A Silent Exonic Mutation in a Rice Integrin-α FG-GAP Repeat-Containing Gene Causes Male-Sterility by Affecting mRNA Splicing

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
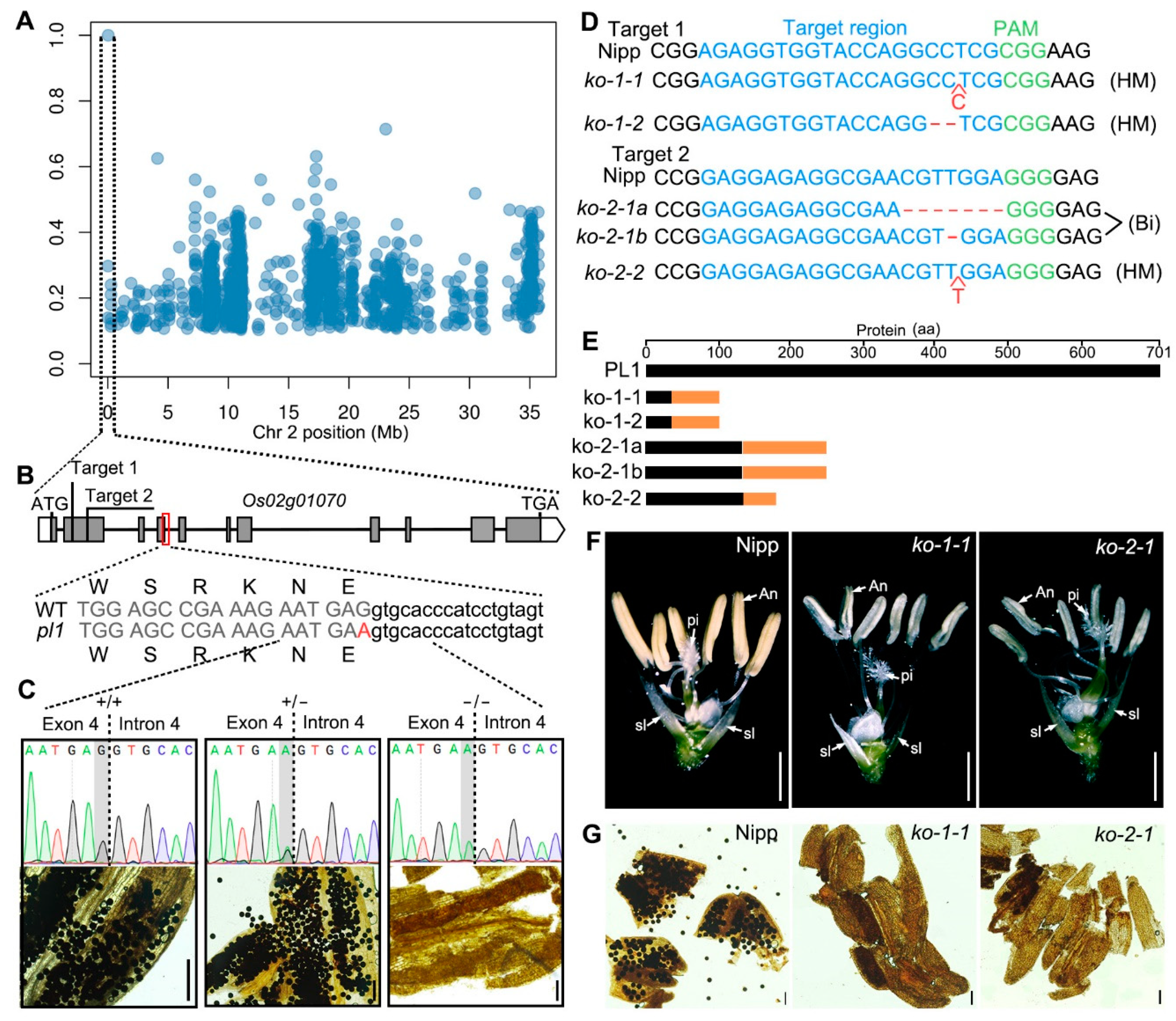
2.1. Isolation of the pl1 Mutant
2.2. Cloning of PL1
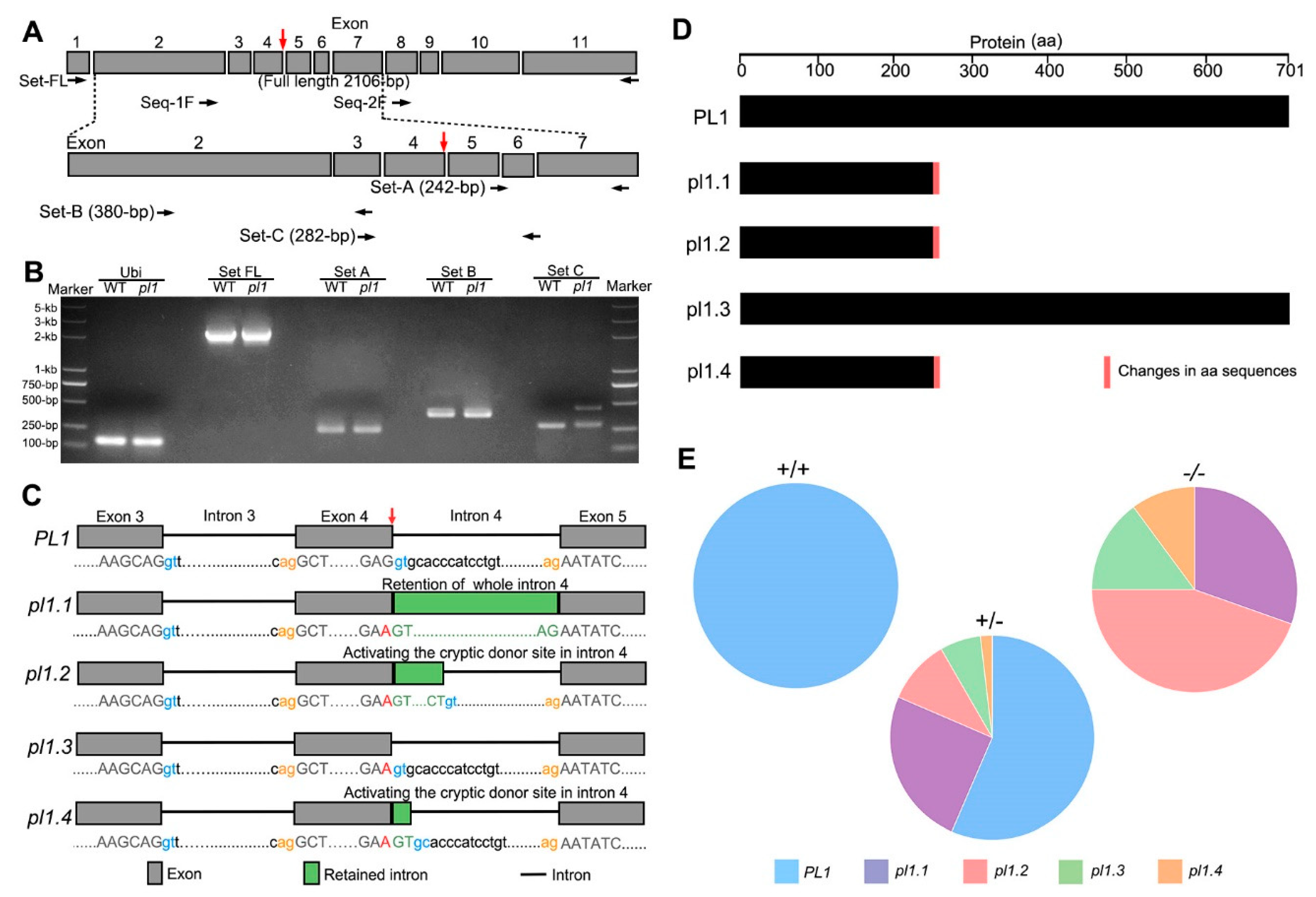
2.3. Normal Splicing of PL1 Is Altered by pl1 Mutation
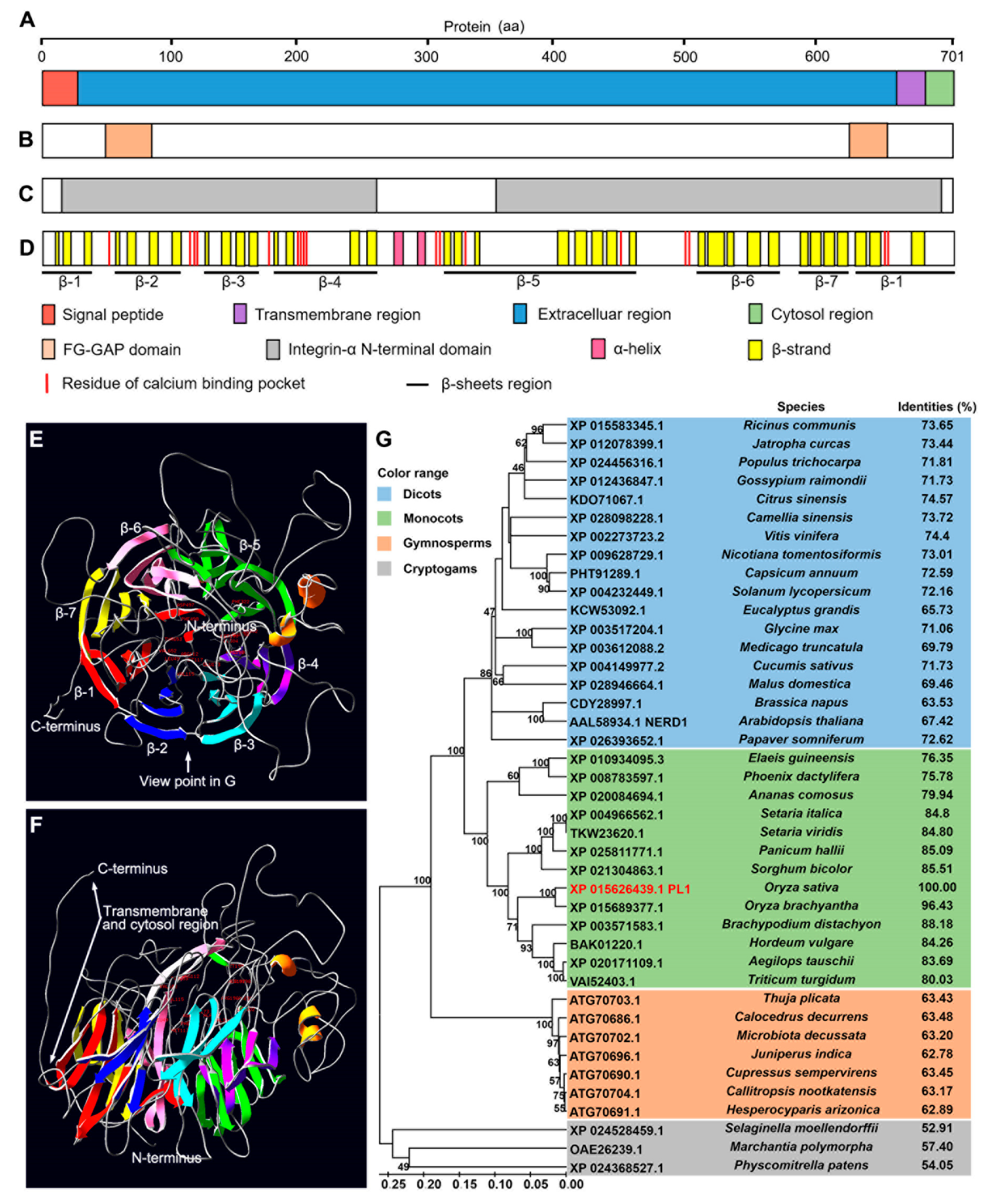
2.4. PL1 Encodes a Conserved Integrin-α FG-GAP Repeat-Containing Protein
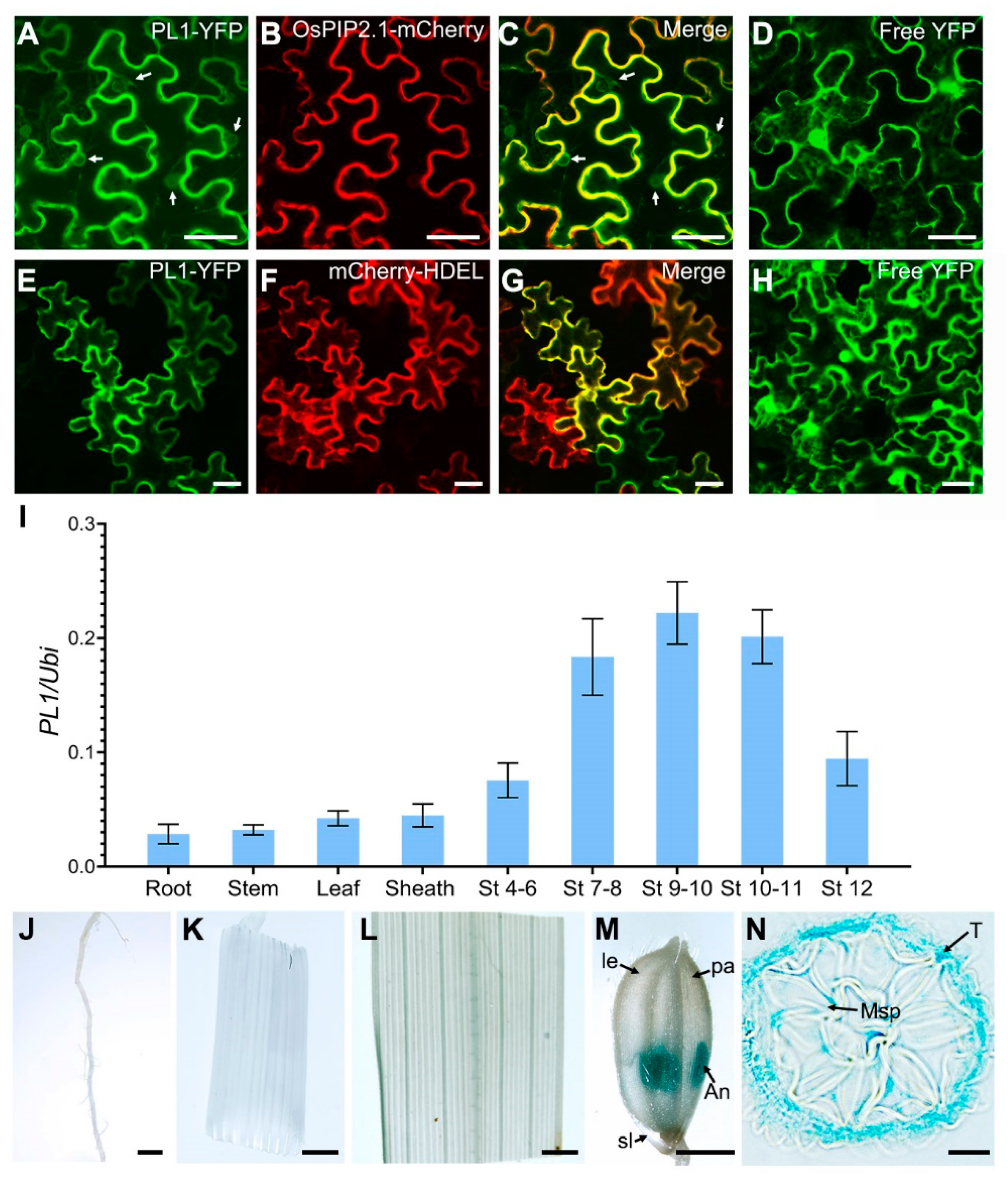
2.5. Expression Analysis of PL1
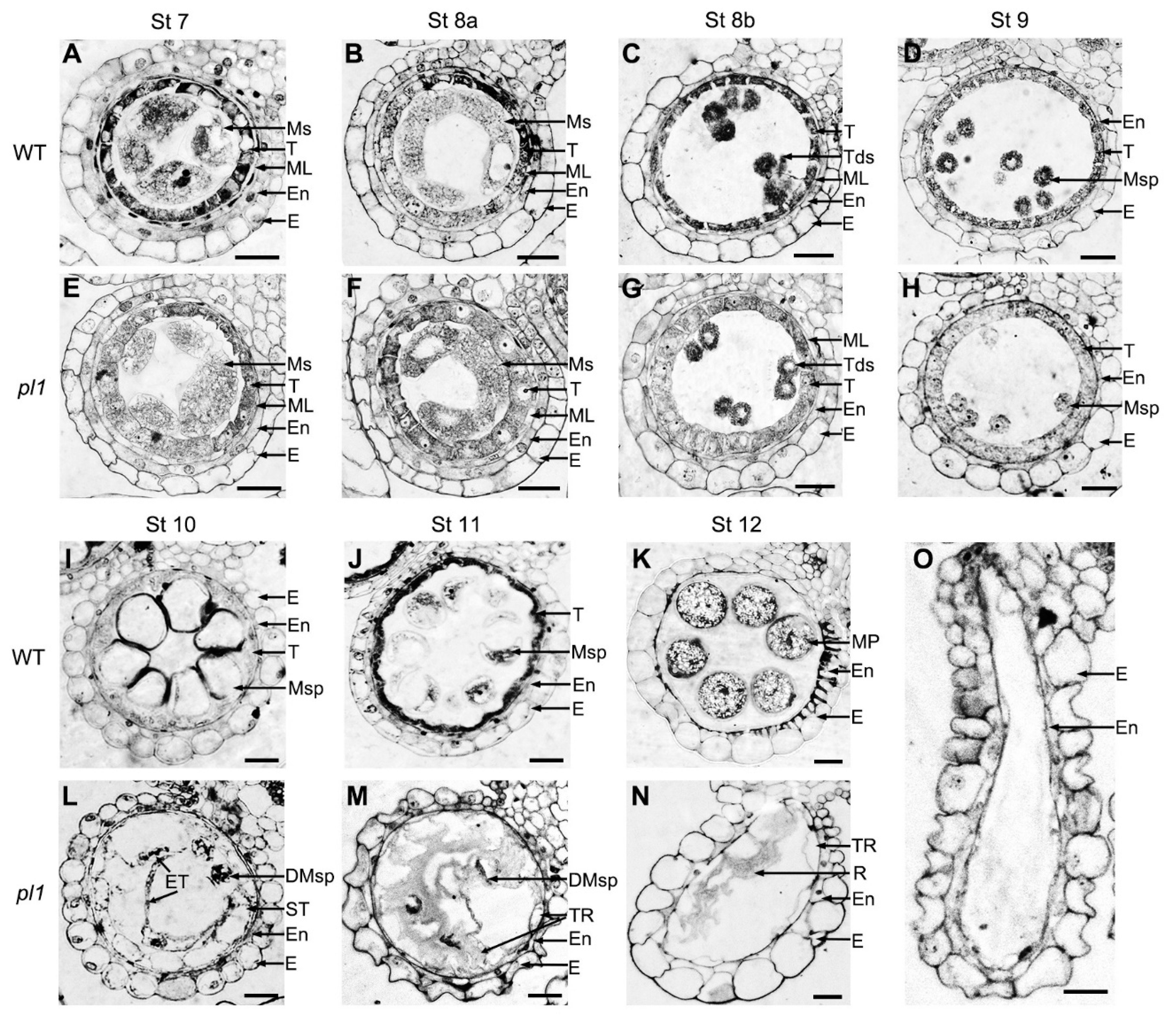
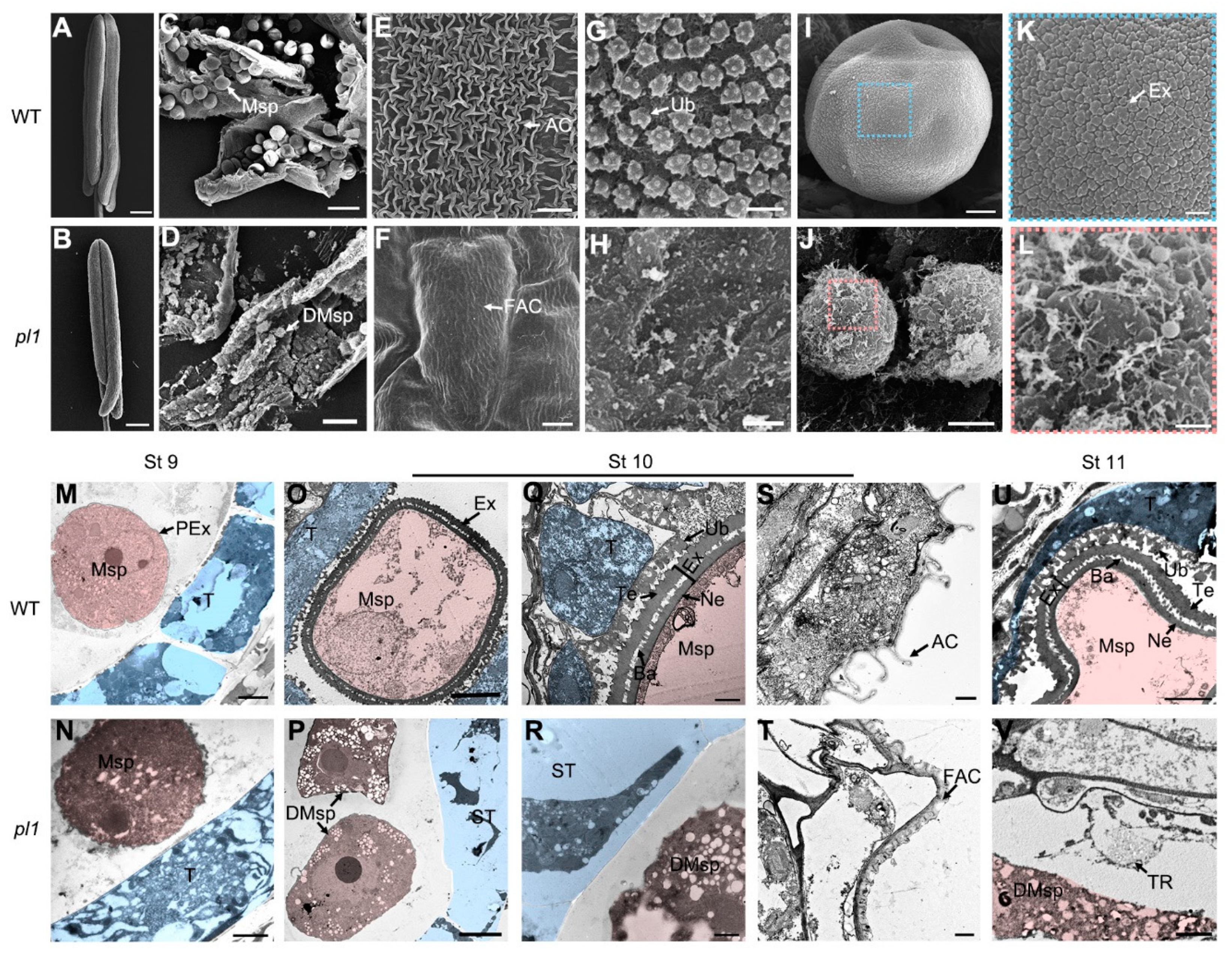
2.6. Cytological Observations of pl1 Anthers
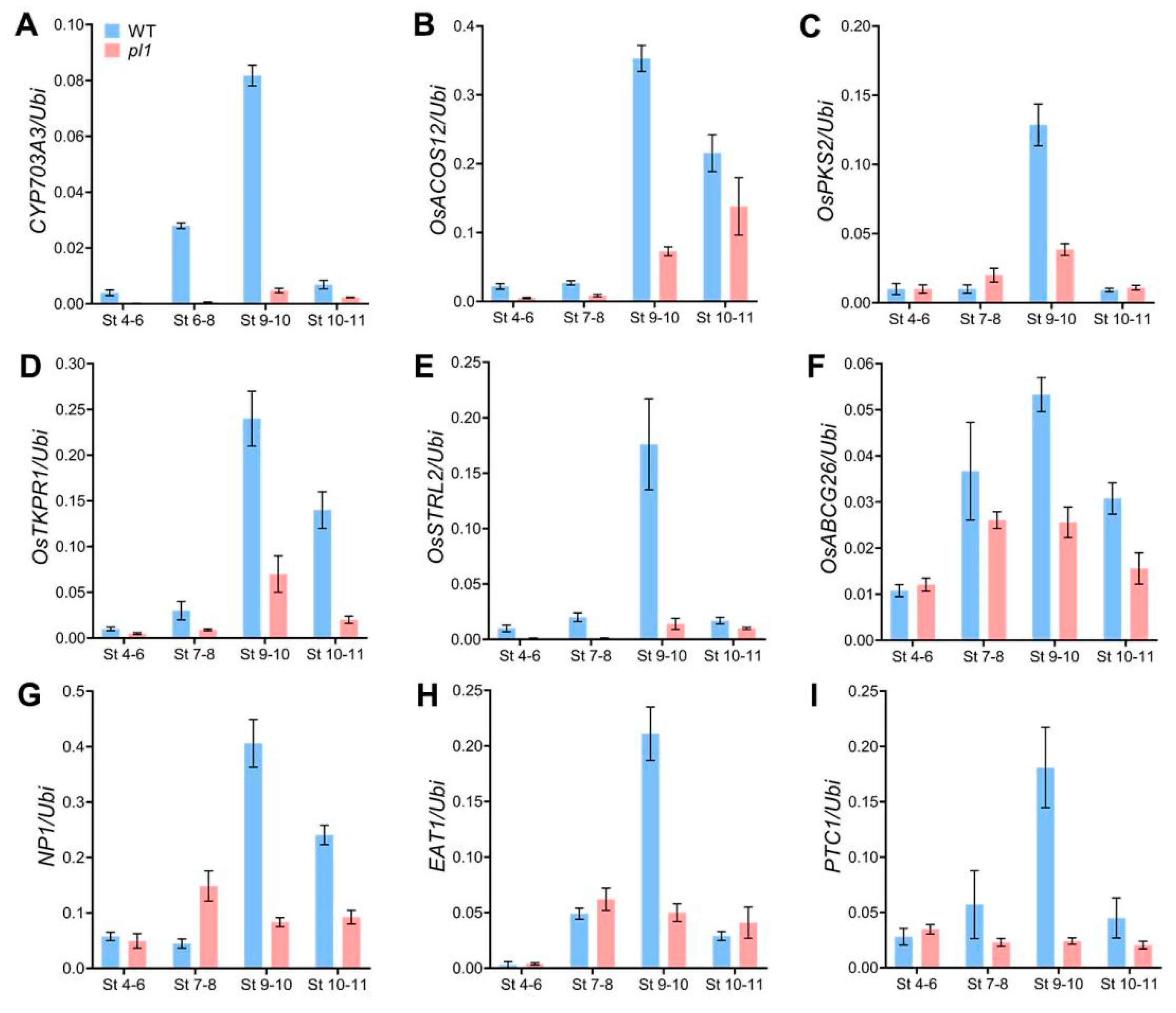
2.7. The pl1 Mutation Altered the Expression of Genes Related to Anther and Pollen Development
3. Discussion
3.1. The pl1 Mutation Affects Splice Donor Site Selection
3.2. Normal Rice Male Development Requires a Certain Amount of Functional PL1 Protein
3.3. Role of PL1 in Anther and Pollen Development
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Phenotypic and Microscopic Observations
4.3. Mapping the pl1 Mutation
4.4. Pre-mRNA Secondary Structure Prediction
4.5. RNA Extraction and RT-PCR Analysis
4.6. Histochemical GUS Assay
4.7. Subcellular Localization Analysis of PL1
4.8. Protein Sequence Analysis of PL1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, A.; Coimbra, S. Advances in plant reproduction: from gametes to seeds. J. Exp. Bot. 2019, 70, 2933–2936. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Schmid, M.; Grossniklaus, U. Plant germline formation: common concepts and developmental flexibility in sexual and asexual reproduction. Development 2015, 142, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, F.; Twell, D. Germline Specification and Function in Plants. Annu. Rev. Plant Biol. 2011, 62, 461–484. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-N. Regulation of sporopollenin synthesis for pollen wall formation in plant. Sci. China Life Sci. 2016, 59, 1335–1337. [Google Scholar] [CrossRef] [Green Version]
- Ariizumi, T.; Toriyama, K. Genetic Regulation of Sporopollenin Synthesis and Pollen Exine Development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef]
- Gómez, J.F.; Talle, B.; Wilson, Z.A. Anther and pollen development: A conserved developmental pathway. J. Integr. Plant Biol. 2015, 57, 876–891. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S. Pollen. Curr. Biol. 2013, 23, R988–R990. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Cui, M.; Yang, L.; Kim, Y.-J.; Zhang, D. Genetic and Biochemical Mechanisms of Pollen Wall Development. Trends Plant Sci. 2015, 20, 741–753. [Google Scholar] [CrossRef]
