Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
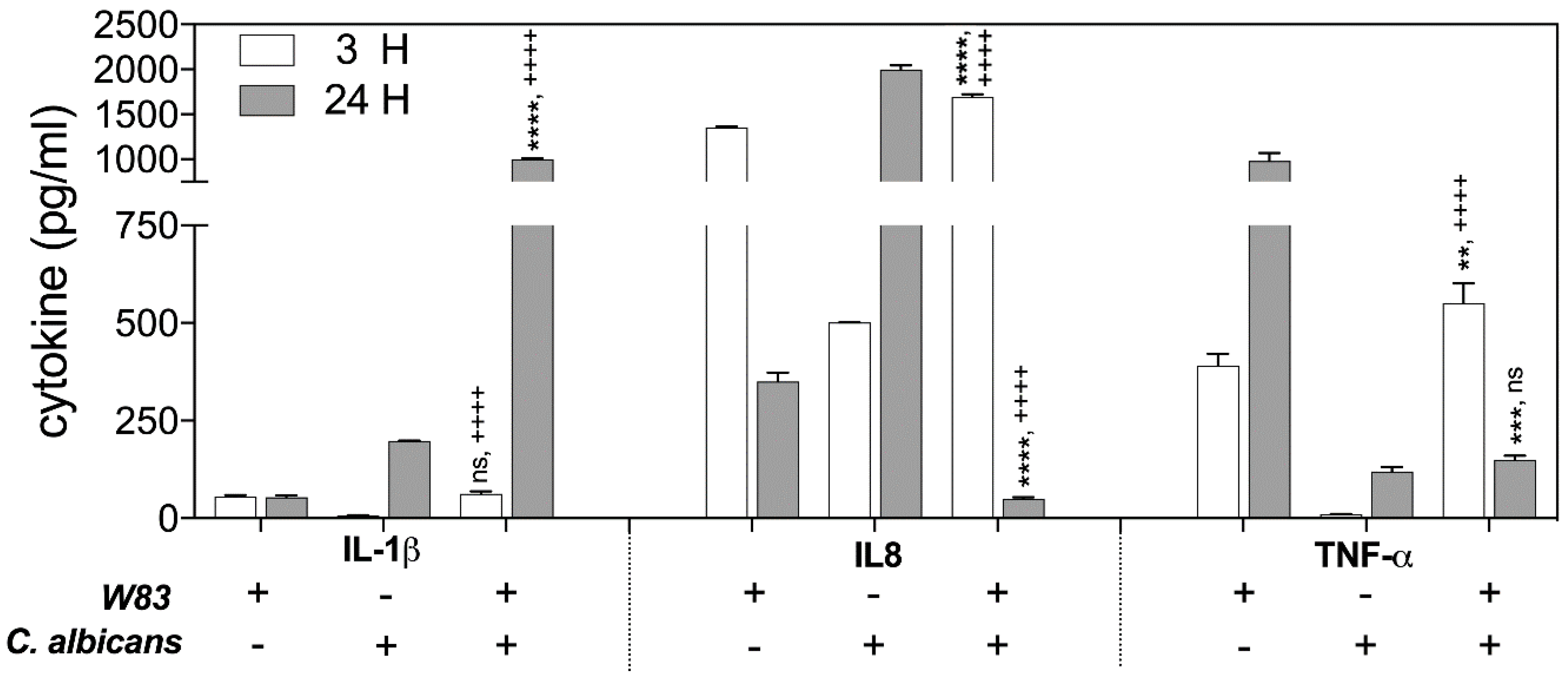
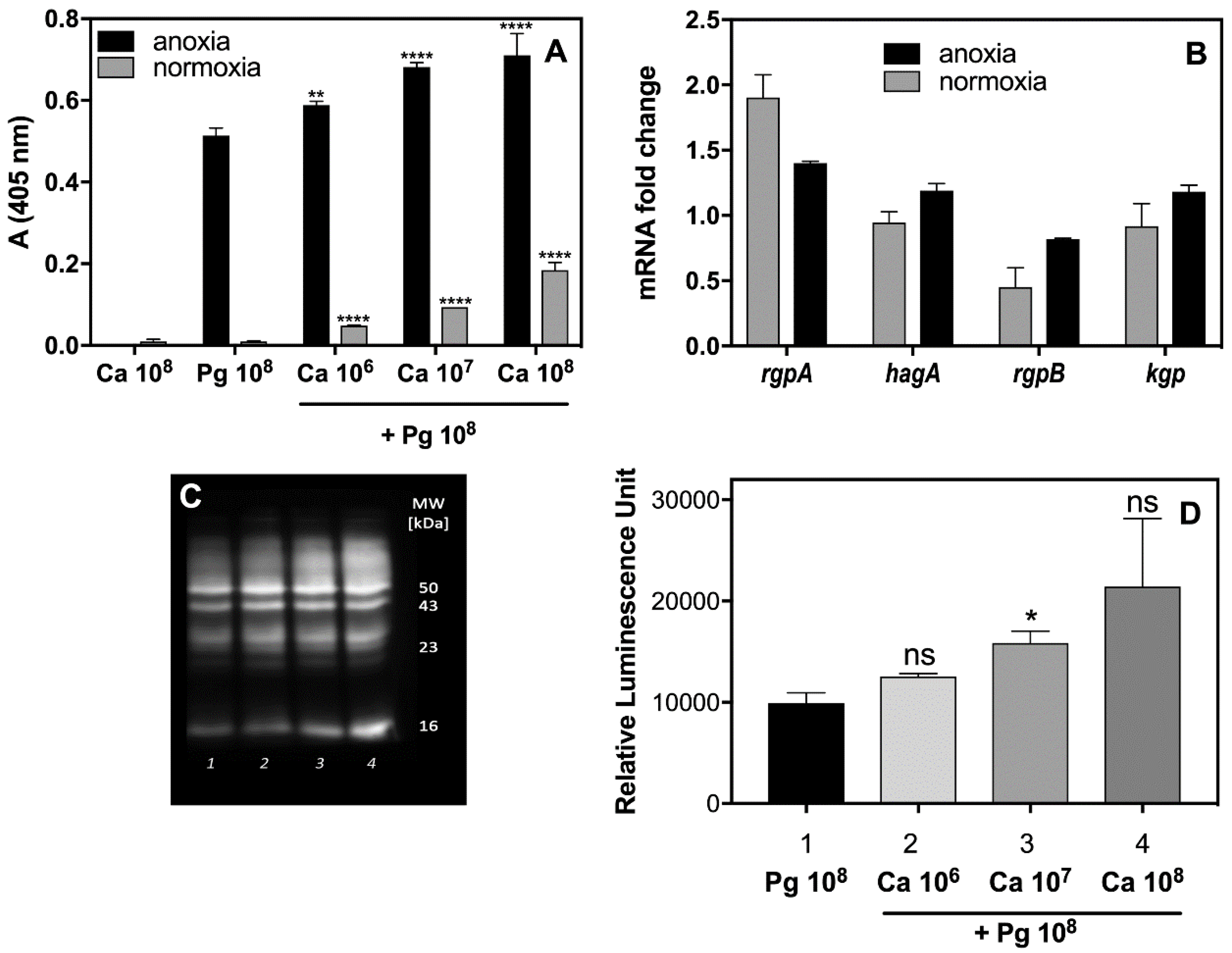
2.1. Collaboration of P. gingivalis Cells with a Fungal Partner during Infection Weakens the Alertness of the Host Defense Response by Macrophages
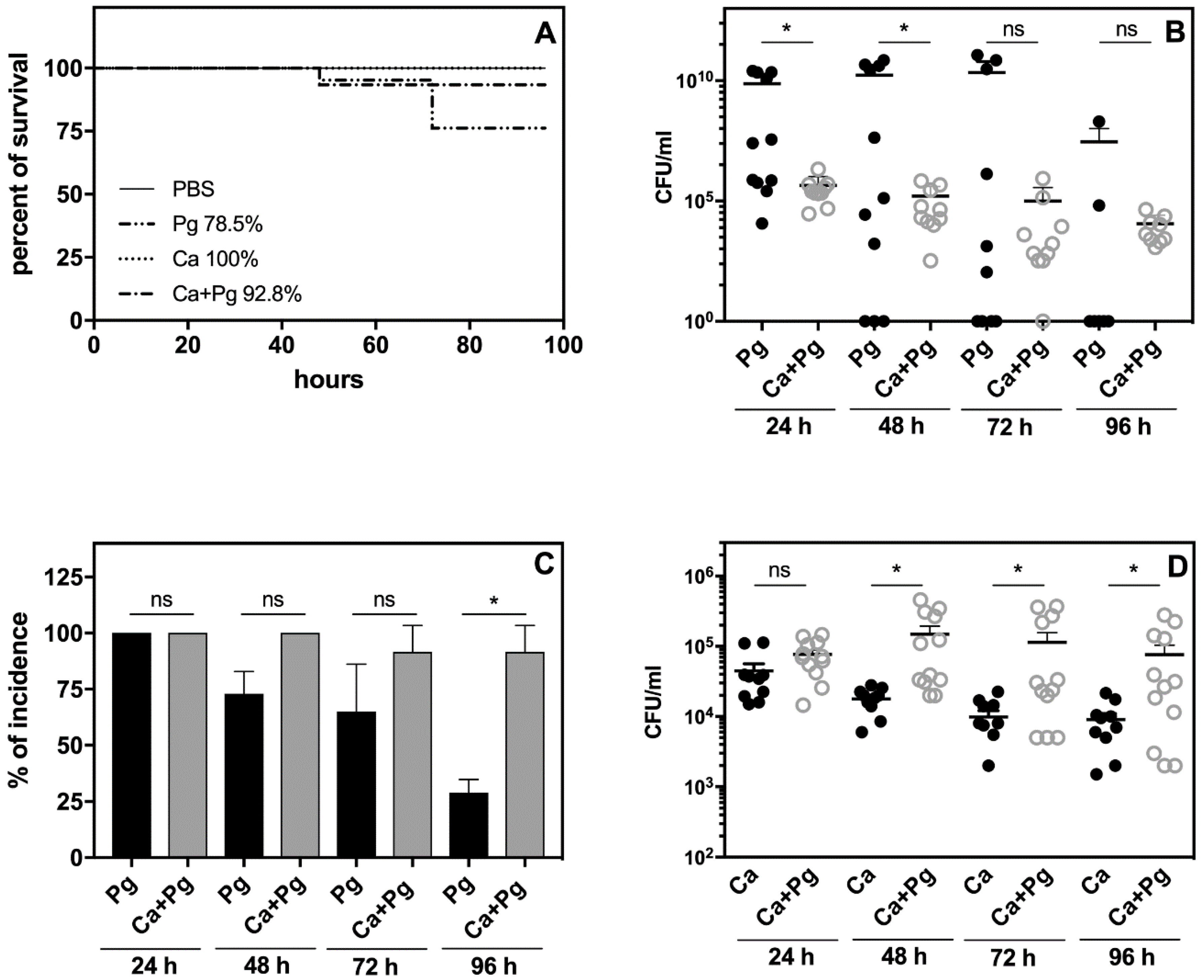
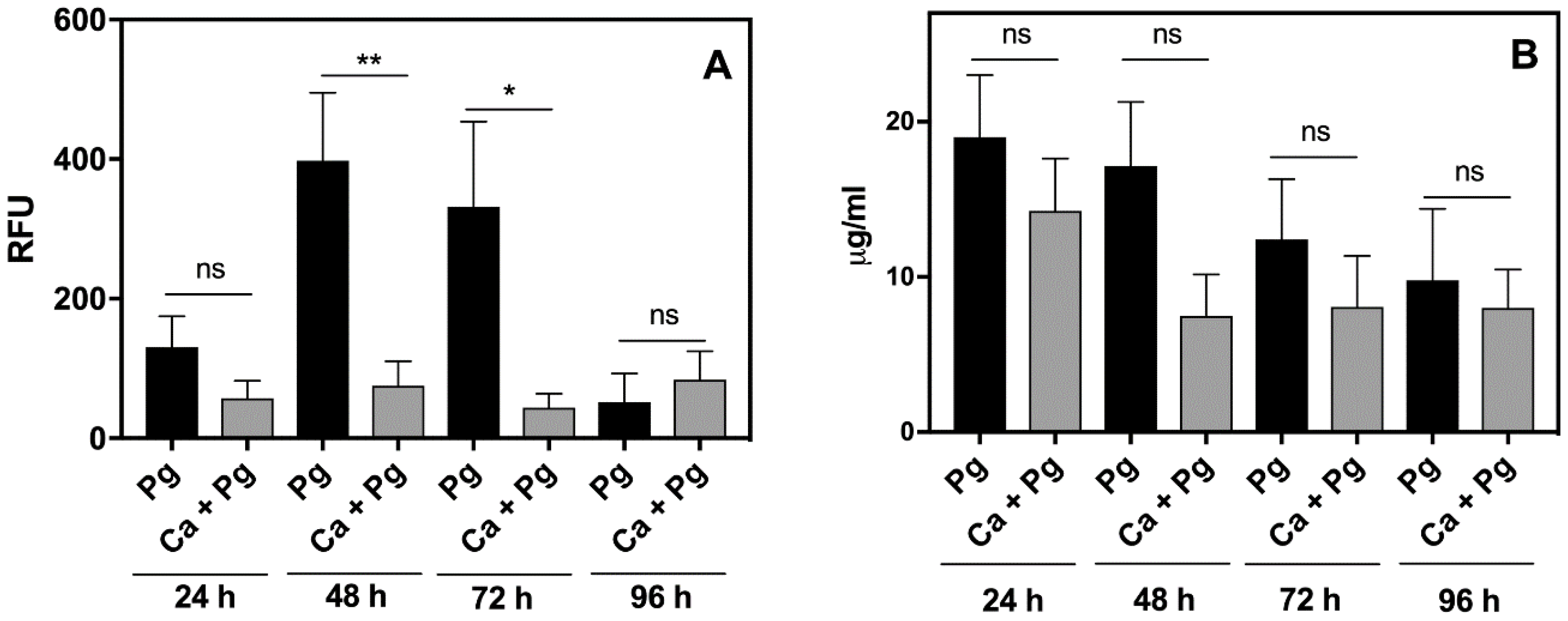
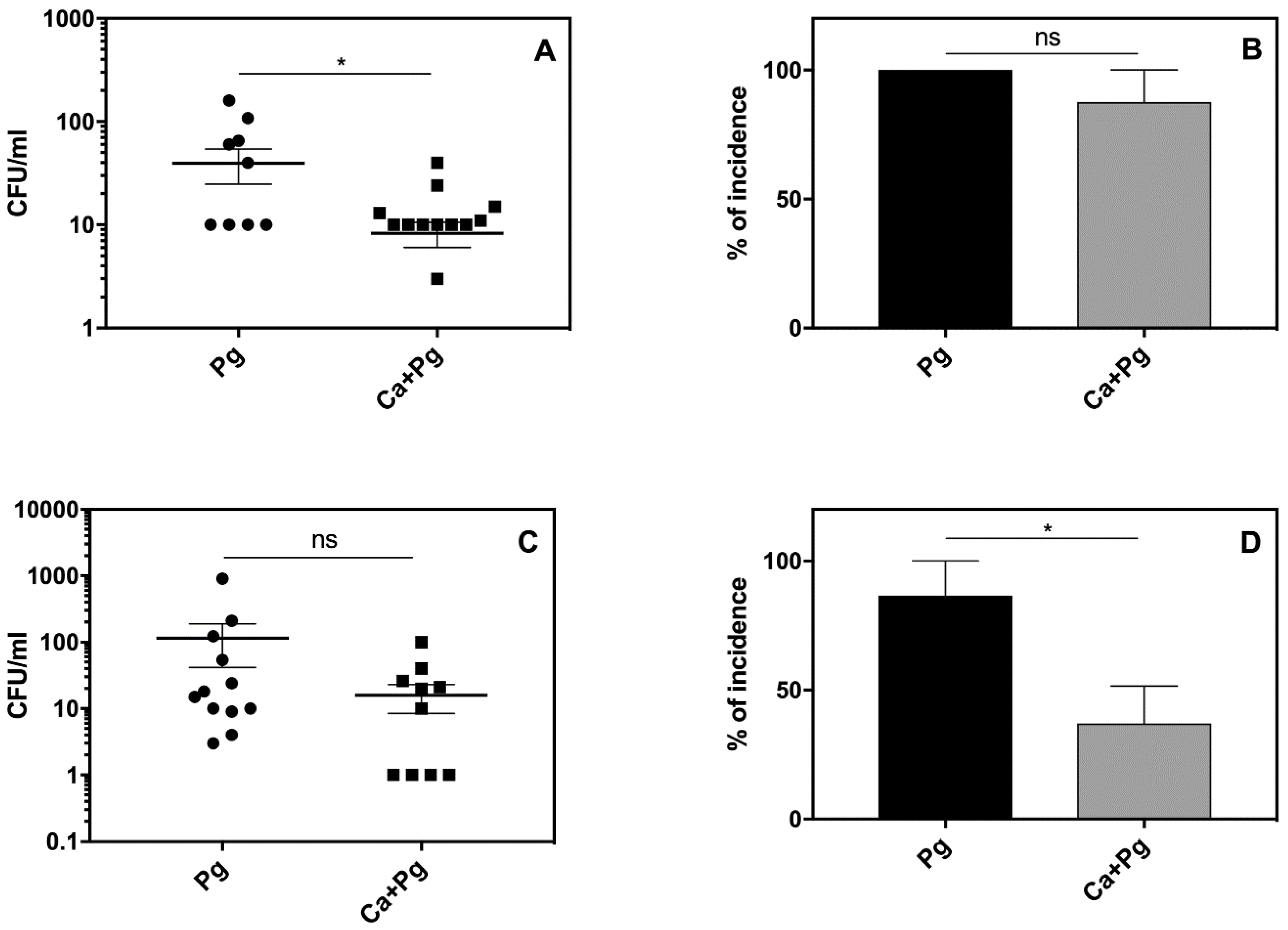
2.2. A Mouse Model of P. gingivalis Host Sensing during Mutual Infection with C. albicans Confirms the Bacterial Protection from Host Recognition by These Fungal Cells
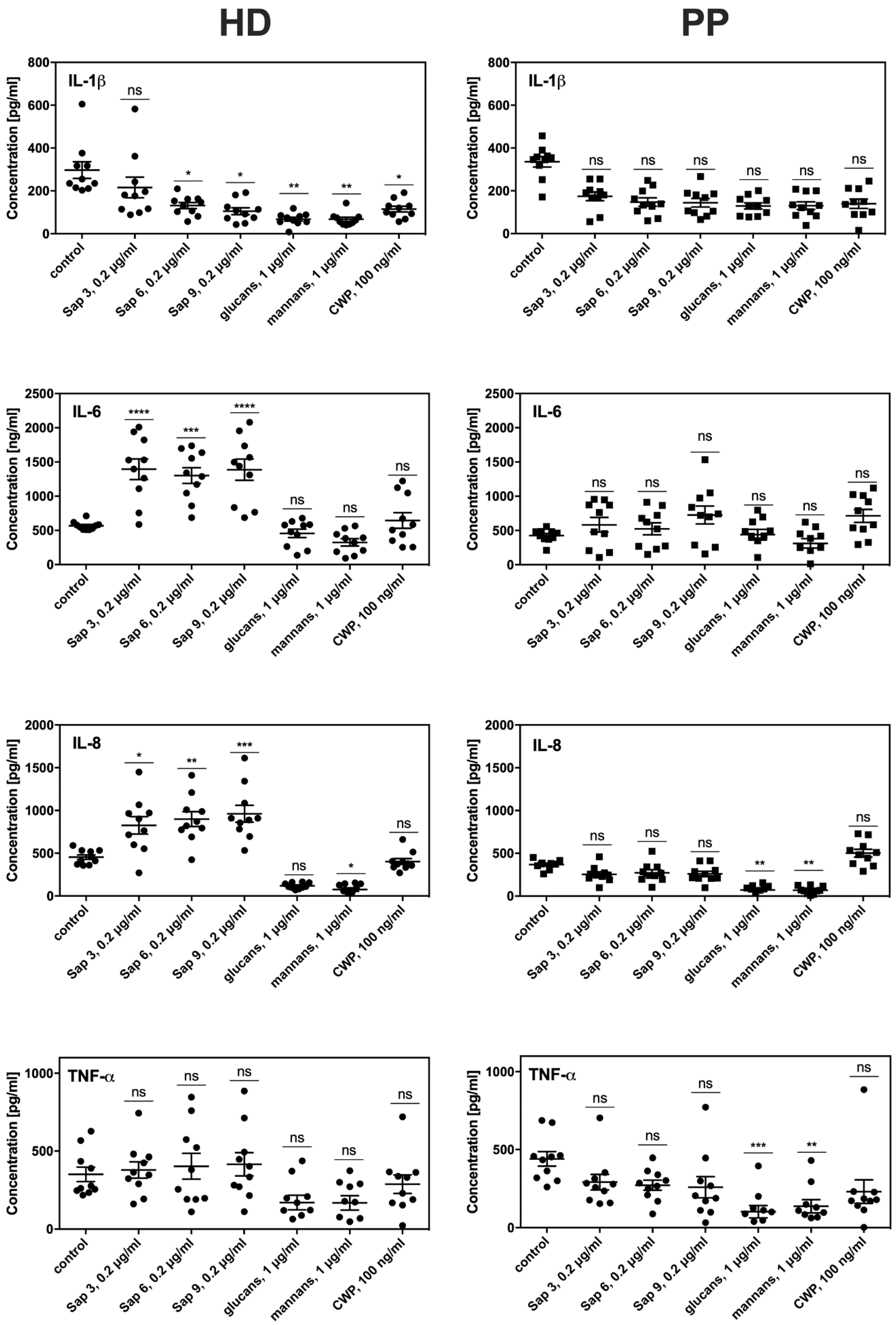
2.3. Altered Susceptibility of Periodontal Patient Fibroblasts to Fungal Infection Compared to Those from Healthy Donors
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Host Cell Cultures
4.2.1. Macrophages
4.2.2. Primary Fibroblasts
4.3. Microbial Cultures
4.4. Preparation of C. albicans Cell Wall Components Including Mannans and Glucans and Cell Wall Proteins
4.5. Recombinant Sap Production
4.6. Mixed-Species Biofilm Formation for Gingipain Detection
4.7. Determination of Gingipain Activity
4.8. Western Blot Analysis
4.9. RNA Isolation, Reverse Transcription, RT-PCR, and RT-qPCR
4.10. Host Cell Infections
4.10.1. THP-1 Co-Culture with Two Microbial Species
4.10.2. Fibroblast Challenge with Virulence Factors of C. albicans Cells
4.11. Cytokine Assay
4.12. Subcutaneous Chamber Model
4.13. Determination of Elastase and Myeloperoxidase Activities
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Als3 | agglutinin-like sequence protein 3 |
| BHI | brain heart infusion |
| CWP | cell wall proteins |
| DMEM | Dulbecco’s modified Eagle medium |
| FBS | fetal bovine serum |
| HagA | hemagglutinin A |
| HD | healthy donors |
| Hwp1 | hyphal cell wall protein 1 |
| Kgp | lysine-specific gingipain |
| LPS | lipopolysaccharide |
| MCP-1 | monocyte chemoattractant protein-1 |
| MPO | myeloperoxidase |
| NE | elastase |
| PMA | phorbol 12-myristate 13-acetate |
| PP | periodontitis patients |
| PPAD | peptidylarginine deiminase |
| RgpA | arginine-specific gingipain A |
| RgpB | arginine-specific gingipain B |
| Sap | secreted aspartic protease |
| THP-1 | monocyte cell line |
| YPD | yeast extract-peptone-dextrose |
References
- Gao, L.; Xu, T.; Huang, G.; Jiang, S.; Gu, Y.; Chen, F. Oral microbiomes: More and more importance in oral cavity and whole body. Protein Cell 2018, 9, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Kriebel, K.; Hieke, C.; Müller-Hilke, B.; Nakata, M.; Kreikemeyer, B. Oral Biofilms from Symbiotic to Pathogenic Interactions and Associated Disease -Connection of Periodontitis and Rheumatic Arthritis by Peptidylarginine Deiminase. Front. Microbiol. 