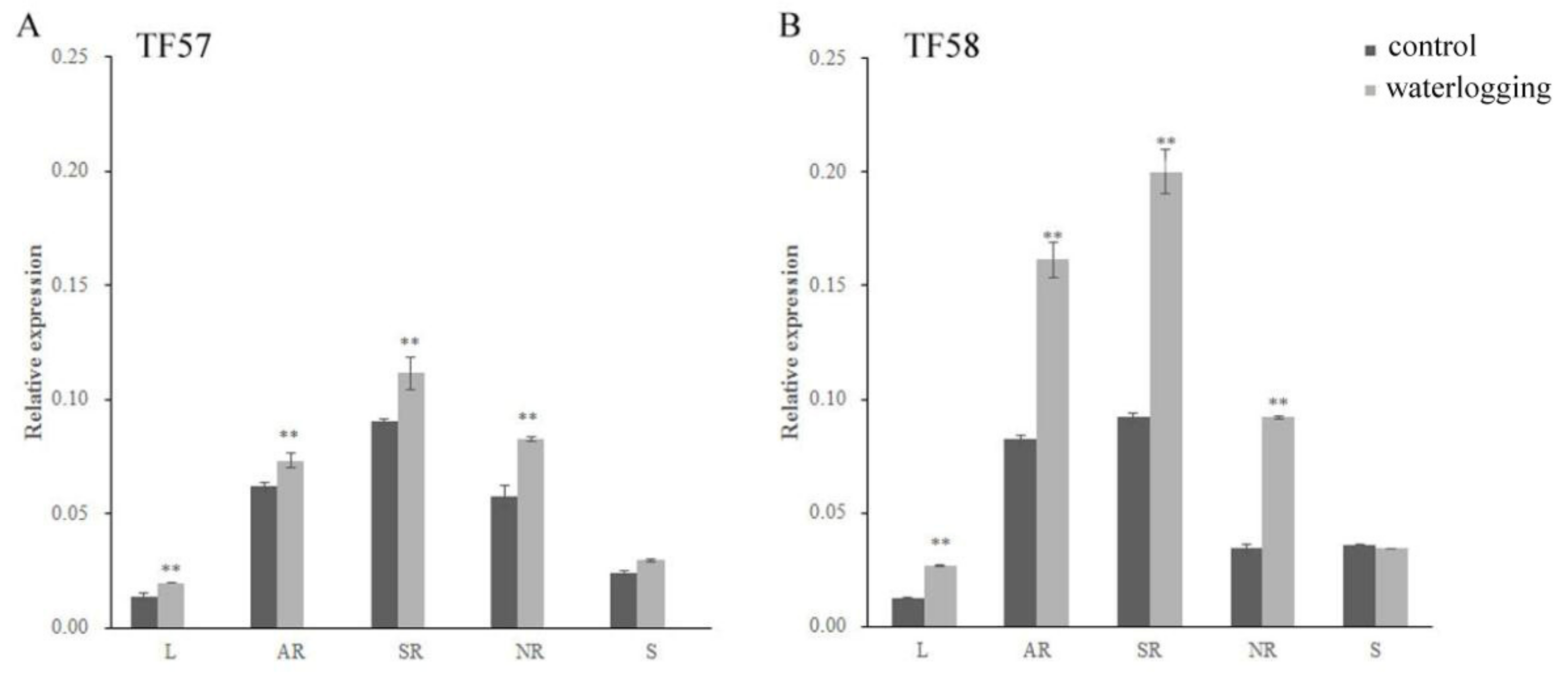
2.2. Expression of the HvERF2.11 Gene under Waterlogging Stress
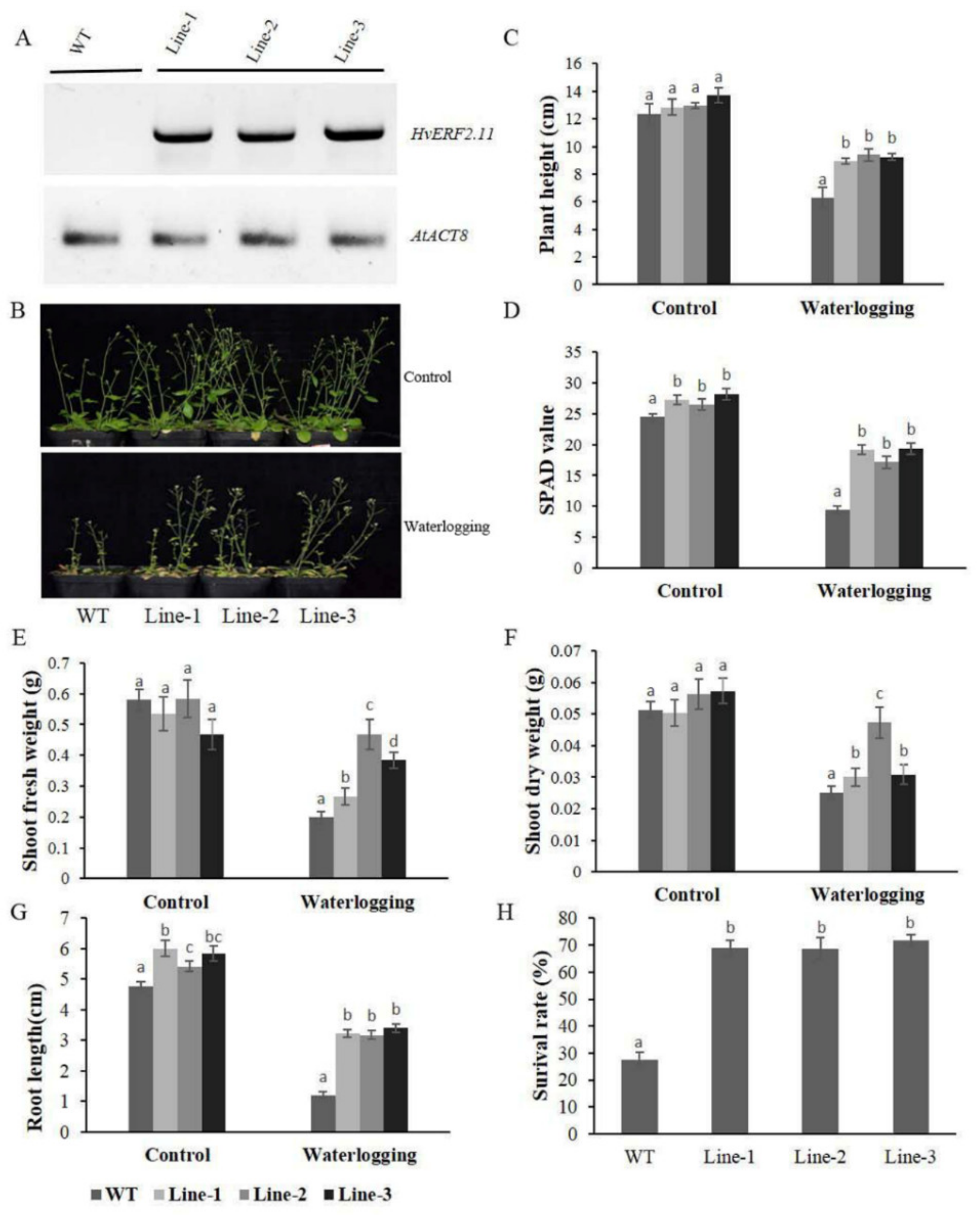
2.3. Overexpression of HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance
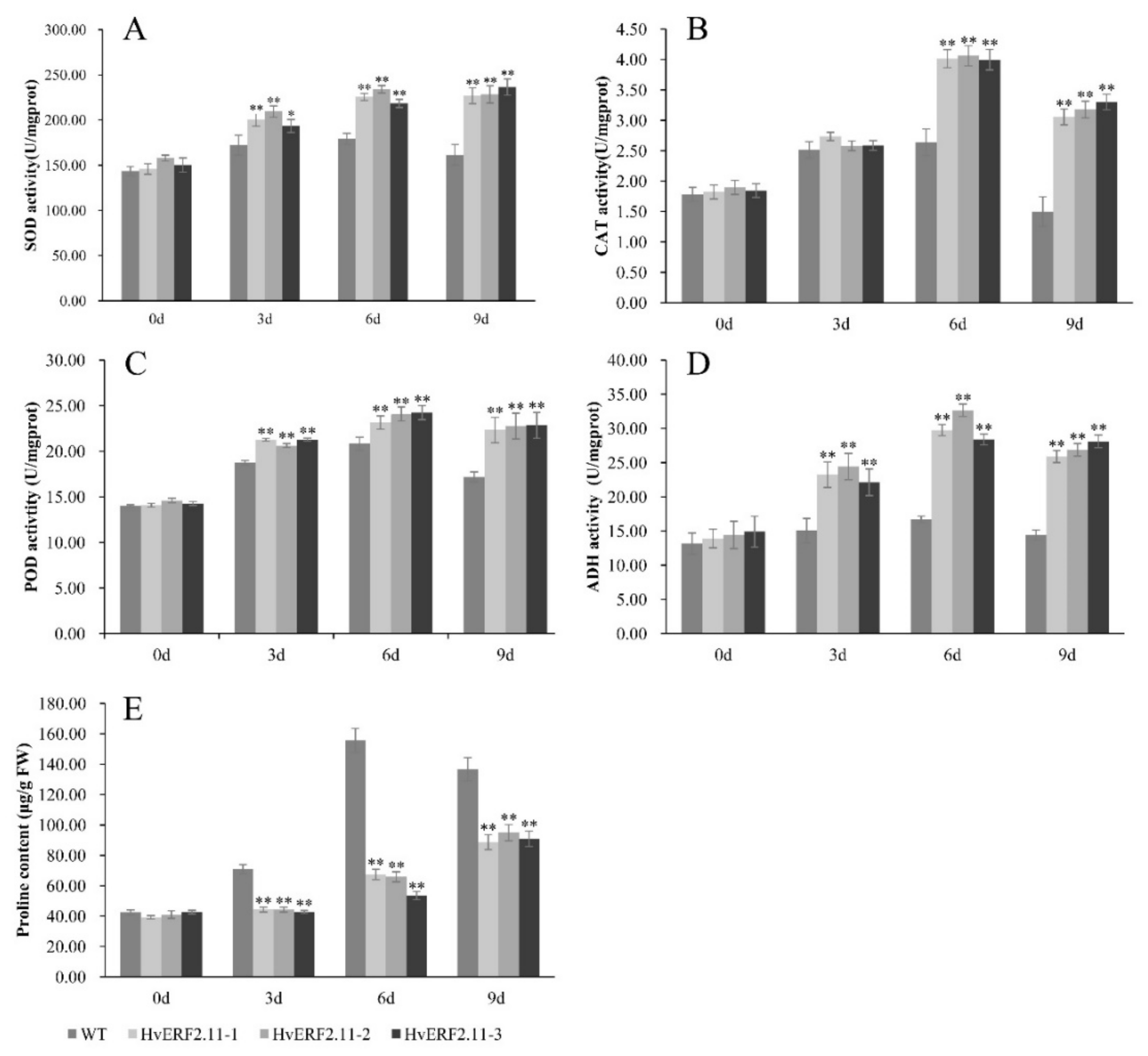
2.4. Overexpressing of HvERF2.11 Increased the Activities of Antioxidant Enzymes
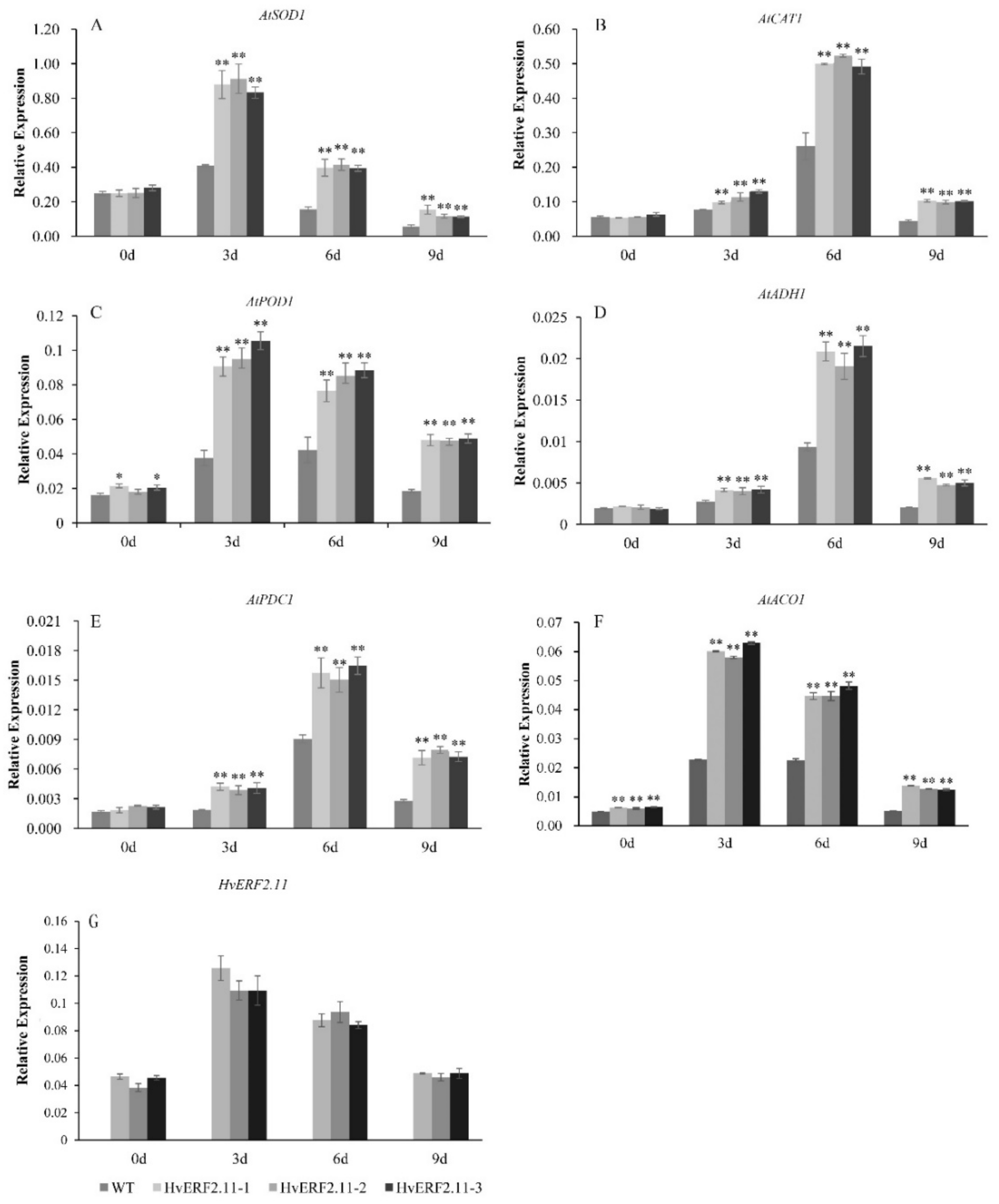
2.5. Overexpression of HvERF2.11 Increases the Expression of Stress-related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Waterlogging Treatment
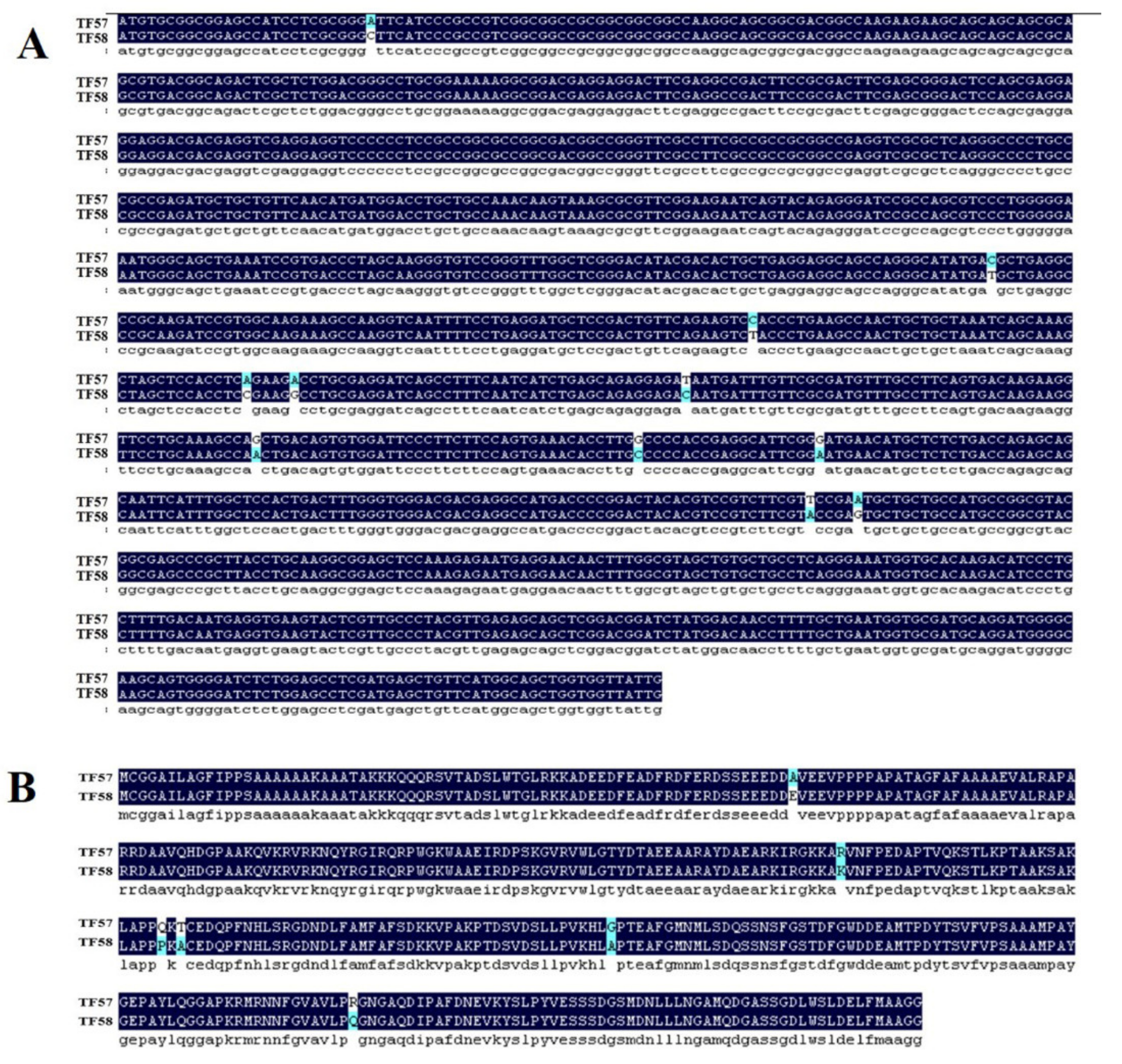
4.2. Total RNA Isolation and HvERF2.11 Gene Cloning
4.3. Sequence Analysis of HvERF2.11 Gene
4.4. Gene Expression Analysis Using qRT-PCR
4.5. Plant Transformation of HvERF2.11
4.6. Analysis of Transgenic Lines for Waterlogging Tolerance
4.7. Detection of the Physiological Parameters Involved in Waterlogging Tolerance
4.8. Expression Analysis of Waterlogging-related Genes in Transgenic Lines
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Armstrong, W.; Drew, M.C. Root Growth and Metabolism under Oxygen Deficiency. Plant Roots: The Hidden Half, 3rd ed.; Marcel Dekker: New York, NY, USA, 2002; pp. 729–761. [Google Scholar]