- Zhang, D.B.; Luo, X.; Zhu, L. Cytological analysis and genetic control of rice anther development. J. Genet. Genom. 2011, 38, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.B.; Wilson, Z.A. Stamen specification and anther development in rice. Chin. Sci. Bull. 2009, 54, 2342–2353. [Google Scholar] [CrossRef]
- Ma, H. A Molecular Portrait of Arabidopsis Meiosis. Arab. Book 2006, 4, e0095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Lou, Y.; Xu, X.; Yang, Z.-N. A Genetic Pathway for Tapetum Development and Function in Arabidopsis. J. Integr. Plant Biol. 2011, 53, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fan, X.-d. Tapetum: regulation and role in sporopollenin biosynthesis in Arabidopsis. Plant Mol. Biol. 2013, 83, 165–175. [Google Scholar] [CrossRef]
- Li, D.-D.; Xue, J.-S.; Zhu, J.; Yang, Z.-N. Gene Regulatory Network for Tapetum Development in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhang, D. Biosynthesis of anther cuticle and pollen exine in rice. Plant Signal. Behav. 2010, 5, 1121–1123. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Shi, J.; Liang, W.; Zhang, D. ATP binding cassette G transporters and plant male reproduction. Plant Signal. Behav. 2016, 11, e1136764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.-H.; Han, M.-J.; Lee, Y.-S.; Kim, Y.-W.; Hwang, I.; Kim, M.-J.; Kim, Y.-K.; Nahm, B.H.; An, G. Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell 2005, 17, 2705–2722. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, D.-S.; Liu, H.-S.; Yin, C.-S.; Li, X.-x.; Liang, W.-q.; Yuan, Z.; Xu, B.; Chu, H.-W.; Wang, J.; et al. The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development. Plant Cell 2006, 18, 2999–3014. [Google Scholar] [CrossRef] [Green Version]
- Niu, N.; Liang, W.; Yang, X.; Jin, W.; Wilson, Z.A.; Hu, J.; Zhang, D. EAT1 promotes tapetal cell death by regulating aspartic proteases during male reproductive development in rice. Nat. Commun. 2013, 4, 1445. [Google Scholar] [CrossRef]
- Fu, Z.; Yu, J.; Cheng, X.; Zong, X.; Xu, J.; Chen, M.; Li, Z.; Zhang, D.; Liang, W. The Rice Basic Helix-Loop-Helix Transcription Factor TDR INTERACTING PROTEIN2 Is a Central Switch in Early Anther Development. Plant Cell 2014, 26, 1512–1524. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.-S.; Li, M.-J.; Sun-Ben Ku, M.; Ho, Y.-C.; Lin, Y.-J.; Chuang, M.-H.; Hsing, H.-X.; Lien, Y.-C.; Yang, H.-T.; Chang, H.-C.; et al. The bHLH142 Transcription Factor Coordinates with TDR1 to Modulate the Expression of EAT1 and Regulate Pollen Development in Rice. Plant Cell 2014, 26, 2486–2504. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Liu, L.; Sun, L.; Yu, P.; Zhang, P.; Abbas, A.; Xiang, X.; Wu, W.; Zhang, Y.; Cao, L. OsMS1 functions as a transcriptional activator to regulate programmed tapetum development and pollen exine formation in rice. Plant Mol. Biol. 2019. [Google Scholar] [CrossRef]
