Proteomics Analysis Reveals that Warburg Effect along with Modification in Lipid Metabolism Improves In Vitro Embryo Development under Low Oxygen

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Evaluation of the Effect of Oxygen Tension (5% vs. 20%) on Morphological Parameters of Embryo Development
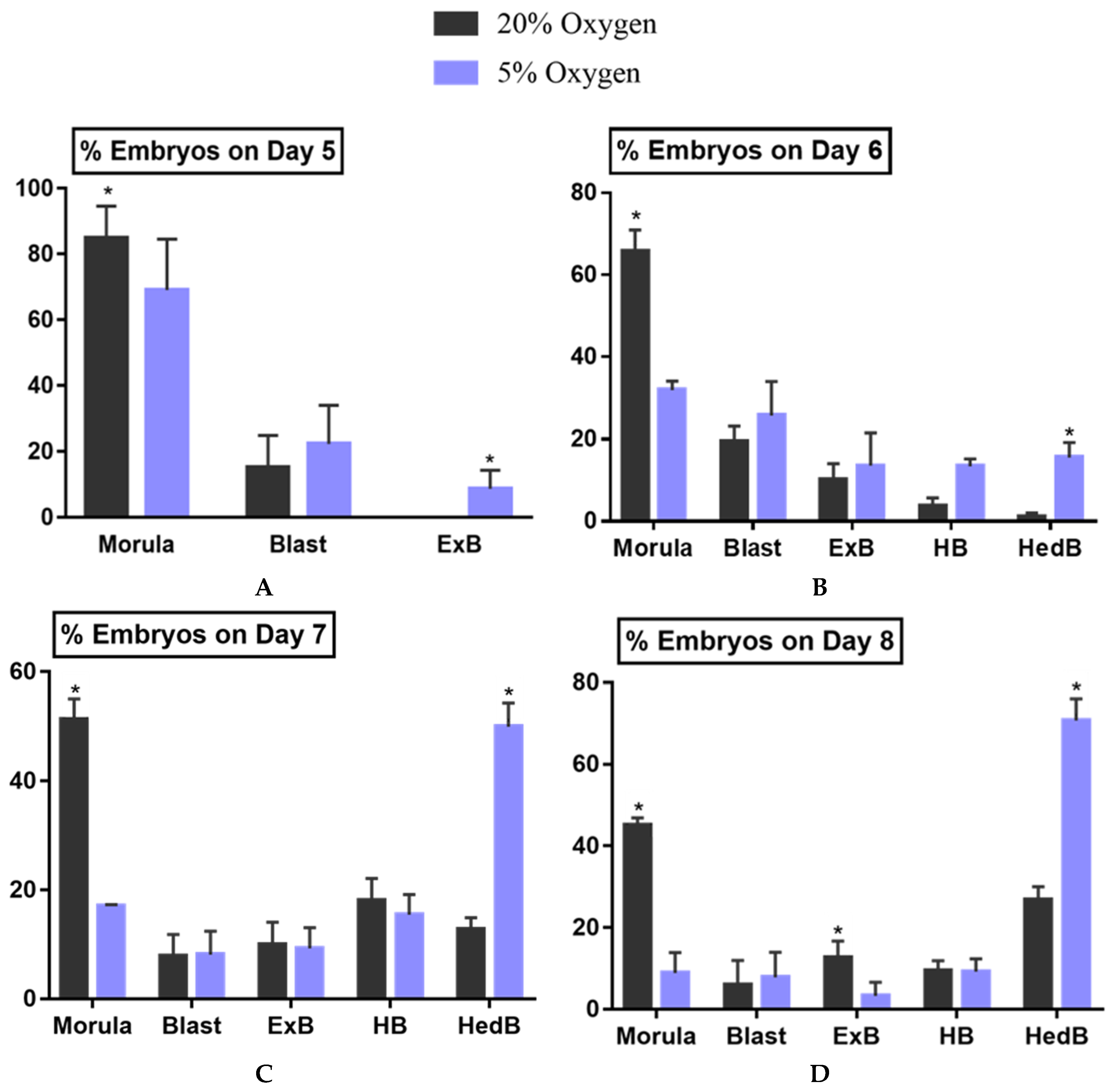
2.1.1. Cleavage, Blastocyst, Hatching Rate and Developmental Kinetics
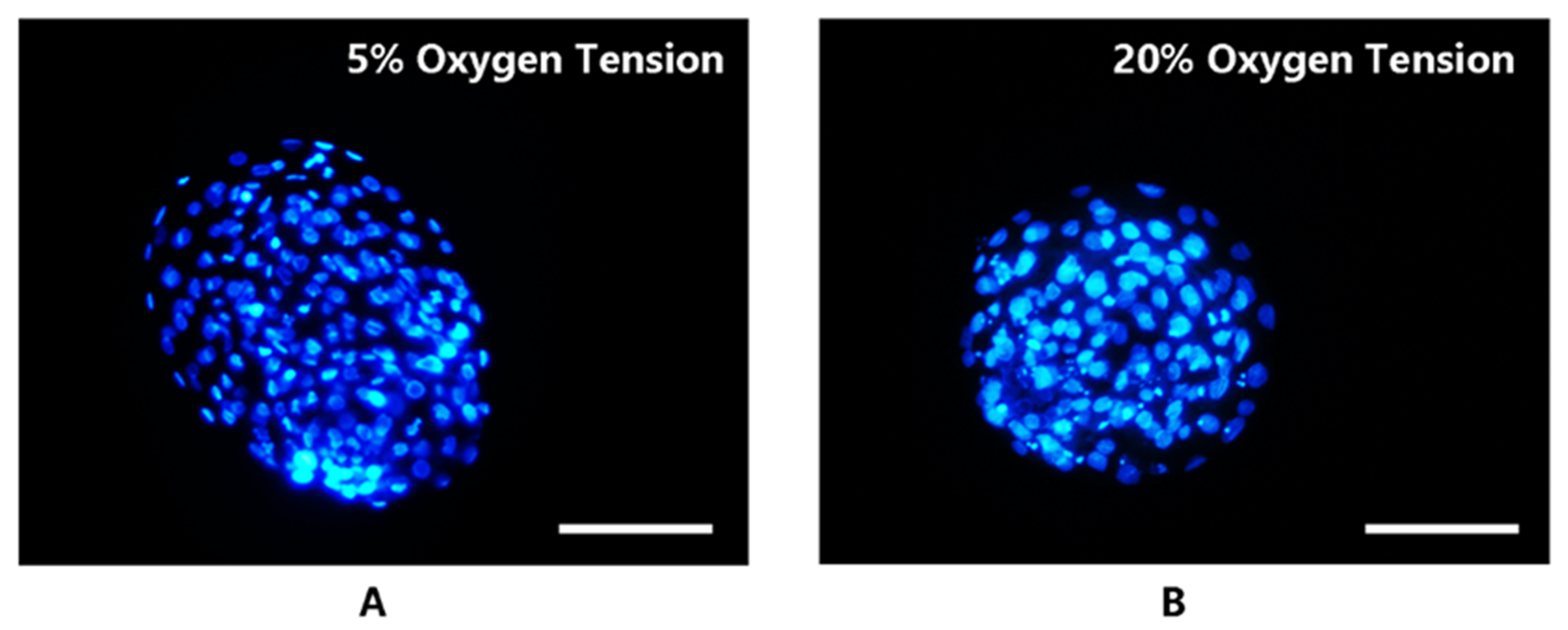
2.1.2. Cell Count
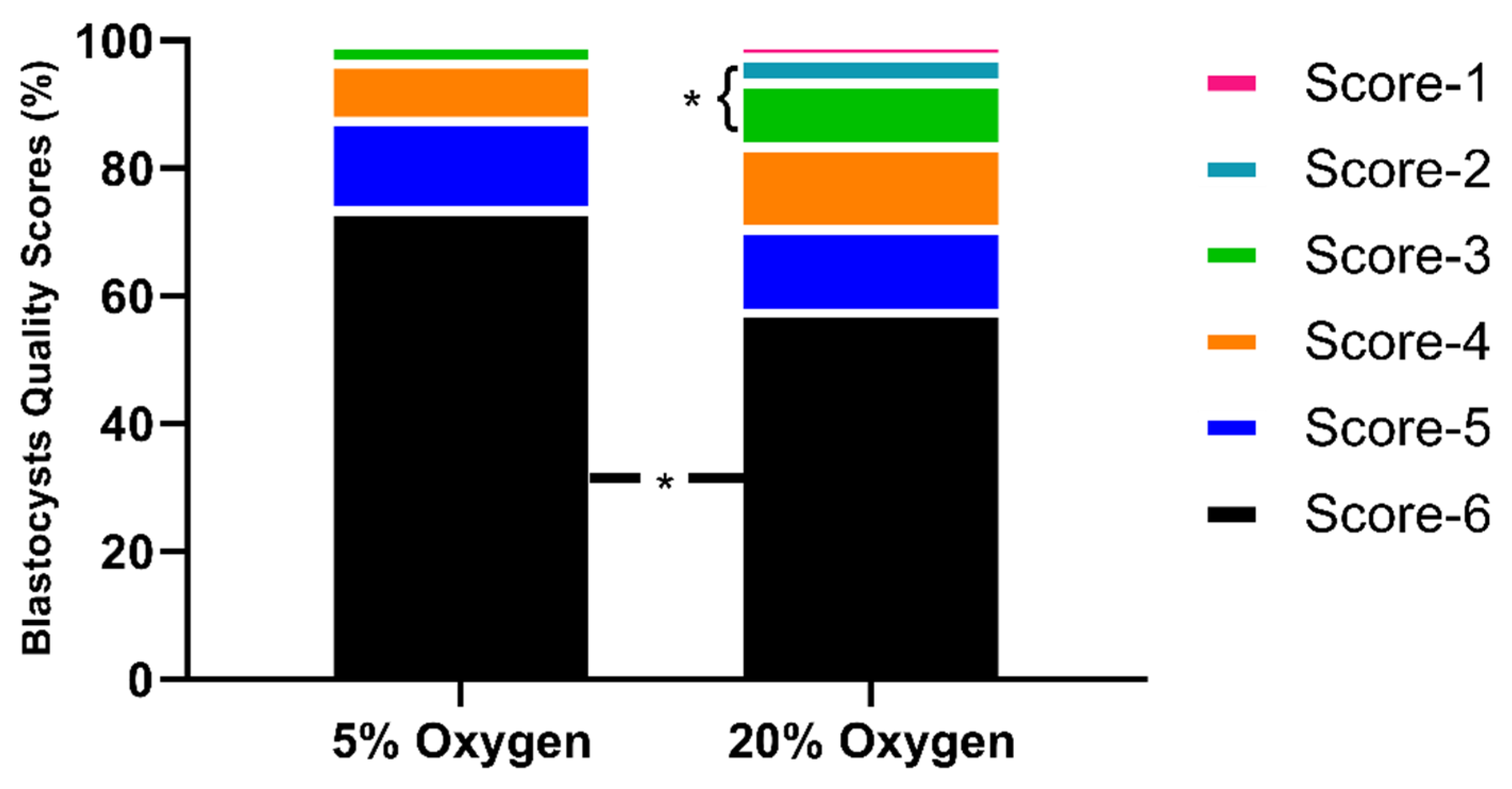
2.1.3. Embryo Quality Scoring
2.1.4. Cryo-Survival Potential of Vitrified Blastocysts
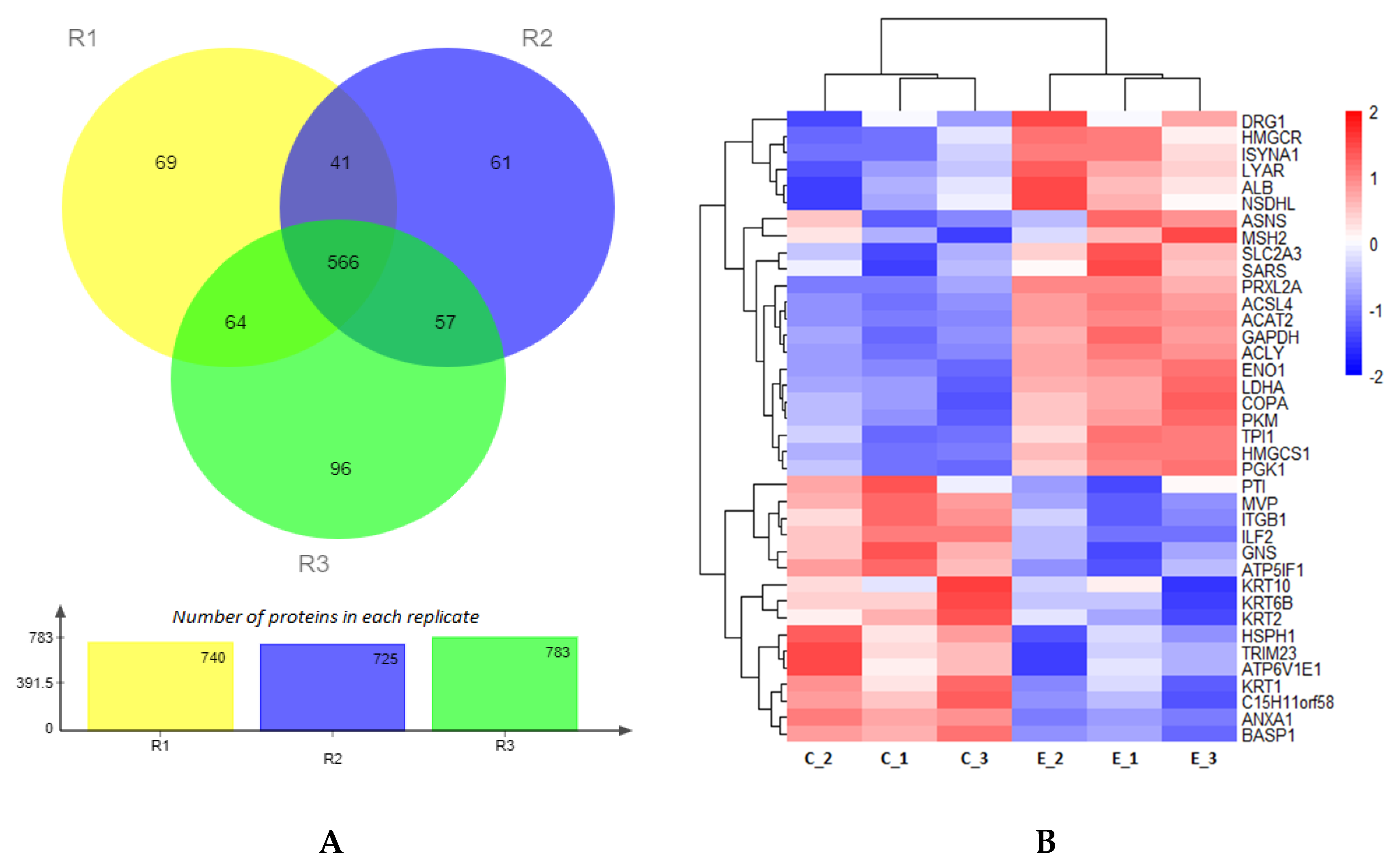
2.2. Proteomic Analysis of Embryos Cultured under 5% or 20% Oxygen Tension
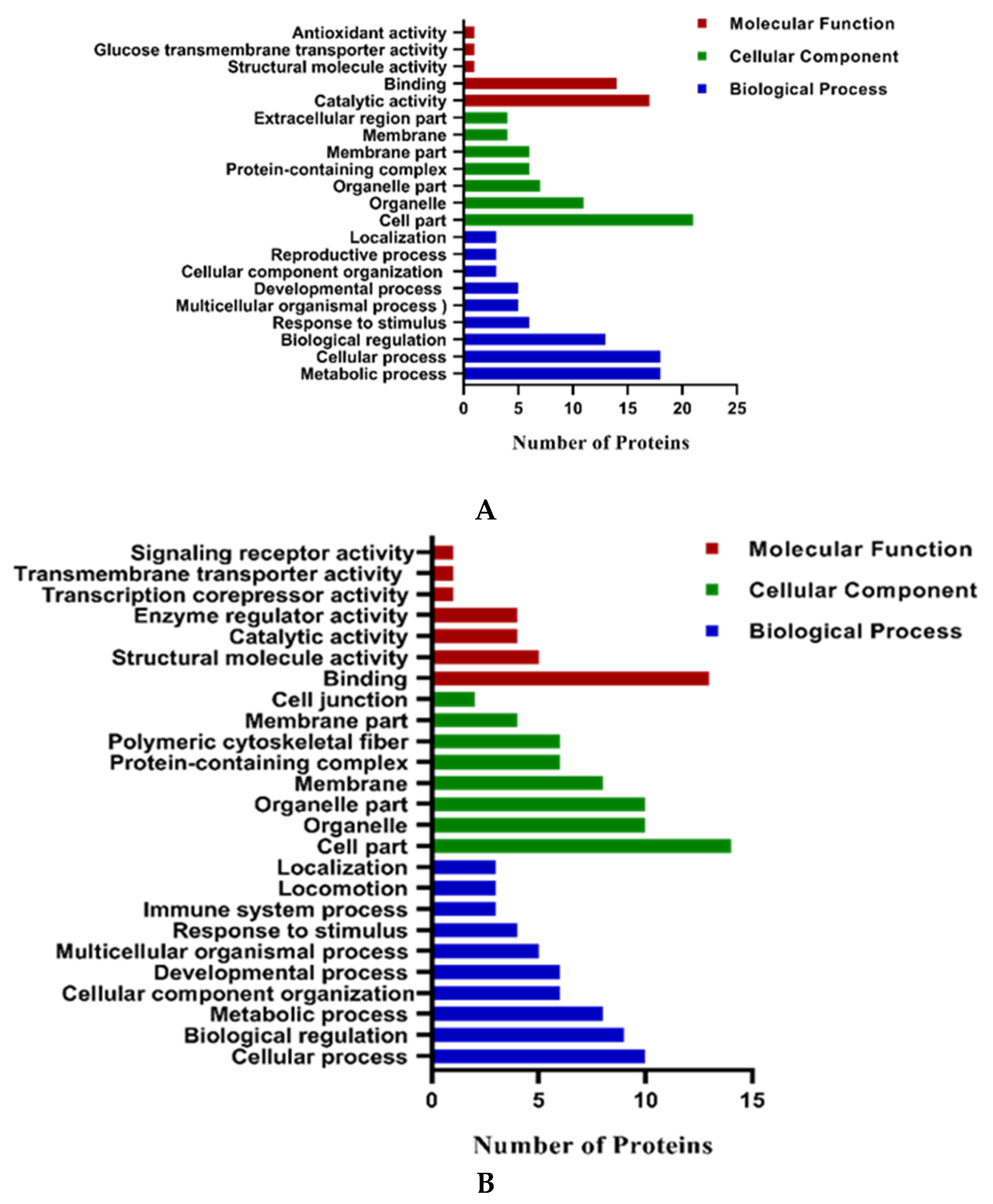
2.2.1. Gene Ontology Analysis of Differentially Expressed Proteins
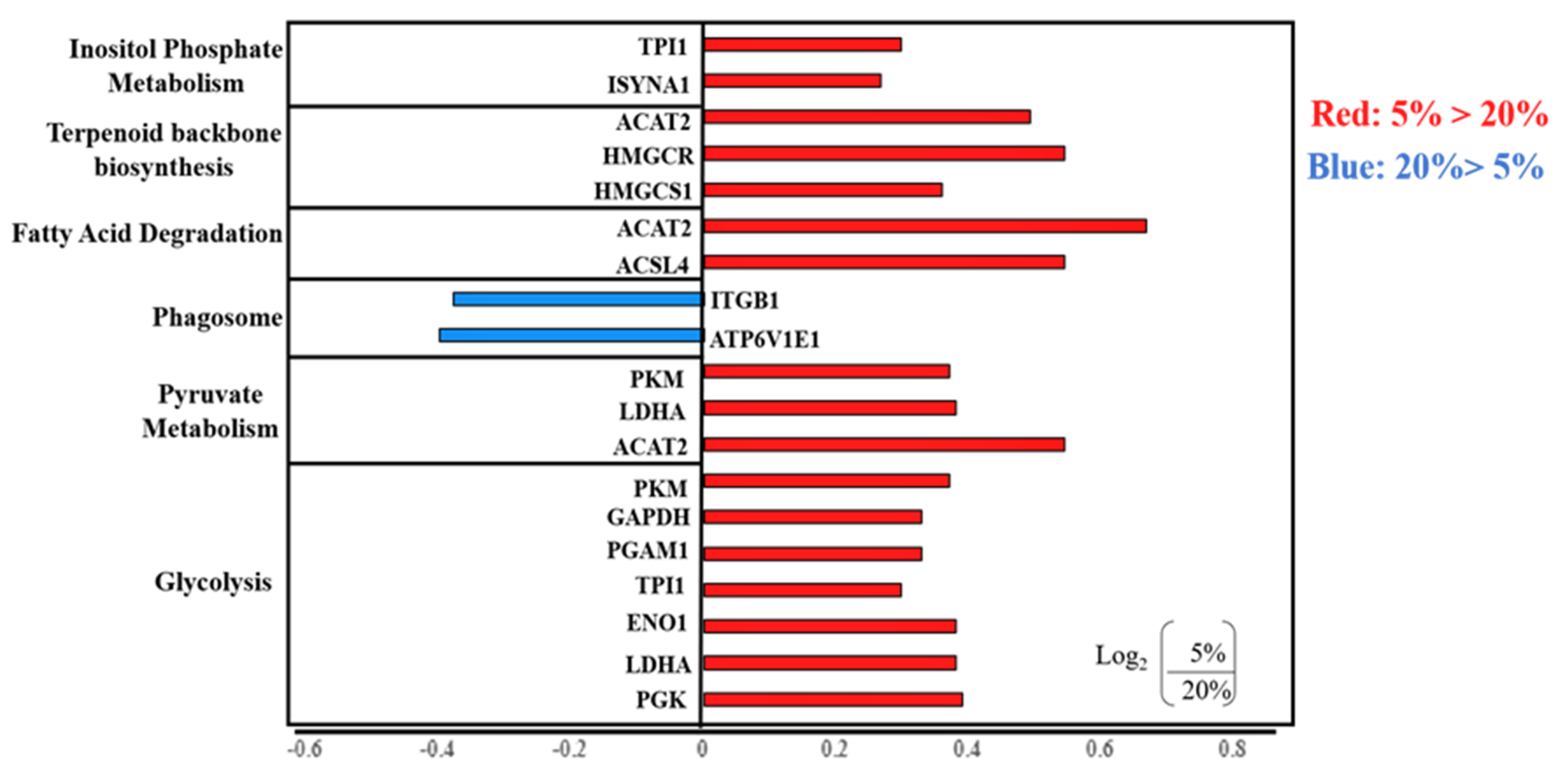
2.2.2. KEGG Analysis of Differentially Expressed Proteins
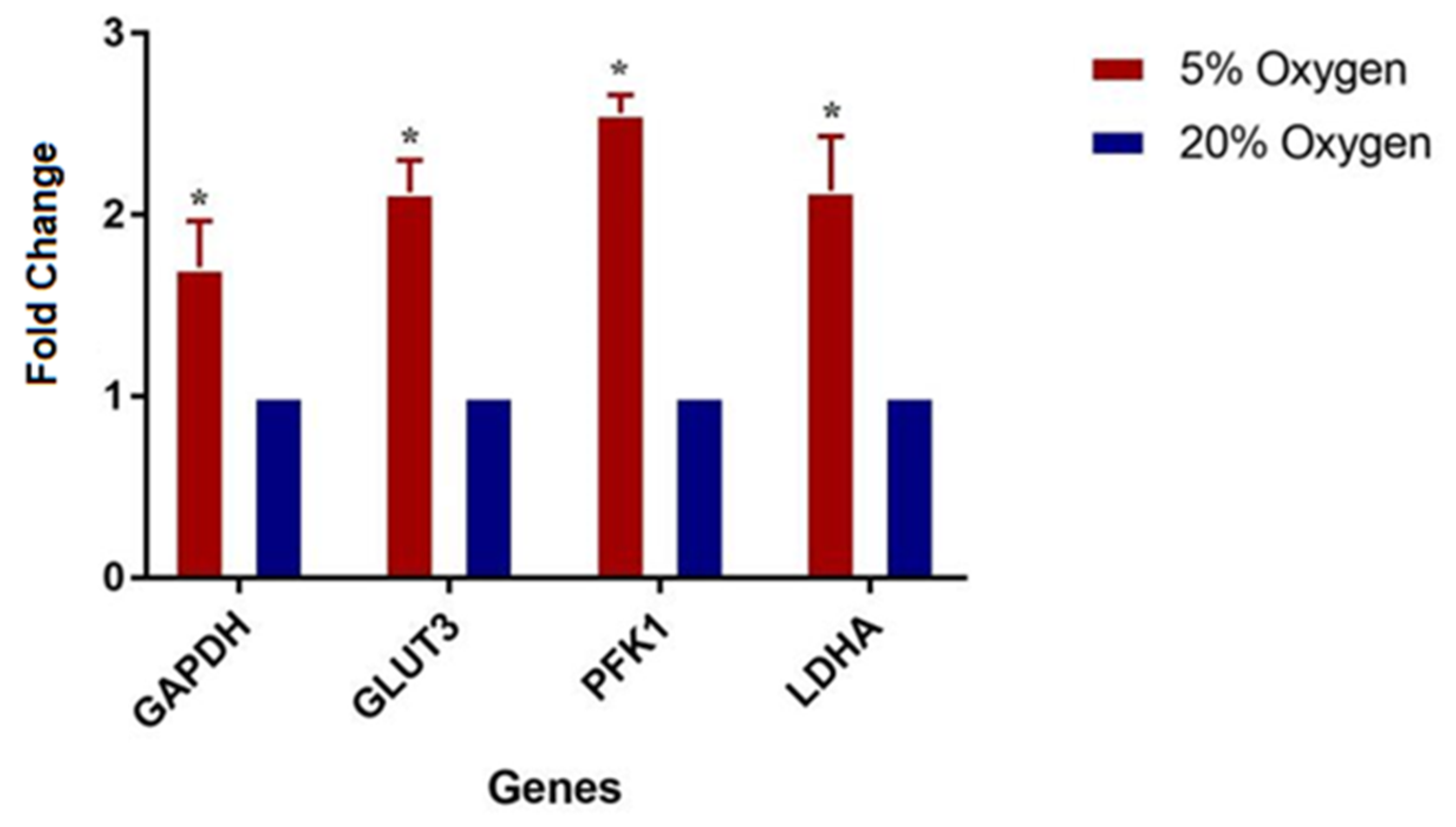
2.2.3. Proteome Profile Validation by Real Time Quantitative PCR
2.2.4. Fluorescent intensity of Lipids
3. Discussion
3.1. Physiological Oxygen Improves Morphological Parameters of Embryo Development
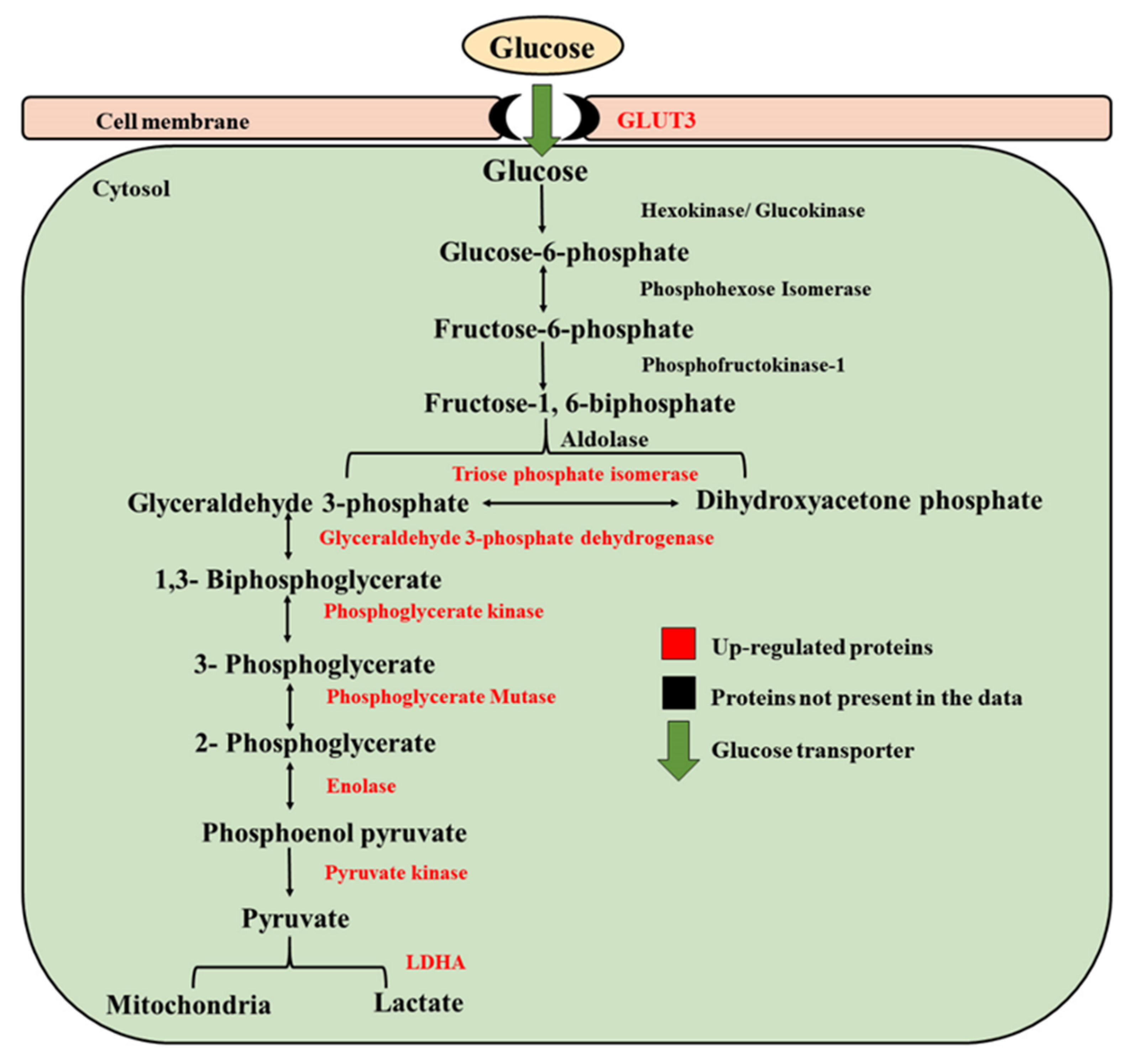
3.2. Embryos Developed under Low Oxygen Showed Pronounced Warburg Effect
3.3. Modification in the Lipid Composition might be Associated with Increased Cryo-Survival Ability of Embryos Cultured under Low Oxygen
4. Materials and Methods
4.1. Morphological Parameters of Embryo Development under Different Oxygen Tension (5% vs. 20%)
4.1.1. In Vitro Oocyte Maturation, Fertilization and Culture
4.1.2. Cell Staining
4.1.3. Embryo Quality Scoring
4.1.4. Vitrification, Thawing and Hatching Rate of Blastocysts
4.1.5. Statistical Analysis
4.2. Proteomic Analysis of Embryos Cultured under 5% or 20% Oxygen Tension
4.2.1. Cell Lysis
4.2.2. Protein Purification
4.2.3. Peptide Recovery and Preparation
4.2.4. iTRAQ Labelling
4.2.5. Mass Spectrometry
4.2.6. Database Search Settings
4.2.7. Bioinformatics Analysis
4.2.8. Real Time qPCR Analysis
4.2.9. Assessment of Lipid Accumulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). MILK and Dairy Products in Human Nutrition; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Kussano, N.R.; Leme, L.O.; Guimaraes, A.L.; Franco, M.M.; Dode, M.A. Molecular markers for oocyte competence in bovine cumulus cells. Theriogenology 2016, 85, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Besenfelder, U.; Havlicek, V.; Kuzmany, A.; Brem, G. Endoscopic approaches to manage In Vitro and in vivo embryo development: Use of the bovine oviduct. Theriogenology 2010, 73, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chaubal, S.A.; Du, F. Optimizing IVF with sexed sperm in cattle. Theriogenology 2009, 71, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, B.; Boccia, L.; Marchandise, J.; Di Palo, R.; George, F.; Donnay, I.; Zicarelli, L. Enrichment of in vitro maturation medium for buffalo (Bubalus bubalis) oocytes with thiol compounds: Effects of cystine on glutathione synthesis and embryo development. Theriogenology 2006, 65, 275–287. [Google Scholar] [CrossRef]
- Wale, P.L.; Gardner, D.K. The effects of chemical and physical factors on mammalian embryo culture and their importance for the practice of assisted human reproduction. Hum. Reprod. Update 2016, 22, 2–22. [Google Scholar] [CrossRef] [Green Version]
- Ajduk, A.; Zernicka-Goetz, M. Quality control of embryo development. Mol. Asp. Med. 2013, 34, 903–918. [Google Scholar] [CrossRef]
- Grupen, C.G. The evolution of porcine embryo In Vitro production. Theriogenology 2014, 81, 24–37. [Google Scholar] [CrossRef]
- Jones, H.W., Jr.; Oehninger, S.; Bocca, S.; Stadtmauer, L.; Mayer, J. Reproductive efficiency of human oocytes fertilized In Vitro. Facts Views Vis. ObGyn 2010, 2, 169–171. [Google Scholar]
- Fischer, B.; Bavister, B.D. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J. Reprod. Fertil. 1993, 99, 673–679. [Google Scholar] [CrossRef]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Harvey, A.J. The role of oxygen in ruminant preimplantation embryo development and metabolism. Anim. Reprod. Sci. 2007, 98, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Elamaran, G.; Singh, K.P.; Singh, M.K.; Singla, S.K.; Chauhan, M.S.; Manik, R.S.; Palta, P. Oxygen concentration and cysteamine supplementation during In Vitro production of buffalo (Bubalus bubalis) embryos affect mRNA expression of BCL-2, BCL-XL, MCL-1, BAX and BID. Reprod. Domest. Anim. 2012, 47, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Betterbed, B.; Wright, R.W., Jr. Development of one-cell ovine embryos in two culture media under two gas atmospheres. Theriogenology 1985, 23, 547–553. [Google Scholar] [CrossRef]
- Khurana, N.K.; Niemann, H. Effects of oocyte quality, oxygen tension, embryo density, cumulus cells and energy substrates on cleavage and morula/blastocyst formation of bovine embryos. Theriogenology 2000, 54, 741–756. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, Y.; Ding, C.; Khoudja, R.Y.; Lin, M.; Awonuga, A.O.; Dai, J.; Puscheck, E.E.; Rappolee, D.A.; Zhou, C. Comparison of 2, 5, and 20% O2 on the development of post-thaw human embryos. J. Assist. Reprod. Genet. 