2018, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Nicu, E.A.; Krom, B.P.; Keijser, B.J.F. Acquiring and maintaining a normal oral microbiome: Current perspective. Front. Cell. Infect. Microbiol. 2014, 4, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Hasegawa, M.; Inohara, N. The Role of Oral Pathobionts in Dysbiosis during Periodontitis Development. J. Dent. Res. 2014, 93, 539–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Helmerhorst, E.J.; Leone, C.W.; Troxler, R.F.; Yaskell, T.; Haffajee, A.D.; Socransky, S.S.; Oppenheim, F.G. Identification of early microbial colonizers in human dental biofilm. J. Appl. Microbiol. 2004, 97, 1311–1318. [Google Scholar] [CrossRef]
- Delaney, C.; Kean, R.; Short, B.; Tumelty, M.; McLean, W.; Nile, C.J.; Ramage, G. Fungi at the Scene of the Crime: Innocent Bystanders or Accomplices in Oral Infections? Curr. Clin. Microbiol. Rep. 2018, 5, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Gaffen, S.L.; Herzberg, M.C.; Taubman, M.A.; Van Dyke, T.E. Recent Advances in Host Defense Mechanisms/Therapies against Oral Infectious Diseases and Consequences for Systemic Disease. Adv. Dent. Res. 2014, 26, 30–37. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Kim, J.; Amar, S. Periodontal disease and systemic conditions: A bidirectional relationship. Odontology 2006, 94, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Olsen, I. Systemic Diseases Caused by Oral Infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- AlJehani, Y.A. Risk Factors of Periodontal Disease: Review of the Literature. Int. J. Dent. 2014, 2014, 182513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, B.; Dietrich, T. Causal theory and the etiology of periodontal diseases. Periodontology 2012, 58, 26–36. [Google Scholar] [CrossRef]
- Brown, S.A.; Whiteley, M. A Novel Exclusion Mechanism for Carbon Resource Partitioning in Aggregatibacter actinomycetemcomitans. J. Bacteriol. 2007, 189, 6407–6414. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.M.; Rumbaugh, K.P.; Whiteley, M. Metabolite Cross-Feeding Enhances Virulence in a Model Polymicrobial Infection. PLoS Pathog. 2011, 7, e1002012. [Google Scholar] [CrossRef]
- Wessel, A.K.; Arshad, T.A.; Fitzpatrick, M.; Connell, J.L.; Bonnecaze, R.T.; Shear, J.B.; Whiteley, M. Oxygen limitation within a bacterial aggregate. MBio 2014, 5, e00992. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Kreth, J. The role of hydrogen peroxide in environmental adaptation of oral microbial communities. Oxid. Med. Cell. Longev. 2012, 2012, 717843. [Google Scholar] [CrossRef] [Green Version]