- Li, H.; Yuan, Z.; Vizcay-Barrena, G.; Yang, C.; Liang, W.; Zong, J.; Wilson, Z.A.; Zhang, D. PERSISTENT TAPETAL CELL1 encodes a PHD-finger protein that is required for tapetal cell death and pollen development in rice. Plant Physiol. 2011, 156, 615–630. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.J.; Wu, D.; Shi, J.X.; He, Y.; Pinot, F.; Grausem, B.; Yin, C.S.; Zhu, L.; Chen, M.J.; Luo, Z.J.; et al. Rice CYP703A3, a cytochrome P450 hydroxylase, is essential for development of anther cuticle and pollen exine. J. Integr. Plant Biol. 2014, 56, 979–994. [Google Scholar] [CrossRef]
- Li, H.; Pinot, F.; Sauveplane, V.; Werck-Reichhart, D.; Diehl, P.; Schreiber, L.; Franke, R.; Zhang, P.; Chen, L.; Gao, Y.; et al. Cytochrome P450 Family Member CYP704B2 Catalyzes the ω -Hydroxylation of Fatty Acids and Is Required for Anther Cutin Biosynthesis and Pollen Exine Formation in Rice. Plant Cell 2010, 22, 173–190. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, D.; Guo, Z.; Shi, Q.; Xiong, S.; Zhang, C.; Zhu, J.; Yang, Z. OsACOS12, an orthologue of Arabidopsis acyl-CoA synthetase5, plays an important role in pollen exine formation and anther development in rice. BMC Plant Biol. 2016, 16, 256. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; He, Z.; Qu, L.; Liu, M.; Zeng, J.; Liang, Y.; Wang, T.; Chen, D.; Xiao, Q.; Zhu, J.; et al. Knockout of OsACOS12 caused male sterility in rice. Mol. Breed. 2017, 37, 126. [Google Scholar] [CrossRef]
- Zhu, X.; Yu, J.; Shi, J.; Tohge, T.; Fernie, A.R.; Meir, S.; Aharoni, A.; Xu, D.; Zhang, D.; Liang, W. The polyketide synthase OsPKS2 is essential for pollen exine and Ubisch body patterning in rice. J. Integr. Plant Biol. 2017, 59, 612–628. [Google Scholar] [CrossRef]
- Zou, T.; Liu, M.; Xiao, Q.; Wang, T.; Chen, D.; Luo, T.; Yuan, G.; Li, Q.; Zhu, J.; Liang, Y.; et al. OsPKS2 is required for rice male fertility by participating in pollen wall formation. Plant Cell Rep. 2018, 37, 759–773. [Google Scholar] [CrossRef]
- Zou, T.; Xiao, Q.; Li, W.; Luo, T.; Yuan, G.; He, Z.; Liu, M.; Li, Q.; Xu, P.; Zhu, J.; et al. OsLAP6/OsPKS1, an orthologue of Arabidopsis PKSA/LAP6, is critical for proper pollen exine formation. Rice 2017, 10, 53. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, Y.-C.; So, J.; Du, Y.; Lo, C. Conserved metabolic steps for sporopollenin precursor formation in tobacco and rice. Physiol. Plant. 2013, 149, 13–24. [Google Scholar] [CrossRef]