- Caruso, P.; Baldoni, E.; Mattana, M.; Pietro, P.D.; Genga, A.; Coraggio, I.; Russo, G.; Picchi, V.; Reforgiato, R.G.; Locatelli, F. Ectopic expression of a rice transcription factor, Mybleu, enhances tolerance of transgenic plants of Carrizo citrange to low oxygen stress. Plant. Cell 2012, 109, 327–339. [Google Scholar] [CrossRef]
- Edwards, J.M.; Roberts, T.H.; Atwell, B.J. Quantifying ATP turnover in anoxic coleoptiles of rice (Oryza sativa) demonstrates preferential allocation of energy to protein synthesis. J. Exp. Bot. 2012, 63, 4389–4402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant. Physiol. Biochem. 2012, 58, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Choi, D. Characterization of genes encoding ABA 8’-hydroxylase in ethylene-induced stem growth of deepwater rice (Oryza sativa L.). Biochem. Biophys. Res. Commun. 2006, 350, 685–690. [Google Scholar] [CrossRef]
- He, C.J.; Morgan, P.W.; Drew, M.C. Transduction of an ethylene signal is required for cell death and lysis in the root cortex of maize during aerenchyma formation induced by hypoxia. Plant. Physiol. 1996, 112, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Gu, M.; Cong, Y.; Zou, C.; Zhang, X.; Wang, H. Combining ability and genetic effects of germination traits of Brassica napus L. under waterlogging stress condition. Agric. Sci. China 2010, 9, 101–105. [Google Scholar] [CrossRef]
