Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression

Abstract
:1. Introduction
2. Results
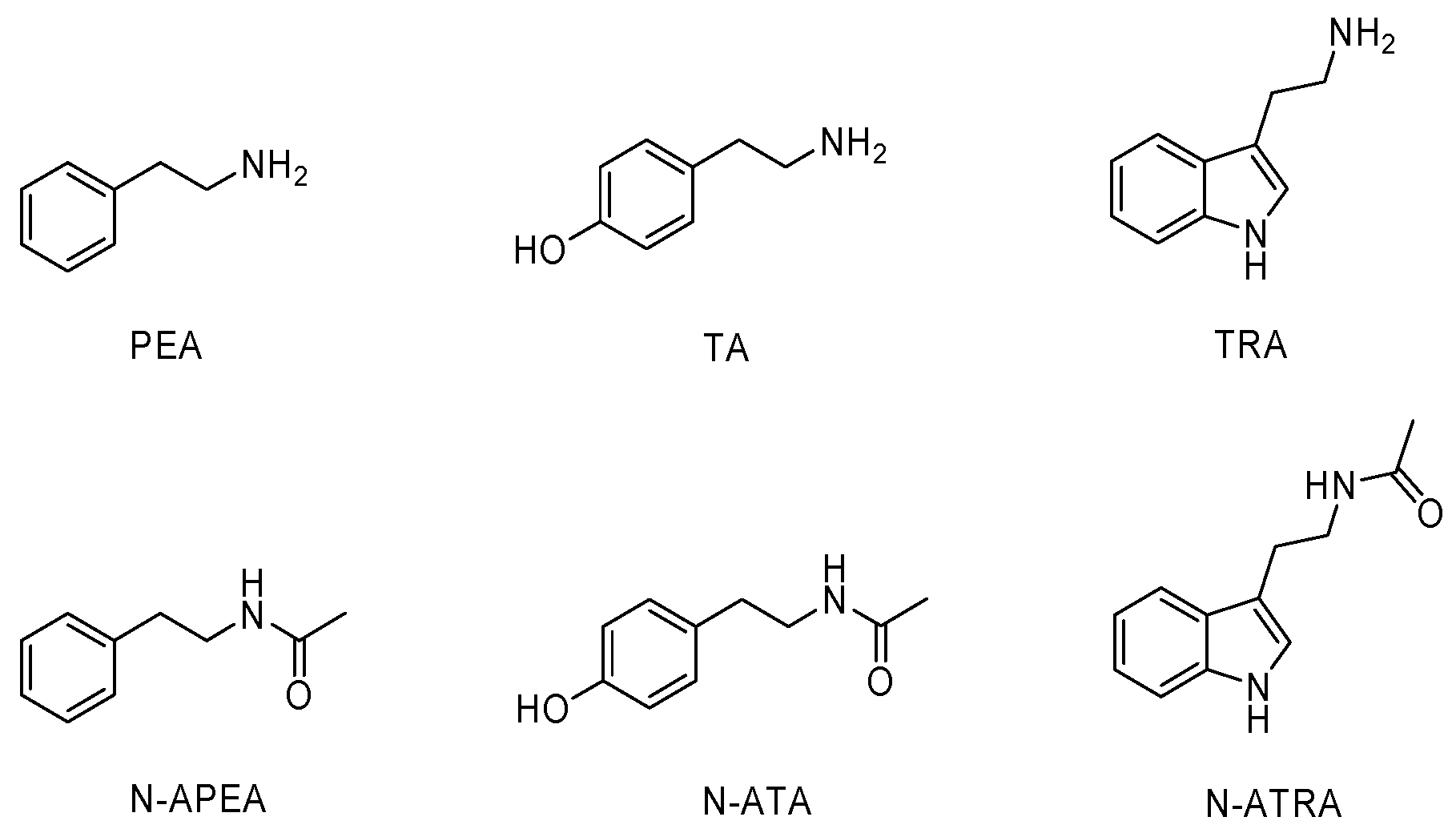
2.1. Identification and Synthesis
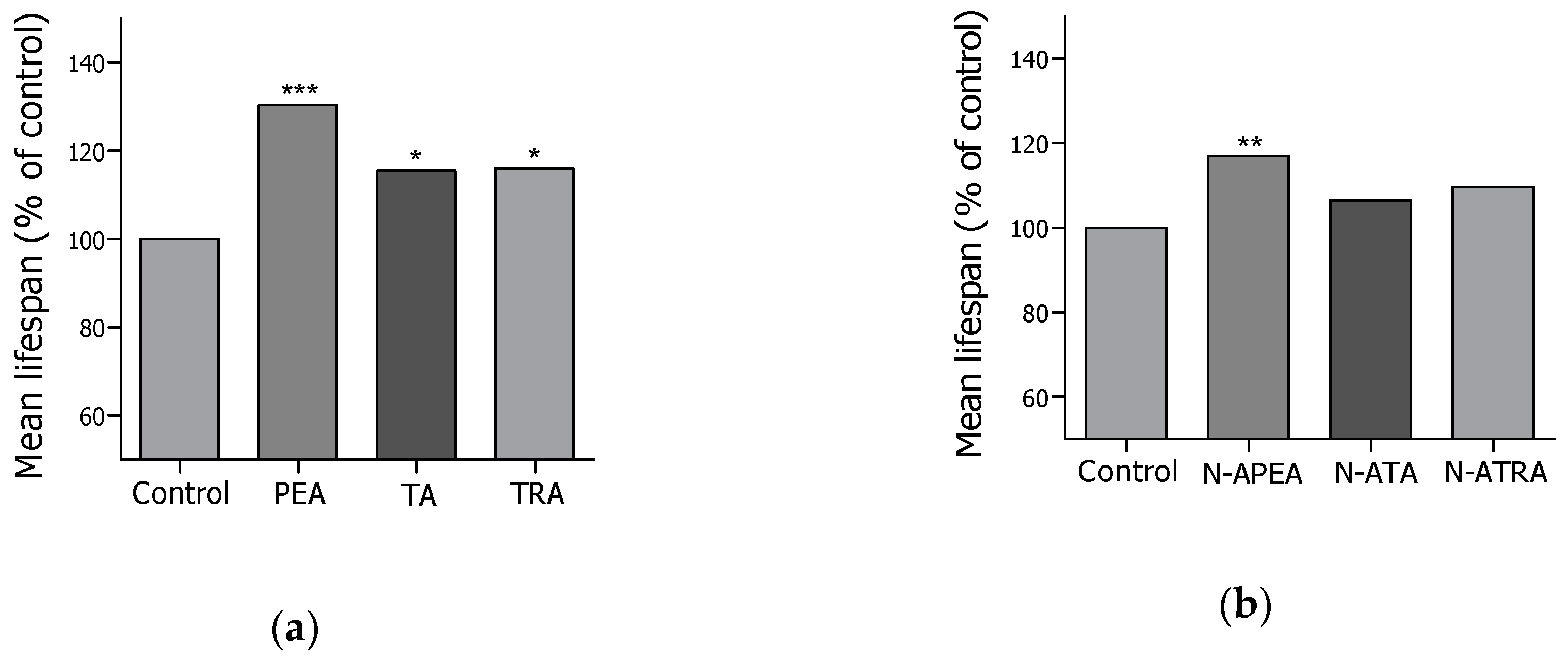
2.2. Lifespan Screening
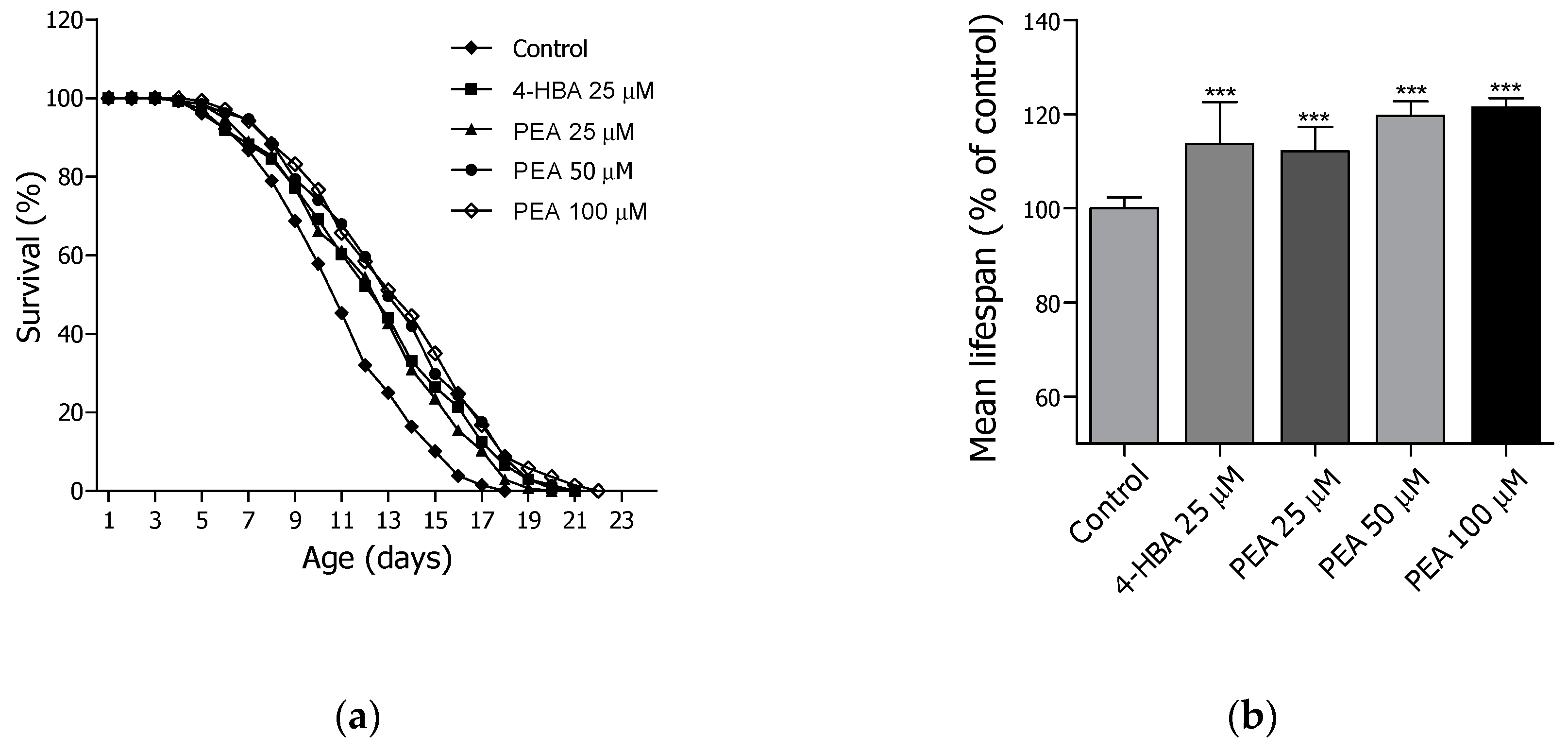
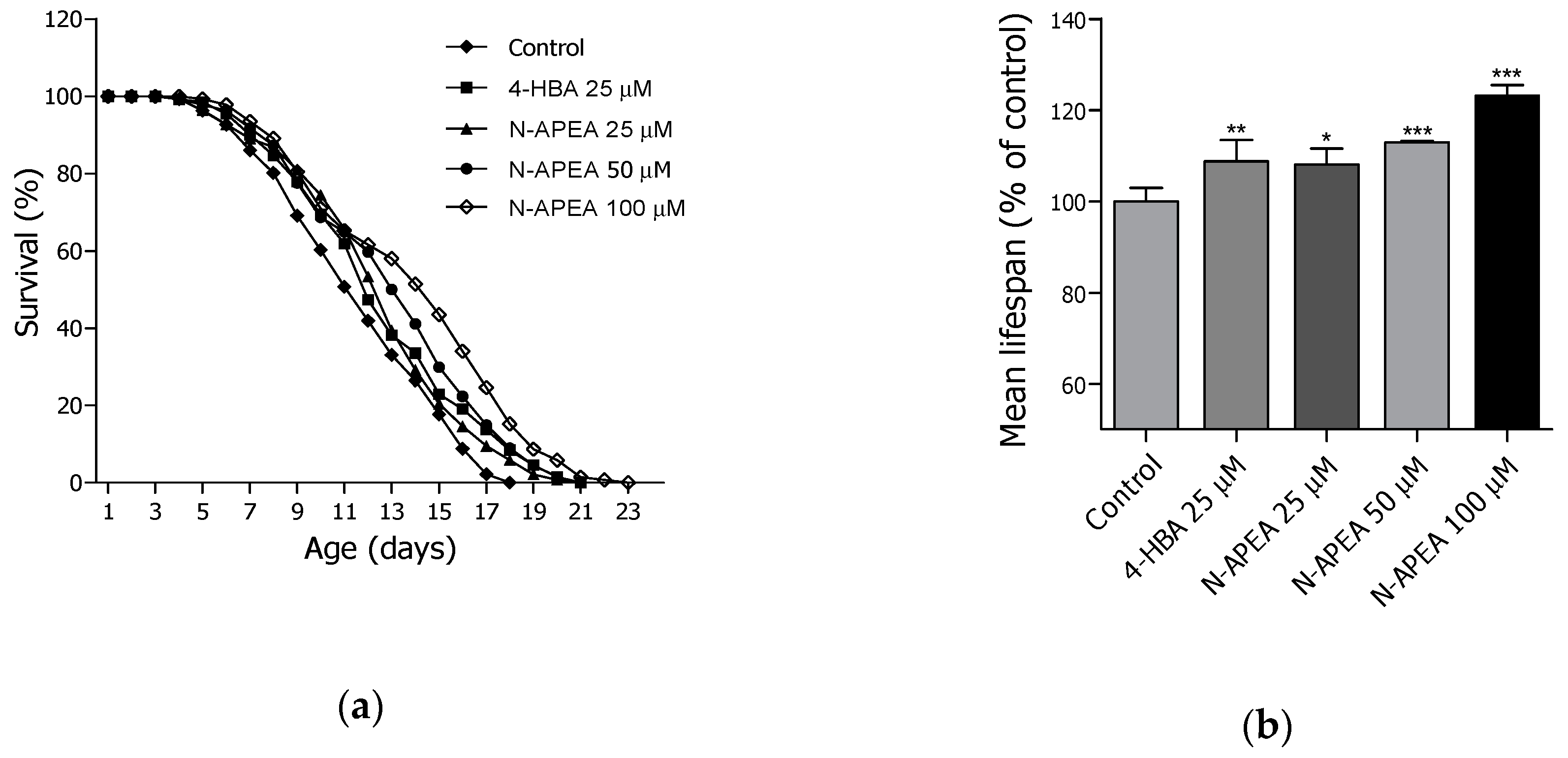
2.3. Lifespan Assay of PEA and N-APEA
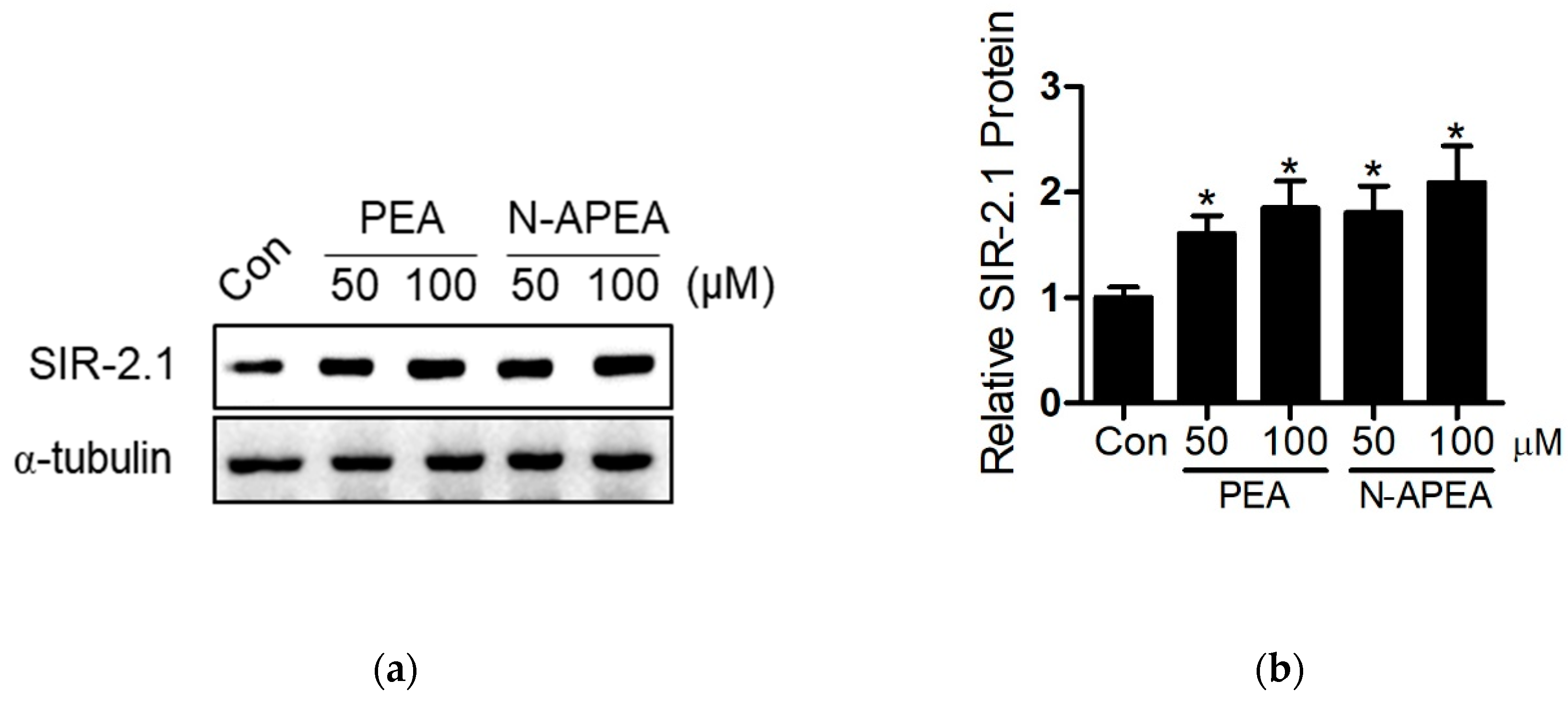
2.4. SIR-2.1 Protein
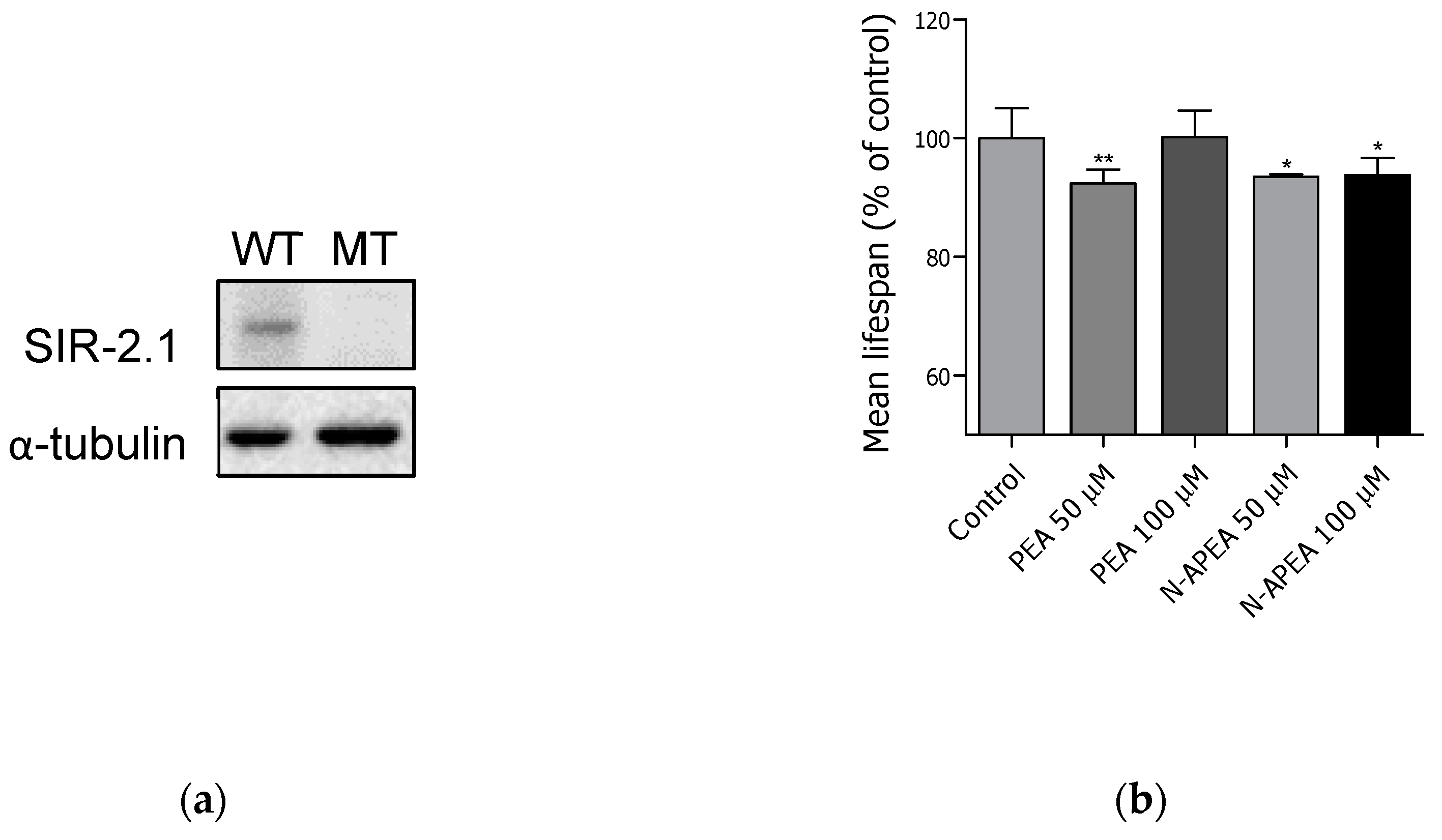
2.5. Lifespan Assay in VC199 sir-2.1(ok434)
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Bacterial Source and Extraction
4.3. Purification
4.4. Compounds Synthesis
4.5. C. Elegans strains and Maintenance
4.6. Lifespan Assay
4.7. Western Blot
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PEA | Phenethylamine |
| TA | Tyramine |
| TRA | Tryptamine |
| N-APEA | N-acetylphenethylamine |
| N-ATA | N-acetyltyramine |
| N-ATRA | N-acetyltryptamine |
| NMR | Nuclear magnetic resonance |
| BGCs | Biosynthetic gene clusters |
| NGM | Nematode growth medium |
References