2016, 33, 919–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karja, N.W.; Wongsrikeao, P.; Murakami, M.; Agung, B.; Fahrudin, M.; Nagai, T.; Otoi, T. Effects of oxygen tension on the development and quality of porcine in vitro fertilized embryos. Theriogenology 2004, 62, 1585–1595. [Google Scholar] [CrossRef]
- Bernardi, M.L.; Flechon, J.E.; Delouis, C. Influence of culture system and oxygen tension on the development of ovine zygotes matured and fertilized In Vitro. J. Reprod. Fertil. 1996, 106, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Dahl, J.A.; Jung, I.; Aanes, H.; Greggains, G.D.; Manaf, A.; Lerdrup, M.; Li, G.; Kuan, S.; Li, B.; Lee, A.Y. Broad histone H3K4me3 domains in mouse oocytes modulate maternal-to-zygotic transition. Nature 2016, 537, 548. [Google Scholar] [CrossRef]
- Flyamer, I.M.; Gassler, J.; Imakaev, M.; Brandão, H.B.; Ulianov, S.V.; Abdennur, N.; Razin, S.V.; Mirny, L.A.; Tachibana-Konwalski, K. Single-nucleus Hi-C reveals unique chromatin reorganization at oocyte-to-zygote transition. Nature 2017, 544, 110. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhu, P.; Wu, X.; Li, X.; Wen, L.; Tang, F. Single-Cell methylome landscapes of mouse embryonic stem cells and early embryos analyzed using reduced representation bisulfite sequencing. Genome Res. 2013, 23, 2126–2135. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Liu, Y.; Inoue, A.; Suzuki, T.; Zhao, K.; Zhang, Y. Establishing chromatin regulatory landscape during mouse preimplantation development. Cell 2016, 165, 1375–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Huang, B.; Chen, H.; Yin, Q.; Liu, Y.; Xiang, Y.; Zhang, B.; Liu, B.; Wang, Q.; Xia, W. The landscape of accessible chromatin in mammalian preimplantation embryos. Nature 2016, 534, 652. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Huang, K.; Cai, C.; Cai, L.; Jiang, C.-Y.; Feng, Y.; Liu, Z.; Zeng, Q.; Cheng, L.; Sun, Y.E. Genetic programs in human and mouse early embryos revealed by single-cell RNA sequencing. Nature 2013, 500, 593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zheng, H.; Huang, B.; Li, W.; Xiang, Y.; Peng, X.; Ming, J.; Wu, X.; Zhang, Y.; Xu, Q. Allelic reprogramming of the histone modification H3K4me3 in early mammalian development. Nature 2016, 537, 553. [Google Scholar] [CrossRef]