- Xu, D.; Qu, S.; Tucker, M.R.; Zhang, D.; Liang, W.; Shi, J. Ostkpr1 functions in anther cuticle development and pollen wall formation in rice. BMC Plant Biol. 2019, 19, 104. [Google Scholar] [CrossRef] [Green Version]
- Qin, P.; Tu, B.; Wang, Y.; Deng, L.; Quilichini, T.D.; Li, T.; Wang, H.; Ma, B.; Li, S. ABCG15 encodes an ABC transporter protein, and is essential for post-meiotic anther and pollen exine development in rice. Plant Cell Physiol 2013, 54, 138–154. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Shi, J.; Liang, W.; Xue, F.; Luo, Q.; Zhu, L.; Qu, G.; Chen, M.; Schreiber, L.; Zhang, D. Two ATP Binding Cassette G Transporters, Rice ATP Binding Cassette G26 and ATP Binding Cassette G15, Collaboratively Regulate Rice Male Reproduction. Plant Physiol. 2015, 169, 2064–2079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liang, W.; Yin, C.; Zong, J.; Gu, F.; Zhang, D. OsC6, encoding a lipid transfer protein, is required for postmeiotic anther development in rice. Plant Physiol. 2010, 154, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Fellenberg, C.; Vogt, T. Evolutionarily conserved phenylpropanoid pattern on angiosperm pollen. Trends Plant Sci. 2015, 20, 212–218. [Google Scholar] [CrossRef]
- De Conti, L.; Baralle, M.; Buratti, E. Exon and intron definition in pre-mRNA splicing. Wiley Interdiscip. Rev.. RNA 2013, 4. [Google Scholar] [CrossRef]
- Cheah, M.T.; Wachter, A.; Sudarsan, N.; Breaker, R.R. Control of alternative RNA splicing and gene expression by eukaryotic riboswitches. Nature 2007, 447, 497–500. [Google Scholar] [CrossRef]
- Zlotorynski, E. Intron definition, exon definition and back-splicing revisited. Nat. Rev. Mol. Cell Biol. 2019, 20, 661. [Google Scholar] [CrossRef] [Green Version]
- Lopez, A.J. ALTERNATIVE SPLICING OF PRE-mRNA: Developmental Consequences and Mechanisms of Regulation. Annu. Rev. Genet. 1998, 32, 279–305. [Google Scholar] [CrossRef]
- Deng, X.; Cao, X. Roles of pre-mRNA splicing and polyadenylation in plant development. Curr. Opin. Plant Biol. 2017, 35, 45–53. [Google Scholar] [CrossRef]
- Scotti, M.M.; Swanson, M.S. RNA mis-splicing in disease. Nat. Rev. Genet. 2016, 17, 19–32. [Google Scholar] [CrossRef]
- Simpson, G.G.; Filipowicz, W. Splicing of precursors to mRNA in higher plants: mechanism, regulation and sub-nuclear organisation of the spliceosomal machinery. Plant Mol. Biol. 1996, 32, 1–41. [Google Scholar] [CrossRef]