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The human microbiota in health and disease. Engineering (Beijing) 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, S.; de Vos, W.M. How to manipulate the microbiota: Fecal microbiota transplantation. In Microbiota of the human body. Advances in experimental medicine and biology; Schwiertz, A., Ed.; Springer: Cham, Switzerland, 2016; Volume 902, pp. 143–153. [Google Scholar]
- Evrensel, A.; Ceylan, M.E. Fecal microbiota transplantation and its usage in neuropsychiatric disorders. Clin. Psychopharmacol. Neurosci. 2016, 14, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Xu, T.; Huang, G.; Jiang, S.; Gu, Y.; Chen, F. Oral microbiomes: More and more importance in oral cavity and whole body. Protein Cell 2018, 9, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I.; Yamazaki, K. Can oral bacteria affect the microbiome of the gut? J. Oral Microbiol. 2019, 11, 1586422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, L.; LoTempio, J.; Marquitz, A.; Daschner, P.; Xi, D.; Flores, R.; Brown, L.; Ranallo, R.; Maruvada, P.; Regan, K.; et al. A review of 10 years of human microbiome research activities at the US National Institutes of Health, Fiscal Years 2007–2016. Microbiome 2019, 7, 31. [Google Scholar]
- Aleman, F.D.D.; Valenzano, D.R. Microbiome evolution during host aging. PLoS Pathog. 2019, 15, e1007727. [Google Scholar] [CrossRef] [PubMed]
- Galkin, F.; Aliper, A.; Putin, E.; Kuznetsov, I.; Gladyshev, V.N.; Zhavoronkov, A. Human microbiome aging clocks based on deep learning and tandem of permutation feature importance and accumulated local effects. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Dinakaran, V.; Mandape, S.N.; Shuba, K.; Pratap, S.; Sakhare, S.S.; Tabatabai, M.A.; Smoot, D.T.; Farmer-Dixon, C.M.; Kesavalu, L.N.; Adunyah, S.E.; et al. Identification of specific oral and gut pathogens in full thickness colon of colitis patients: Implications for colon motility. Front. Microbiol. 2019, 9, 3220. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.S.; Cimermancic, P.; Schulze, C.J.; Brown, L.C.W.; Martin, J.; Mitreva, M.; Clardy, J.; Linington, R.G.; Fischbach, M.A. A systematic analysis of biosynthetic gene clusters in the human microbiome reveals a common family of antibiotics. Cell 2014, 158, 1402–1414. [Google Scholar] [CrossRef] [Green Version]