- Chen, F.; Fu, Q.; Pu, L.; Zhang, P.; Huang, Y.; Hou, Z.; Xu, Z.; Chen, D.; Huang, F.; Deng, T. Integrated analysis of quantitative proteome and transcriptional profiles reveals the dynamic function of maternally expressed proteins after parthenogenetic activation of buffalo oocyte. Mol. Cell. Proteom. 2018, 17, 1875–1891. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Huang, Y.; Wang, Z.; Chen, F.; Huang, D.; Lu, Y.; Liang, X.; Zhang, M. Proteome profile and quantitative proteomic analysis of buffalo (Bubalus bubalis) follicular fluid during follicle development. Int. J. Mol. Sci. 2016, 17, 618. [Google Scholar] [CrossRef] [Green Version]
- Memili, E.; Peddinti, D.; Shack, L.; Nanduri, B.; McCarthy, F.; Sagirkaya, H.; Burgess, S.C. Bovine germinal vesicle oocyte and cumulus cell proteomics. Reproduction 2007, 133, 1107–1120. [Google Scholar] [CrossRef] [Green Version]
- Peddinti, D.; Memili, E.; Burgess, S.C. Proteomics-based systems biology modeling of bovine germinal vesicle stage oocyte and cumulus cell interaction. PLoS ONE 2010, 5, e11240. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, D.R.; Fröhlich, T.; Otte, K.A.; Beck, A.; Habermann, F.A.; Wolf, E.; Arnold, G.J. Stage-Specific proteome signatures in early bovine embryo development. J. Proteome Res. 2014, 13, 4363–4376. [Google Scholar] [CrossRef]
- Ma, M.; Guo, X.; Wang, F.; Zhao, C.; Liu, Z.; Shi, Z.; Wang, Y.; Zhang, P.; Zhang, K.; Wang, N. Protein expression profile of the mouse metaphase-II oocyte. J. Proteome Res. 2008, 7, 4821–4830. [Google Scholar] [CrossRef]
- Vitale, A.M.; Calvert, M.E.K.; Mallavarapu, M.; Yurttas, P.; Perlin, J.; Herr, J.; Coonrod, S. Proteomic profiling of murine oocyte maturation. Mol. Reprod. Dev. 2007, 74, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ni, X.; Guo, Y.; Guo, X.; Wang, Y.; Zhou, Z.; Huo, R.; Sha, J. Proteomic-Based identification of maternal proteins in mature mouse oocytes. BMC Genom. 2009, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kou, Z.; Jing, Z.; Zhang, Y.; Guo, X.; Dong, M.; Wilmut, I.; Gao, S. Proteome of mouse oocytes at different developmental stages. Proc. Natl. Acad. Sci. USA 2010, 107, 17639–17644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, M.J.; Siatkowski, M.; Paudel, Y.; Balbach, S.T.; Baeumer, N.; Crosetto, N.; Drexler, H.C.; Fuellen, G.; Boiani, M. Proteomic analysis of mouse oocytes reveals 28 candidate factors of the “reprogrammome”. J. Proteome Res. 2011, 10, 2140–2153. [Google Scholar] [CrossRef] [PubMed]