- Hirano, H.Y.; Eiguchi, M.; Sano, Y. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol. 1998, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.-F.; Lu, Y.; Wang, M.; yu, H.; Yi, C.; Tang, S.; et al. Wx, the Ancestral Allele of Rice Waxy Gene. Mol. Plant 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-Y.; Zheng, F.-Q.; Shen, G.-Z.; Gao, J.-P.; Snustad, D.P.; Li, M.-G.; Zhang, J.-L.; Hong, M.-M. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J. 1995, 7, 613–622. [Google Scholar] [CrossRef]
- Johnson, K.C.M.; Yu, Y.; Gao, L.; Eng, R.C.; Wasteneys, G.O.; Chen, X.; Li, X. A partial loss-of-function mutation in an Arabidopsis RNA polymerase III subunit leads to pleiotropic defects. J. Exp. Bot. 2016, 67, 2219–2230. [Google Scholar] [CrossRef] [Green Version]
- Hématy, K.; Bellec, Y.; Podicheti, R.; Bouteiller, N.; Anne, P.; Morineau, C.; Haslam, R.P.; Beaudoin, F.; Napier, J.A.; Mockaitis, K.; et al. The Zinc-Finger Protein SOP1 Is Required for a Subset of the Nuclear Exosome Functions in Arabidopsis. PLoS Genet. 2016, 12, e1005817. [Google Scholar] [CrossRef] [Green Version]
- Bellec, Y.; Harrar, Y.; Butaeye, C.; Darnet, S.; Bellini, C.; Faure, J.-D. Pasticcino2 is a protein tyrosine phosphatase-like involved in cell proliferation and differentiation in Arabidopsis. Plant J. 2002, 32, 713–722. [Google Scholar] [CrossRef]
- Schwarte, S.; Bauwe, H. Identification of the Photorespiratory 2-Phosphoglycolate Phosphatase, PGLP1, in Arabidopsis. Plant Physiol. 2007, 144, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnolegy 2012, 30, 174–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondol, P.C.; Xu, D.; Duan, L.; Shi, J.; Wang, C.; Chen, X.; Chen, M.; Hu, J.; Liang, W.; Zhang, D. Defective Pollen Wall 3 (DPW3), a novel alpha integrin-like protein, is required for pollen wall formation in rice. New Phytol. 2019, n/a. [Google Scholar] [CrossRef] [PubMed]
- Buratti, E.; Baralle, F.E. Influence of RNA secondary structure on the pre-mRNA splicing process. Mol Cell Biol 2004, 24, 10505–10514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nackley, A.G.; Shabalina, S.A.; Tchivileva, I.E.; Satterfield, K.; Korchynskyi, O.; Makarov, S.S.; Maixner, W.; Diatchenko, L. Human catechol-O-methyltransferase haplotypes modulate protein expression by altering mRNA secondary structure. Science 2006, 5807, 1930–1933. [Google Scholar] [CrossRef] [Green Version]
- Springer, T. Folding of the N-terminal, ligand-binding region of integrin -subunits into a -propeller domain. Proc. Natl. Acad. Sci. United States Am. 1997, 94, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Peremyslov, V.; Why, S.; Moussaoui, I.; Ketter, A.; Cool, R.; Moreno, M.; Vejlupkova, Z.; Dolja, V.; Fowler, J. A Broadly-Conserved NERD Genetically Interacts with the Exocyst to Affect Root Growth and Cell Expansion. J. Exp. Bot. 2018, 69. [Google Scholar] [CrossRef]