- Klok, E.J.; Wilson, I.W.; Wilson, D.; Chaoman, S.C.; Ewing, R.W.; Somerville, S.C.; Peacocl, W.J.; Dolferus, R.; Dennis, E.S. Expression profile analysis of the Low-Oxygen response in arabidopsis root cultures. Plant. Cell 2002, 14, 2481–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, Y.; Lu, H.; Kong, X.; Dai, J.; Li, Z. Growth, lint yield and changes in physiological attributes of cotton under temporal waterlogging. Field Crops Res. 2016, 194, 83–93. [Google Scholar] [CrossRef]
- Zhang, P.; Lyu, D.G.; Jia, L.T.; He, J.L.; Qin, S.J. Physiological and de novo transcriptome analysis of the fermentation mechanism of Cerasus sachalinensis roots in response to short-term waterlogging. BMC Genom. 2017, 18, 649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, Z.H.; Zheng, L.L.; Wang, J.; Gao, Z.; Wu, S.B.; Qi, Z.; Wang, Y.C. Transcriptomic profiling of the salt-stress response in the wild recretohalophyte Reaumuria trigyna. BMC Genom. 2013, 14, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hu, Y.X.; Li, X.L.; Yu, X.D.; Li, Q.L. Molecular characterization and function analysis of SlNAC2 in Suaeda liaotungensis K. Gene 2014, 543, 190–197. [Google Scholar] [CrossRef]