- Katz-Jaffe, M.G.; Linck, D.W.; Schoolcraft, W.B.; Gardner, D.K. A proteomic analysis of mammalian preimplantation embryonic development. Reproduction 2005, 130, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.-S.; Jeon, Y.-J.; Kim, D.-W.; Yang, T.-H.; Soh, Y.; Lee, H.K.; Choi, N.-J.; Park, S.-B.; Seo, K.S. Identification of maturation and protein synthesis related proteins from porcine oocytes during in vitro maturation. Proteome Sci. 2011, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Ellederova, Z.; Halada, P.; Man, P.; Kubelka, M.; Motlik, J.; Kovarova, H. Protein patterns of pig oocytes during In Vitro maturation. Biol. Reprod. 2004, 71, 1533–1539. [Google Scholar] [CrossRef]
- Susor, A.; Ellederova, Z.; Jelinkova, L.; Halada, P.; Kavan, D.; Kubelka, M.; Kovarova, H. Proteomic analysis of porcine oocytes during in vitro maturation reveals essential role for the ubiquitin C-terminal hydrolase-L1. Reproduction 2007, 134, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.D.; Manandhar, G.; Spate, L.; Sutovsky, M.; Zimmerman, S.; Sachdev, S.C.; Hannink, M.; Prather, R.S.; Sutovsky, P. Discovery of putative oocyte quality markers by comparative ExacTag proteomics. J. Proteom. Clin. Appl. 2010, 4, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.; Schäfer, J.; Kuhn, K.; Kienle, S.; Schwarz, J.; Schmidt, G.; Neumann, T.; Hamon, C. Tandem mass tags: A novel quantification strategy for comparative analysis of complex protein mixtures by MS/MS. Anal. Chem. 2003, 75, 1895–1904. [Google Scholar] [CrossRef]
- Bishop, D.W. Metabolic conditions within the oviduct of the rabbit. Int. J. Fertil. 1957, 2, 11. [Google Scholar]
- Mastroianni, L.; Jones, R. Oxygen tension within the rabbit fallopian tube. Reproduction 1965, 9, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Graves, C. O2 levels in female rabbit reproductive-tract. Proc. J. Anim. Sci. 1974, 39, 994. [Google Scholar]
- Maas, D.H.; Storey, B.T.; Mastroianni, L., Jr. Oxygen tension in the oviduct of the rhesus monkey (Macaca mulatta). Fertil. Steril. 1976, 27, 1312–1317. [Google Scholar] [CrossRef]
- Kasterstein, E.; Strassburger, D.; Komarovsky, D.; Bern, O.; Komsky, A.; Raziel, A.; Friedler, S.; Ron-El, R. The effect of two distinct levels of oxygen concentration on embryo development in a sibling oocyte study. J. Assist. Reprod. Genet. 2013, 30, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wale, P.; Gardner, D. Time-Lapse analysis of mouse embryo development in oxygen gradients. Reprod. Biomed. Online 2010, 21, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Van Soom, A.; Coopman, F.; Mintiens, K.; Boerjan, M.; Van Zeveren, A.; de Kruif, A.; Peelman, L. Influence of oxygen tension on apoptosis and hatching in bovine embryos cultured In Vitro. Theriogenology 2003, 59, 1585–1596. [Google Scholar] [CrossRef]