- Hintao, J.; Teanpaisan, R.; Chongsuvivatwong, V.; Ratarasan, C.; Dahlen, G. The microbiological profiles of saliva, supragingival and subgingival plaque and dental caries in adults with and without type 2 diabetes mellitus. Oral Microbiol. Immunol. 2007, 22, 175–181. [Google Scholar] [CrossRef]
- Urzúa, B.; Hermosilla, G.; Gamonal, J.; Morales-Bozo, I.; Canals, M.; Barahona, S.; Cóccola, C.; Cifuentes, V. Yeast diversity in the oral microbiota of subjects with periodontitis: Candida albicans and Candida dubliniensis colonize the periodontal pockets. Med. Mycol. 2008, 46, 783–793. [Google Scholar] [CrossRef]
- Krom, B.P.; Kidwai, S.; Ten Cate, J.M. Candida and other fungal species: Forgotten players of healthy oral microbiota. J. Dent. Res. 2014, 93, 445–451. [Google Scholar] [CrossRef]
- Persoon, I.F.; Buijs, M.J.; Özok, A.R.; Crielaard, W.; Krom, B.P.; Zaura, E.; Brandt, B.W. The mycobiome of root canal infections is correlated to the bacteriome. Clin. Oral Investig. 2017, 21, 1871–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, K.; Chang, C.J.; Hsu, P.C.; Sun, H.S.; Tseng, C.C.; Wang, J.R. Detection of putative periodontal pathogens in non-Insulin-Dependent diabetes mellitus and non-Diabetes mellitus by polymerase chain reaction. J. Periodontal Res. 2001, 36, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Melton, J.J.; Redding, S.W.; Kirkpatrick, W.R.; Reasner, C.A.; Ocampo, G.L.; Venkatesh, A.; Mealey, B.L. Recovery of Candida dubliniensis and other Candida species from the oral cavity of subjects with periodontitis who had well-Controlled and poorly controlled type 2 diabetes: A pilot study. Spec. Care Dent. 2010, 30, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Al Mubarak, S.; Robert, A.A.; Baskaradoss, J.K.; Al-Zoman, K.; Al Sohail, A.; Alsuwyed, A.; Ciancio, S. The prevalence of oral Candida infections in periodontitis patients with type 2 diabetes mellitus. J. Infect. Public Health 2013, 6, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.A.; Wu, J.; Hayes, R.B.; Ahn, J. The oral fungal mycobiome: Characteristics and relation to periodontitis in a pilot study. BMC Microbiol. 2017, 17, 157. [Google Scholar] [CrossRef]
- Canabarro, A.; Valle, C.; Farias, M.R.; Santos, F.B.; Lazera, M.; Wanke, B. Association of subgingival colonization of Candida albicans and other yeasts with severity of chronic periodontitis. J. Periodontal Res. 2013, 48, 428–432. [Google Scholar] [CrossRef]
- Wilson, D.; Naglik, J.R.; Hube, B. The Missing Link between Candida albicans Hyphal Morphogenesis and Host Cell Damage. PLOS Pathog. 2016, 12, e1005867. [Google Scholar] [CrossRef] [Green Version]