- Yuan, J.; Kessler, S. A Genome-Wide Association Study Reveals a Novel Regulator of Ovule Number and Fertility in Arabidopsis thaliana. PLoS Genet. 2018, 15, e1007934. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Li, S.; Liu, M.; Wang, T.; Xiao, Q.; Chen, D.; Li, Q.; Liang, Y.; Zhu, J.; Liang, Y.; et al. An atypical strictosidine synthase, OsSTRL2, plays key roles in anther development and pollen wall formation in rice. Sci. Rep. 2017, 7, 6863. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Chen, Z.; Yan, W.; Xie, G.; Lu, J.; Yao, N.; Yang, G.; Xia, J.; Tang, X. An ABC transporter, OsABCG26, is required for anther cuticle and pollen exine formation and pollen-pistil interactions in rice. Plant Sci. 2016, 253, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Chen, Z.; Wang, N.; Xie, G.; Lu, J.; Yan, W.; Zhou, J.; Tang, X.; Deng, X.W. Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene. Proc. Natl. Acad. Sci. 2016, 113, 14145–14150. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Lin, S.; Shi, J.; Yu, J.; Zhu, L.; Yang, X.; Zhang, D.; Liang, W. Rice No Pollen 1 (NP1) is required for anther cuticle formation and pollen exine patterning. Plant J. 2017, 91, 263–277. [Google Scholar] [CrossRef] [Green Version]
- Padgett, R.A.; Grabowski, P.J.; Konarska, M.M.; Seiler, S.; Sharp, P.A. SPLICING OF MESSENGER RNA PRECURSORS. Annu. Rev. Biochem. 1986, 55, 1119–1150. [Google Scholar] [CrossRef]
- Patthy, L. Introns and exons. Curr. Opin. Struct. Biol. 1994, 4, 383–392. [Google Scholar] [CrossRef]
- Maniatis, T.; Reed, R. The role of small nuclear ribonucleoprotein particles in pre-mRNA splicing. Nature 1987, 325, 673–678. [Google Scholar] [CrossRef]
- Reed, R.; Griffith, J.; Maniatis, T. Purification and visualization of native spliceosomes. Cell 1988, 53, 949–961. [Google Scholar] [CrossRef]
- Huang, S.; Spector, D.L. Will the real splicing sites please light up? Curr. Biol. 1992, 2, 188–190. [Google Scholar] [CrossRef]
- Orkin, S.; Kazazian, H. The Mutation and Polymorphism of the Human beta-Globin Gene and its Surrounding DNA. Annu. Rev. Genet. 1984, 18, 131–171. [Google Scholar] [CrossRef]
- Reed, R.; Maniatis, T. Intron sequences involved in lariat formation during pre-mRNA splicing. Cell 1985, 41, 95–105. [Google Scholar] [CrossRef]
- Shapiro, M.B.; Senapathy, P. RNA splice junctions of different classes of eukaryotes: sequence statistics and functional implications in gene expression. Nucleic Acids Res. 1987, 15, 7155–7174. [Google Scholar] [CrossRef] [Green Version]