- Stamati, K.; Mudera, V.; Cheema, U. Evolution of oxygen utilization in multicellular organisms and implications for cell signalling in tissue engineering. J. Tissue Eng. 2011, 2. [Google Scholar] [CrossRef]
- Fatehi, A.N.; Roelen, B.A.; Colenbrander, B.; Schoevers, E.J.; Gadella, B.M.; Bevers, M.M.; Van Den Hurk, R. Presence of cumulus cells during in vitro fertilization protects the bovine oocyte against oxidative stress and improves first cleavage but does not affect further development. Zygote 2005, 13, 177–185. [Google Scholar] [CrossRef]
- Basini, G.; Simona, B.; Santini, S.E.; Grasselli, F. Reactive oxygen species and anti-oxidant defences in swine follicular fluids. Reprod. Fertil. Dev. 2008, 20, 269–274. [Google Scholar] [CrossRef]
- Mingoti, G.Z.; Castro, V.S.D.C.; Méo, S.C.; Barretto, L.S.S.; Garcia, J.M. The effects of macromolecular and serum supplements and oxygen tension during bovine In Vitro procedures on kinetics of oocyte maturation and embryo development. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Somfai, T.; Inaba, Y.; Aikawa, Y.; Ohtake, M.; Kobayashi, S.; Konishi, K.; Nagai, T.; Imai, K. Development of bovine embryos cultured in CR1aa and IVD101 media using different oxygen tensions and culture systems. Acta Vet. Hung. 2010, 58, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg effect: How does it benefit cancer cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantaleon, M.; Harvey, M.; Pascoe, W.; James, D.; Kaye, P. Glucose transporter GLUT3: Ontogeny, targeting, and role in the mouse blastocyst. Proc. Natl. Acad. Sci. USA 1997, 94, 3795–3800. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schoolcraft, W.B. Noninvasive assessment of human embryo nutrient consumption as a measure of developmental potential. Fertil. Steril. 2001, 76, 1175–1180. [Google Scholar] [CrossRef]
- Gardner, D.K.; Leese, H.J. Assessment of embryo viability prior to transfer by the noninvasive measurement of glucose uptake. J. Exp. Zool. 1987, 242, 103–105. [Google Scholar] [CrossRef]
- Christofk, H.R.; Vander Heiden, M.G.; Harris, M.H.; Ramanathan, A.; Gerszten, R.E.; Wei, R.; Fleming, M.D.; Schreiber, S.L.; Cantley, L.C. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature 2008, 452, 230–233. [Google Scholar] [CrossRef]