- Naglik, J.R.; Richardson, J.P.; Moyes, D.L. Candida albicans Pathogenicity and Epithelial Immunity. PLoS Pathog. 2014, 10, e1004257. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms formed by Candida albicans bloodstream isolates display phenotypic and transcriptional heterogeneity that are associated with resistance and pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, E.P.; Cowley, E.S.; Nobile, C.J.; Hartooni, N.; Newman, D.K.; Johnson, A.D. Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr. Biol. 2014, 24, 2411–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, R.G.; Anil, S.; Samaranayake, L.P. The effect of oral bacteria on Candida albicans germ-Tube formation. APMIS 2001, 109, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Bartnicka, D.; Karkowska-Kuleta, J.; Zawrotniak, M.; Satała, D.; Michalik, K.; Zielinska, G.; Bochenska, O.; Kozik, A.; Ciaston, I.; Koziel, J.; et al. Adhesive protein-mediated cross-Talk between Candida albicans and Porphyromonas gingivalis in dual species biofilm protects the anaerobic bacterium in unfavorable oxic environment. Sci. Rep. 2019, 9, 4376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, I.M.G.; Nobbs, A.H.; Ricomini-Filho, A.P.; Jenkinson, H.F.; Del Bel Cury, A.A. Interkingdom cooperation between Candida albicans, Streptococcus oralis and Actinomyces oris modulates early biofilm development on denture material. Pathog. Dis. 2016, 74, 1–8. [Google Scholar]
- Thein, Z.M.; Samaranayake, Y.H.; Samaranayake, L.P. Effect of oral bacteria on growth and survival of Candida albicans biofilms. Arch. Oral Biol. 2006, 51, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Sztukowska, M.N.; Dutton, L.C.; Delaney, C.; Ramsdale, M.; Ramage, G.; Jenkinson, H.F.; Nobbs, A.H.; Lamont, R.J. Community Development between Porphyromonas gingivalis and Candida albicans Mediated by InlJ and Als3. MBio 2018, 9, e00202-18. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G.; Lamont, R.J. Dancing with the Stars: How Choreographed Bacterial Interactions Dictate Nososymbiocity and Give Rise to Keystone Pathogens, Accessory Pathogens, and Pathobionts. Trends Microbiol. 2016, 24, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Karkowska-Kuleta, J.; Bartnicka, D.; Zawrotniak, M.; Zielinska, G.; Kieronska, A.; Bochenska, O.; Ciaston, I.; Koziel, J.; Potempa, J.; Baster, Z.; et al. The activity of bacterial peptidylarginine deiminase is important during formation of dual-Species biofilm by periodontal pathogen Porphyromonas gingivalis and opportunistic fungus Candida albicans. Pathog. Dis. 2018, 76, fty033. [Google Scholar] [CrossRef]
- Bor, B.; Cen, L.; Agnello, M.; Shi, W.; He, X. Morphological and physiological changes induced by contact-Dependent interaction between Candida albicans and Fusobacterium nucleatum. Sci. Rep. 2016, 6, 27956. [Google Scholar] [CrossRef]