- Carmel, I.; Tal, S.; Vig, I.; Ast, G. Comparative analysis detects dependencies among the 5\” splice-site positions. RNA 2004, 10, 828–840. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, D.S.; Krainer, A.R. Mechanisms for selecting 5′ splice sites in mammalian pre-mRNA splicing. Trends Genet. 1994, 10, 100–106. [Google Scholar] [CrossRef]
- Jin, Y.; Yang, Y.; Zhang, P. New insights into RNA secondary structure in the alternative splicing of pre-mRNAs. RNA Biol. 2011, 8, 450–457. [Google Scholar] [CrossRef]
- Rubtsov, P.M. Role of pre-mRNA secondary structures in the regulation of alternative splicing. Mol. Biol. 2016, 50, 823–830. [Google Scholar] [CrossRef]
- Ding, Y.; Tang, Y.; Kwok, C.K.; Zhang, Y.; Bevilacqua, P.C.; Assmann, S.M. In vivo genome-wide profiling of RNA secondary structure reveals novel regulatory features. Nature 2014, 505, 696–700. [Google Scholar] [CrossRef]
- Wu, Q.; Krainer, A.R. U1-Mediated Exon Definition Interactions Between AT-AC and GT-AG Introns. Science 1996, 274, 1005. [Google Scholar] [CrossRef]
- McCullough, A.J.; Lou, H.; Schuler, M.A. Factors affecting authentic 5’ splice site selection in plant nuclei. Mol Cell Biol 1993, 13, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, D.M.; Drabenstot, S.D.; Buchanan, K.L.; Lai, H.; Zhu, H.; Dyer, D.W.; Roe, B.A.; Murphy, J.W. Introns and splicing elements of five diverse fungi. Eukaryot Cell 2004, 3, 1088–1100. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Smant, G.; Stokkermans, J.; Qin, L.; Helder, J.; Baum, T.; Schots, A.; Davis, E. Genomic organization of four β-1,4-endoglucanase genes in plant-parasitic cyst nematodes and its evolutionary implications. Gene 1998, 220, 61–70. [Google Scholar] [CrossRef]
- Chiu, E.Y.-H.; Lin, Y.-H.; Wu, W.; Song, Q.; Chang, P.-F.L.; Gao, L.-Y.; Chou, C.-C.; Ueng, P.P. Alternative splicing and genetic diversity of the white collar-1 (wc-1) gene in cereal Phaeosphaeria pathogens. Eur. J. Plant Pathol. 2010, 127, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Meng, Z.; Liang, W.; Kudla, J.; Tucker, M.; Luo, Z.; Chen, M.; Xu, D.; Zhao, G.; Wang, J.; et al. A Rice Ca Binding Protein Is Required for Tapetum Function and Pollen Formation. Plant Physiol. 2016, 172, 1772–1786. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zhang, D.b. Molecular Control of Male Fertility for Crop Hybrid Breeding. Trends Plant Sci. 2018, 23, 53–65. [Google Scholar] [CrossRef]