- Lunt, S.Y.; Muralidhar, V.; Hosios, A.M.; Israelsen, W.J.; Gui, D.Y.; Newhouse, L.; Ogrodzinski, M.; Hecht, V.; Xu, K.; Acevedo, P.N.M. Pyruvate kinase isoform expression alters nucleotide synthesis to impact cell proliferation. Mol. Cell 2015, 57, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; DeBerardinis, R.J. Understanding the intersections between metabolism and cancer biology. Cell 2017, 168, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Krisher, R.L.; Prather, R.S. A role for the Warburg effect in preimplantation embryo development: Metabolic modification to support rapid cell proliferation. Mol. Reprod. Dev. 2012, 79, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Gasparrini, B.J.T. In Vitro embryo production in buffalo species: State of the art. Theriogenology 2002, 57, 237–256. [Google Scholar] [CrossRef]
- Verma, M.; Pandey, S.; Bhat, I.A.; Mukesh, B.; Anand, J.; Chandra, V.; Sharma, G.T. Impact of l-carnitine on lipid content and post thaw survivability of buffalo embryos produced In Vitro. Cryobiology 2018, 82, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, F. Genetic improvement through reproductive technology. Anim. Reprod. Sci. 1996, 42, 205–214. [Google Scholar] [CrossRef]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development In Vitro versus In Vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef]
- Pereira, R.; Marques, C. Animal oocyte and embryo cryopreservation. Cell Tissue Bank. 2008, 9, 267–277. [Google Scholar] [CrossRef]
- Arav, A.; Zeron, Y.; Leslie, S.; Behboodi, E.; Anderson, G.B.; Crowe, J.H. Phase transition temperature and chilling sensitivity of bovine oocytes. Cryobiology 1996, 33, 589–599. [Google Scholar] [CrossRef]
- Seidel, G.E., Jr. Modifying oocytes and embryos to improve their cryopreservation. Theriogenology 2006, 65, 228–235. [Google Scholar] [CrossRef]
- Rasmussen, T.P. Genomic medicine and lipid metabolism: LDL targets and stem cell research approaches. In Translational Cardiometabolic Genomic Medicine; Elsevier: Amsterdam, The Netherlands, 2016; pp. 99–118. [Google Scholar]
- Merilainen, G.; Poikela, V.; Kursula, P.; Wierenga, R.K. The thiolase reaction mechanism: The importance of Asn316 and His348 for stabilizing the enolate intermediate of the Claisen condensation. Biochemistry 2009, 48, 11011–11025. [Google Scholar] [CrossRef]
- Küch, E.-M.; Vellaramkalayil, R.; Zhang, I.; Lehnen, D.; Brügger, B.; Stremmel, W.; Ehehalt, R.; Poppelreuther, M.; Füllekrug, J. Differentially localized acyl-CoA synthetase 4 isoenzymes mediate the metabolic channeling of fatty acids towards phosphatidylinositol. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 227–239. [Google Scholar] [CrossRef]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef]