- Dogan, S.; Gunzer, F.; Guenay, H.; Hillmann, G.; Geurtsen, W. Infection of primary human gingival fibroblasts by Porphyromonas gingivalis and Prevotella intermedia. Clin. Oral Investig. 2000, 4, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Krysan, D.J.; Sutterwala, F.S.; Wellington, M. Catching Fire: Candida albicans, Macrophages, and Pyroptosis. PLoS Pathog. 2014, 10, e1004139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, R.S.; O′Brien-Simpson, N.M.; Holden, J.A.; Lenzo, J.C.; Fong, S.B.; Reynolds, E.C. Unprimed, M1 and M2 Macrophages Differentially Interact with Porphyromonas gingivalis. PLoS ONE 2016, 11, e0158629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fokkema, S.J. Peripheral blood monocyte responses in periodontitis. Int. J. Dent. Hyg. 2012, 10, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z. The use of THP-1 cells as a model for mimicking the function and regulation of monocytes and macrophages in the vasculature. Atherosclerosis 2012, 221, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, K.; Khalaf, H.; Bengtsson, T. Gingipains from Porphyromonas gingivalis play a significant role in induction and regulation of CXCL8 in THP-1 cells. BMC Microbiol. 2014, 14, 193. [Google Scholar] [CrossRef] [Green Version]
- Stathopoulou, P.G.; Benakanakere, M.R.; Galicia, J.C.; Kinane, D.F. The host cytokine response to Porphyromonas gingivalis is modified by gingipains. Oral Microbiol. Immunol. 2009, 24, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Genco, C.A.; Cutler, C.W.; Kapczynski, D.; Maloney, K.; Arnold, R.R. A novel mouse model to study the virulence of and host response to Porphyromonas (Bacteroides) gingivalis. Infect. Immun. 1991, 59, 1255–1263. [Google Scholar] [CrossRef] [Green Version]
- Houri-Haddad, Y.; Soskolne, W.A.; Halabi, A.; Barak, V.; Shapira, L. Repeat bacterial challenge in a subcutaneous chamber model results in augmented tumour necrosis factor-alpha and interferon-Gamma response, and suppression of interleukin-10. Immunology 2000, 99, 215–220. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-Species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A.; Ohshima, M.; Sugano, N.; Otsuka, K.; Ito, K. Profiling the cytokines in gingival crevicular fluid using a cytokine antibody array. J. Periodontol. 2006, 77, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Figueredo, C.M.S.; Ribeiro, M.S.M.; Fischer, R.G.; Gustafsson, A. Increased Interleukin-1β Concentration in Gingival Crevicular Fluid as a Characteristic of Periodontitis. J. Periodontol. 