- Blum, H.E. The human microbiome. Adv. Med. Sci. 2017, 62, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Gems, D. Mechanisms of aging: Public or private. Genetics 2002, 3, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Weinert, B.T.; Timiras, P.S. Invited Review: Theories of aging. J. Appl. Physiol. 2003, 95, 1706–1716. [Google Scholar] [CrossRef]
- North, B.J.; Verdin, E. Sirtuins: Sir2-related NAD-dependent protein deacetylases. Genome Biol. 2004, 5, 224. [Google Scholar] [CrossRef] [Green Version]
- Blander, G.; Guarente, L. The Sir2 family of protein deacetylases. Annu. Rev. Biochem. 2004, 73, 417–735. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Cruzat, V.F.; Newshome, P.; Cheng, J.; Chen, Y.; Lu, Y. Regulation of SIRT1 in aging: Roles in mitochondrial function and biogenesis. Mech. Ageing Dev. 2016, 155, 10–21. [Google Scholar] [CrossRef]
- Greiss, S.; Hall, J.; Ahmed, S.; Gartner, A.C. elegans SIR-2.1 translocation is linked to a proapoptotic pathway parallel to cep-1/p53 during DNA damage-induced apoptosis. Genes Dev. 2008, 22, 2831–2842. [Google Scholar] [CrossRef] [Green Version]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kwon, G.; Park, J.; Kim, J.