- Wu, Y.; Fox, T.W.; Trimnell, M.R.; Wang, L.; Xu, R.-j.; Cigan, A.M.; Huffman, G.A.; Garnaat, C.W.; Hershey, H.; Albertsen, M.C. Development of a novel recessive genetic male sterility system for hybrid seed production in maize and other cross-pollinating crops. Plant Biotechnol. J. 2016, 14, 1046–1054. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Chen, D.; Zou, T.; Zeng, J.; Gao, F.; He, Z.; Zhou, D.; He, Z.; Yuan, G.; Liu, M.; et al. Defective Leptotene Chromosome 1 (DLC1) encodes a type-B response regulator and is required for rice meiosis. Plant J. 2019, 99, 556–570. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Dangol, S.; Singh, R.; Chen, Y.; Jwa, N.-S. Visualization of Multicolored in vivo Organelle Markers for Co-Localization Studies in Oryza sativa. Mol Cells 2017, 40, 828–836. [Google Scholar] [CrossRef]
- Gomord, V.; Denmat, L.A.; Fitchette-Lainé, A.C.; Satiat-Jeunemaitre, B.; Faye, L. The C-terminal HDEL sequence is sufficient for retention of secretory proteins in the endoplasmic reticulum (ER) but promotes vacuolar targeting of proteins that escape the ER. Plant J. 1997, 11, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Lichocka, M.; Schmelzer, E. Subcellular Localization Experiments and FRET-FLIM Measurements in Plants. Bio-protocol 2014, 4, e1018. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol Biol Evol 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, T.; Zhou, D.; Li, W.; Yuan, G.; Tao, Y.; He, Z.; Zhang, X.; Deng, Q.; Wang, S.; Zheng, A.; et al. A Silent Exonic Mutation in a Rice Integrin-α FG-GAP Repeat-Containing Gene Causes Male-Sterility by Affecting mRNA Splicing. Int. J. Mol. Sci. 2020, 21, 2018. https://doi.org/10.3390/ijms21062018
Zou T, Zhou D, Li W, Yuan G, Tao Y, He Z, Zhang X, Deng Q, Wang S, Zheng A, et al. A Silent Exonic Mutation in a Rice Integrin-α FG-GAP Repeat-Containing Gene Causes Male-Sterility by Affecting mRNA Splicing. International Journal of Molecular Sciences. 2020; 21(6):2018. https://doi.org/10.3390/ijms21062018
Chicago/Turabian StyleZou, Ting, Dan Zhou, Wenjie Li, Guoqiang Yuan, Yang Tao, Zhiyuan He, Xu Zhang, Qiming Deng, Shiquan Wang, Aiping Zheng, and et al. 2020. "A Silent Exonic Mutation in a Rice Integrin-α FG-GAP Repeat-Containing Gene Causes Male-Sterility by Affecting mRNA Splicing" International Journal of Molecular Sciences 21, no. 6: 2018. https://doi.org/10.3390/ijms21062018
APA StyleZou, T., Zhou, D., Li, W., Yuan, G., Tao, Y., He, Z., Zhang, X., Deng, Q., Wang, S., Zheng, A., Zhu, J., Liang, Y., Liu, H., Wang, A., Wang, L., Li, P., & Li, S. (2020). A Silent Exonic Mutation in a Rice Integrin-α FG-GAP Repeat-Containing Gene Causes Male-Sterility by Affecting mRNA Splicing. International Journal of Molecular Sciences, 21(6), 2018. https://doi.org/10.3390/ijms21062018