1999, 70, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Slade, G.D.; Beck, J.D.; Offenbacher, S. Gingival crevicular fluid interleukin-1beta, prostaglandin E2 and periodontal status in a community population. J. Clin. Periodontol. 2007, 34, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Ozmeriç, N.; Bal, B.; Baloş, K.; Berker, E.; Bulut, S. The correlation of gingival crevicular fluid interleukin-8 levels and periodontal status in localized juvenile periodontitis. J. Periodontol. 1998, 69, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- MacKenzie, A.; Wilson, H.L.; Kiss-Toth, E.; Dower, S.K.; North, R.A.; Surprenant, A. Rapid secretion of interleukin-1beta by microvesicle shedding. Immunity 2001, 15, 825–835. [Google Scholar] [CrossRef]
- Qu, Y.; Franchi, L.; Nunez, G.; Dubyak, G.R. Nonclassical IL-1 beta secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. J. Immunol. 2007, 179, 1913–1925. [Google Scholar] [CrossRef] [Green Version]
- Calkins, C.C.; Platt, K.; Potempa, J.; Travis, J. Inactivation of Tumor Necrosis Factor-α by Proteinases (Gingipains) from the Periodontal Pathogen, Porphyromonas gingivalis. J. Biol. Chem. 1998, 273, 6611–6614. [Google Scholar] [CrossRef] [Green Version]
- Mikolajczyk-Pawlinska, J.; Travis, J.; Potempa, J. Modulation of interleukin-8 activity by gingipains from Porphyromonas gingivalis: Implications for pathogenicity of periodontal disease. FEBS Lett. 1998, 440, 282–286. [Google Scholar] [CrossRef] [Green Version]
- Palm, E.; Khalaf, H.; Bengtsson, T. Porphyromonas gingivalis downregulates the immune response of fibroblasts. BMC Microbiol. 2013, 13, 155. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.I.; Xie, Z.; Sobue, T.; Thompson, A.; Biyikoglu, B.; Ricker, A.; Ikonomou, L.; Dongari-Bagtzoglou, A. Synergistic interaction between Candida albicans and commensal oral streptococci in a novel in vitro mucosal model. Infect. Immun. 2012, 80, 620–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huebener, P.; Abou-Khamis, T.; Zymek, P.; Bujak, M.; Ying, X.; Chatila, K.; Haudek, S.; Thakker, G.; Frangogiannis, N.G. CD44 is critically involved in infarct healing by regulating the inflammatory and fibrotic response. J. Immunol. 2008, 180, 2625–2633. [Google Scholar] [CrossRef] [PubMed]
- Luisa Gil, M.; Murciano, C.; Yáñez, A.; Gozalbo, D. Role of Toll-Like receptors in systemic Candida albicans infections. Front. Biosci. (Landmark Ed.) 2016, 21, 278–302. [Google Scholar] [PubMed] [Green Version]
- Pinheiro, C.R.; Coelho, A.L.; de Oliveira, C.E.; Gasparoto, T.H.; Garlet, G.P.; Silva, J.S.; Santos, C.F.; Cavassani, K.A.; Hogaboam, C.M.; Campanelli, A.P. Recognition of Candida albicans by gingival fibroblasts: The role of TLR2, TLR4/CD14, and MyD88. Cytokine 2018, 106, 67–75. [Google Scholar] [CrossRef]