- Xu, X.; Chen, M.; Ji, J.; Xu, Q.; Qi, X.; Chen, X. Comparative RNA-seq based transcriptome profiling of waterlogging response in cucumber hypocotyls reveals novel insights into the de novo adventitious root primordia initiation. BMC Plant. Biol. 2017, 17, 129. [Google Scholar] [CrossRef] [Green Version]
- Wessler, S.R. Homing into the origin of the AP2 DNA binding domain. Trends in Plant. Sci. 2005, 10, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold inducible gene expression. Biochem. Bioph. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant. Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosmacz, M.; Parlanti, S.; Schwarzländer, M.; Kragler, F.; Licausi, F.; Van Dongen, J.T. The stability and nuclear localization of the transcription factor RAP2.12 are dynamically regulated by oxygen concentration. Plant. Cell Environ. 2015, 38, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: An ethylene response transcription factor that is important for hypoxia survival. Plant. Physiol. 2010, 153, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, D.; Lee, S.; Isa, N.; Gramuglia, S.; Fukao, T.; Bassel, G.; Correia, C.; Corbineau, F.; Theodoulou, F.; Bailey-Serres, J. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef]
- Licausi, F.; Van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant. J. 2010, 62, 302–315. [Google Scholar] [PubMed]
- Papdi, C.; Pérez-Salamó, I.; Joseph, M.P.; Giuntoli, B.; Bögre, L.; Koncz, C.; Szabados, L. The low oxygen, oxidative and osmotic stress responses synergistically act through the ethylene response factor VII genes RAP2.12, RAP2.2 and RAP2.3. Plant. J. 2015, 82, 772–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant. Cell. 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukao, T.; Bailey-Serres, J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romina, P.; Abeledo, L.G.; Miralles, D.J. Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant. Soil 2014, 378, 265–277. [Google Scholar]
- Luan, H.Y.; Guo, B.J.; Pan, Y.H.; Lv, C.; Shen, H.Q.; Xu, R.G. Morpho-anatomical and physiological responses to waterlogging stress in different barley (Hordeum vulgare L.) genotypes. Plant. Growth Regul. 2018, 85, 399–409. [Google Scholar] [CrossRef]
- Guo, B.J.; Wei, Y.F.; Xu, R.B.; Lin, S.; Luan, H.Y.; Lv, C.; Zhang, X.Z.; Song, X.Y.; Xu, R.G. Genome-Wide analysis of APETALA2/ethylene-responsive factor (AP2/ERF) gene family in barley (Hordeum vulgare L.). PLoS ONE 2016, 11, e0161322. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Huang, S.N.; Wang, G.; Xuan, J.P.; Guo, Z.R. Overexpression of Actinidia deliciosa pyruvate decarboxylase 1 gene enhances waterlogging stress in transgenic Arabidopsis thaliana. Plant. Physiol. Bioch. 2016, 106, 244–252. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Huang, S.N.; Chen, Y.H.; Wang, G.; Guo, Z.R. Identification and characterization of two waterlogging responsive alcohol dehydrogenase genes (AdADH1 and AdADH2) in Actinidia deliciosa. Mol. Breed. 2017, 37, 52–65. [Google Scholar] [CrossRef]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant. Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Gao, F.; Zhou, J.; Deng, R.Y.; Zhao, H.X.; Li, C.L.; Chen, H.; Suzuki, T.; Park, S.U.; Wu, Q. Overexpression of a tartary buckwheat R2R3-MYB transcription factor gene, FtMYB9, enhances tolerance to drought and salt stresses in transgenic Arabidopsis. J. Plant. Physiol. 2017, 214, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.Y.; Shen, H.Q.; Pan, Y.H.; Guo, B.J.; Lv, C.; Xu, R.G. Elucidating the hypoxic stress response in barley (Hordeum vulgare L.) during waterlogging: A proteomics approach. Sci. Rep. 2018, 8, 9655. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtold, N.; Pelletier, G. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods. Mol. Biol. 1998, 82, 259–266. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant. J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luan, H.; Guo, B.; Shen, H.; Pan, Y.; Hong, Y.; Lv, C.; Xu, R. Overexpression of Barley Transcription Factor HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance. Int. J. Mol. Sci. 2020, 21, 1982. https://doi.org/10.3390/ijms21061982
Luan H, Guo B, Shen H, Pan Y, Hong Y, Lv C, Xu R. Overexpression of Barley Transcription Factor HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance. International Journal of Molecular Sciences. 2020; 21(6):1982. https://doi.org/10.3390/ijms21061982
Chicago/Turabian StyleLuan, Haiye, Baojian Guo, Huiquan Shen, Yuhan Pan, Yi Hong, Chao Lv, and Rugen Xu. 2020. "Overexpression of Barley Transcription Factor HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance" International Journal of Molecular Sciences 21, no. 6: 1982. https://doi.org/10.3390/ijms21061982
APA StyleLuan, H., Guo, B., Shen, H., Pan, Y., Hong, Y., Lv, C., & Xu, R. (2020). Overexpression of Barley Transcription Factor HvERF2.11 in Arabidopsis Enhances Plant Waterlogging Tolerance. International Journal of Molecular Sciences, 21(6), 1982. https://doi.org/10.3390/ijms21061982