- Su, W.; Sun, J.; Shimizu, K.; Kadota, K. TCC-GUI: A shiny-based application for differential expression analysis of RNA-Seq count data. BMC Res. Notes 2019, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Pavani, K.C.; Baron, E.E.; Faheem, M.; Chaveiro, A.; Da Silva, F.M. Optimisation of total RNA extraction from bovine oocytes and embryos for gene expression studies and effects of cryoprotectants on total RNA extraction. Cytol. Genet. 2015, 49, 232–239. [Google Scholar] [CrossRef]
- Sood, T.J.; Lagah, S.V.; Sharma, A.; Singla, S.K.; Mukesh, M.; Chauhan, M.S.; Manik, R.; Palta, P. Selection of suitable internal control genes for accurate normalization of real-time quantitative PCR data of buffalo (Bubalus bubalis) blastocysts produced by SCNT and IVF. Cell. Reprogram. 2017, 19, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters. | N | Cell Count (Mean ± S.E.M) |
|---|---|---|
| 5% Oxygen | 15 | 151.36 ± 2.70 |
| 20% Oxygen | 15 | 89.18 ± 2.60 * |
| Parameters | N | Hatching Rate (Mean ± S.E.M) |
|---|---|---|
| 5% Oxygen | 42 | 82.67 ± 4.44 |
| 20% Oxygen | 38 | 60.69 ± 1.80 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahzad, Q.; Pu, L.; Ahmed Wadood, A.; Waqas, M.; Xie, L.; Shekhar Pareek, C.; Xu, H.; Liang, X.; Lu, Y. Proteomics Analysis Reveals that Warburg Effect along with Modification in Lipid Metabolism Improves In Vitro Embryo Development under Low Oxygen. Int. J. Mol. Sci. 2020, 21, 1996. https://doi.org/10.3390/ijms21061996
Shahzad Q, Pu L, Ahmed Wadood A, Waqas M, Xie L, Shekhar Pareek C, Xu H, Liang X, Lu Y. Proteomics Analysis Reveals that Warburg Effect along with Modification in Lipid Metabolism Improves In Vitro Embryo Development under Low Oxygen. International Journal of Molecular Sciences. 2020; 21(6):1996. https://doi.org/10.3390/ijms21061996
Chicago/Turabian StyleShahzad, Qaisar, Liping Pu, Armughan Ahmed Wadood, Muhammad Waqas, Long Xie, Chandra Shekhar Pareek, Huiyan Xu, Xianwei Liang, and Yangqing Lu. 2020. "Proteomics Analysis Reveals that Warburg Effect along with Modification in Lipid Metabolism Improves In Vitro Embryo Development under Low Oxygen" International Journal of Molecular Sciences 21, no. 6: 1996. https://doi.org/10.3390/ijms21061996
APA StyleShahzad, Q., Pu, L., Ahmed Wadood, A., Waqas, M., Xie, L., Shekhar Pareek, C., Xu, H., Liang, X., & Lu, Y. (2020). Proteomics Analysis Reveals that Warburg Effect along with Modification in Lipid Metabolism Improves In Vitro Embryo Development under Low Oxygen. International Journal of Molecular Sciences, 21(6), 1996. https://doi.org/10.3390/ijms21061996