- Wilensky, A.; Tzach-Nahman, R.; Potempa, J.; Shapira, L.; Nussbaum, G. Porphyromonas gingivalis gingipains selectively reduce CD14 expression, leading to macrophage hyporesponsiveness to bacterial infection. J. Innate Immun. 2015, 7, 127–135. [Google Scholar] [CrossRef]
- Desta, T.; Graves, D.T. Fibroblast apoptosis induced by Porphyromonas gingivalis is stimulated by a gingipain and caspase-Independent pathway that involves apoptosis-Inducing factor. Cell. Microbiol. 2007, 9, 2667–2675. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.-C.; Chang, L.-C.; Chen, T.-C.; Lee, K.-C.; Lee, K.-F.; Chen, C.-N.; Yu, H.-R. Sterol Regulatory Element-Binding Protein-1c Regulates Inflammasome Activation in Gingival Fibroblasts Infected with High-Glucose-Treated Porphyromonas gingivalis. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef]
- Maksylewicz, A.; Bysiek, A.; Lagosz, K.B.; Macina, J.M.; Kantorowicz, M.; Bereta, G.; Sochalska, M.; Gawron, K.; Chomyszyn-Gajewska, M.; Potempa, J.; et al. BET Bromodomain Inhibitors Suppress Inflammatory Activation of Gingival Fibroblasts and Epithelial Cells From Periodontitis Patients. Front. Immunol. 2019, 10, 933. [Google Scholar] [CrossRef] [Green Version]
- Gawron, K.; Łazarz-Bartyzel, K.; Kowalska, A.; Bereta, G.; Nowakowska, Z.; Plakwicz, P.; Potempa, J.; Fertala, A.; Chomyszyn-Gajewska, M. Fibroblasts from recurrent fibrotic overgrowths reveal high rate of proliferation in vitro-Findings from the study of hereditary and idiopathic gingival fibromatosis. Connect. Tissue Res. 2019, 60, 29–39. [Google Scholar] [CrossRef]
- Kocourek, J.; Ballou, C.E. Method for fingerprinting yeast cell wall mannans. J. Bacteriol. 1969, 100, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.-I.; Lee, Y.C. Carbohydrate analysis by a phenol-Sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Zawrotniak, M.; Bochenska, O.; Karkowska-Kuleta, J.; Seweryn-Ozog, K.; Aoki, W.; Ueda, M.; Kozik, A.; Rapala-Kozik, M. Aspartic Proteases and Major Cell Wall Components in Candida albicans Trigger the Release of Neutrophil Extracellular Traps. Front. Cell. Infect. Microbiol. 2017, 7, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, W.; Kitahara, N.; Miura, N.; Morisaka, H.; Yamamoto, Y.; Kuroda, K.; Ueda, M. Comprehensive characterization of secreted aspartic proteases encoded by a virulence gene family in Candida albicans. J. Biochem. 2011, 150, 431–438. [Google Scholar] [CrossRef]