-K.; Lim, Y.-H. SIR-2.1-dependent lifespan extension of Caenorhabditis elegans by oxyresveratrol and resveratrol. Exp. Biol. Med. (Maywood) 2016, 241, 1757–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.; Jeon, H.; Cha, D.S. 4-Hydroxybenzoic acid-mediated lifespan extension in Caenorhabditis elegans. J. Funct. Foods 2014, 7, 630–640. [Google Scholar] [CrossRef]
- Lee, E.B.; Kim, J.H.; Kim, Y.J.; Noh, Y.J.; Kim, S.J.; Hwang, I.H.; Kim, D.K. Lifespan-extending property of 6-shogaol from Zingiber officinale Roscoe in Caenorhabditis elegans. Arch. Pharm. Res. 2018, 41, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Garcia, L.R. SIR-2.1 integrates metabolic homeostasis with the reproductive neuromuscular excitability in early aging male Caenorhabditis elegans. Elife 2014, 3, e01730. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut microbiota-produced tryptamine activates an epithelial G-protein-coupled receptor to increase colonic secretion. Cell Host Microbe 2018, 23, 775–785. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.A.; Macreadie, I.G. Inhibition of respiration in yeast by 2-phenylethylamine. Curr. Bioact. Compd. 2018, 14, 67–69. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Park, S.J.; Jeong, D.K.; Kim, K.-W.; Lim, S.-H.; Lee, S.-H.; Choe, S.-J.; Chang, Y.-H.; Park, I.; Kook, J.-K. Isolation and characterization of the mutans streptococci from the dental plaques in Koreans. J. Microbiol. 2007, 45, 246–255. [Google Scholar]
- Hu, Y.; Legako, A.G.; Espindola, A.P.D.M.; MacMillan, J.B. Erythrolic acids A−E, meroterpenoids from a marine-derived Erythrobacter sp. J. Org. Chem. 2012, 77, 3401–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbu, N.; Nagarjun, N.; Jacob, M.; Kalaiarasi, J.M.V.K.; Dhakshinamoorthy, A. Acetylation of alcohols, amines, phenols, thiols under catalyst and solvent-free conditions. Chemistry 2019, 1, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. In C. elegans: A Practical Approach; Hope, I.A., Ed.; Oxford University Press: New York, NY, USA, 1999; pp. 51–67. [Google Scholar]
- Amrit, F.R.G.; Ratnappan, R.; Keith, S.A.; Ghazi, A. The C. elegans lifespan assay toolkit. Methods 2014, 68, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.E.; Lee, Y.; Lee, S.-J.V. Western blot analysis of C. elegans proteins. In Hypoxia. Methods in Molecular Biology; Huang, L., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1742, pp. 213–225. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dose (µM) | Mean Lifespan1 (Day) | Maximum Lifespan (day) | Change in Mean Lifespan2 (%) | Log-Rank Test |
|---|---|---|---|---|---|
| Control | - | 11.1 ± 0.3 | 18 | / | / |
| 4-HBA | 25 | 12.7 ± 0.3 | 21 | 13.8 | *** p < 0.001 |
| PEA | 25 | 12.5 ± 0.3 | 20 | 12.4 | *** p < 0.001 |
| 50 | 13.3 ± 0.3 | 21 | 19.8 | *** p < 0.001 | |
| 100 | 13.6 ± 0.3 | 22 | 21.6 | *** p < 0.001 |
| Treatment | Dose (µM) | Mean Lifespan1 (day) | Maximum Lifespan (day) | Change in Mean Lifespan (%) | Log-Rank Test |
|---|---|---|---|---|---|
| Control | - | 11.7 ± 0.3 | 18 | - | - |
| 4-HBA | 25 | 12.7 ± 0.3 | 21 | 8.3 | ** p < 0.01 |
| N-APEA | 25 | 12.6 ± 0.3 | 21 | 7.8 | * p < 0.05 |
| 50 | 13.2 ± 0.3 | 21 | 12.6 | *** p < 0.001 | |
| 100 | 14.0 ± 0.4 | 23 | 19.9 | *** p < 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Bang, I.H.; Noh, Y.J.; Kim, D.K.; Bae, E.J.; Hwang, I.H. Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression. Int. J. Mol. Sci. 2020, 21, 2212. https://doi.org/10.3390/ijms21062212
Kim JH, Bang IH, Noh YJ, Kim DK, Bae EJ, Hwang IH. Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression. International Journal of Molecular Sciences. 2020; 21(6):2212. https://doi.org/10.3390/ijms21062212
Chicago/Turabian StyleKim, Jun Hyeong, In Hyuk Bang, Yun Jeong Noh, Dae Keun Kim, Eun Ju Bae, and In Hyun Hwang. 2020. "Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression" International Journal of Molecular Sciences 21, no. 6: 2212. https://doi.org/10.3390/ijms21062212
APA StyleKim, J. H., Bang, I. H., Noh, Y. J., Kim, D. K., Bae, E. J., & Hwang, I. H. (2020). Metabolites Produced by the Oral Commensal Bacterium Corynebacterium durum Extend the Lifespan of Caenorhabditis elegans via SIR-2.1 Overexpression. International Journal of Molecular Sciences, 21(6), 2212. https://doi.org/10.3390/ijms21062212