- Rapala-Kozik, M.; Bochenska, O.; Zawrotniak, M.; Wolak, N.; Trebacz, G.; Gogol, M.; Ostrowska, D.; Aoki, W.; Ueda, M.; Kozik, A. Inactivation of the antifungal and immunomodulatory properties of human cathelicidin LL-37 by aspartic proteases produced by the pathogenic yeast Candida albicans. Infect. Immun. 2015, 83, 2518–2530. [Google Scholar] [CrossRef] [Green Version]
- Potempa, J.; Mikolajczyk-Pawlinska, J.; Brassell, D.; Nelson, D.; Thøgersen, I.B.; Enghild, J.J.; Travis, J. Comparative Properties of Two Cysteine Proteinases (Gingipains R), the Products of Two Related but Individual Genes of Porphyromonas gingivalis. J. Biol. Chem. 1998, 273, 21648–21657. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. The single-Step method of RNA isolation by acid guanidinium thiocyanate–Phenol–Chloroform extraction: Twenty-Something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence |
|---|---|
| 16S rRNA For | TGTAGATGACTGATGGTGAAAACC |
| 16S rRNA Rev | ACGTCATCCCCACCTTCCTC |
| rgpA For | TGGACAGGTTGTAAACTTTGCGCC |
| rgpA Rev | TTGCCTTGTTCCGAAGTTTCGCTC |
| kgp For | GCTCAGTACATCCTGCAGAAGTTC |
| kgp Rev | CTATAAGAAGCCTGATTCTGAGGC |
| hagA For | TGATGACGTGGCTGTTTCTGGTGA |
| hagA Rev | TTGTACTGGCCGGGAGCTACATTT |
| rgpB For | AAGATATCTATAAGAGCGTCTTCA |
| rgpB Rev | CGACCGATGAAGACTTCG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartnicka, D.; Gonzalez-Gonzalez, M.; Sykut, J.; Koziel, J.; Ciaston, I.; Adamowicz, K.; Bras, G.; Zawrotniak, M.; Karkowska-Kuleta, J.; Satala, D.; et al. Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue. Int. J. Mol. Sci. 2020, 21, 1984. https://doi.org/10.3390/ijms21061984
Bartnicka D, Gonzalez-Gonzalez M, Sykut J, Koziel J, Ciaston I, Adamowicz K, Bras G, Zawrotniak M, Karkowska-Kuleta J, Satala D, et al. Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue. International Journal of Molecular Sciences. 2020; 21(6):1984. https://doi.org/10.3390/ijms21061984
Chicago/Turabian StyleBartnicka, Dominika, Miriam Gonzalez-Gonzalez, Joanna Sykut, Joanna Koziel, Izabela Ciaston, Karina Adamowicz, Grazyna Bras, Marcin Zawrotniak, Justyna Karkowska-Kuleta, Dorota Satala, and et al. 2020. "Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue" International Journal of Molecular Sciences 21, no. 6: 1984. https://doi.org/10.3390/ijms21061984
APA StyleBartnicka, D., Gonzalez-Gonzalez, M., Sykut, J., Koziel, J., Ciaston, I., Adamowicz, K., Bras, G., Zawrotniak, M., Karkowska-Kuleta, J., Satala, D., Kozik, A., Zyla, E., Gawron, K., Lazarz-Bartyzel, K., Chomyszyn-Gajewska, M., & Rapala-Kozik, M. (2020). Candida albicans Shields the Periodontal Killer Porphyromonas gingivalis from Recognition by the Host Immune System and Supports the Bacterial Infection of Gingival Tissue. International Journal of Molecular Sciences, 21(6), 1984. https://doi.org/10.3390/ijms21061984