Forkhead Box P3 Methylation and Expression in Men with Obstructive Sleep Apnea

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Subject Characteristics
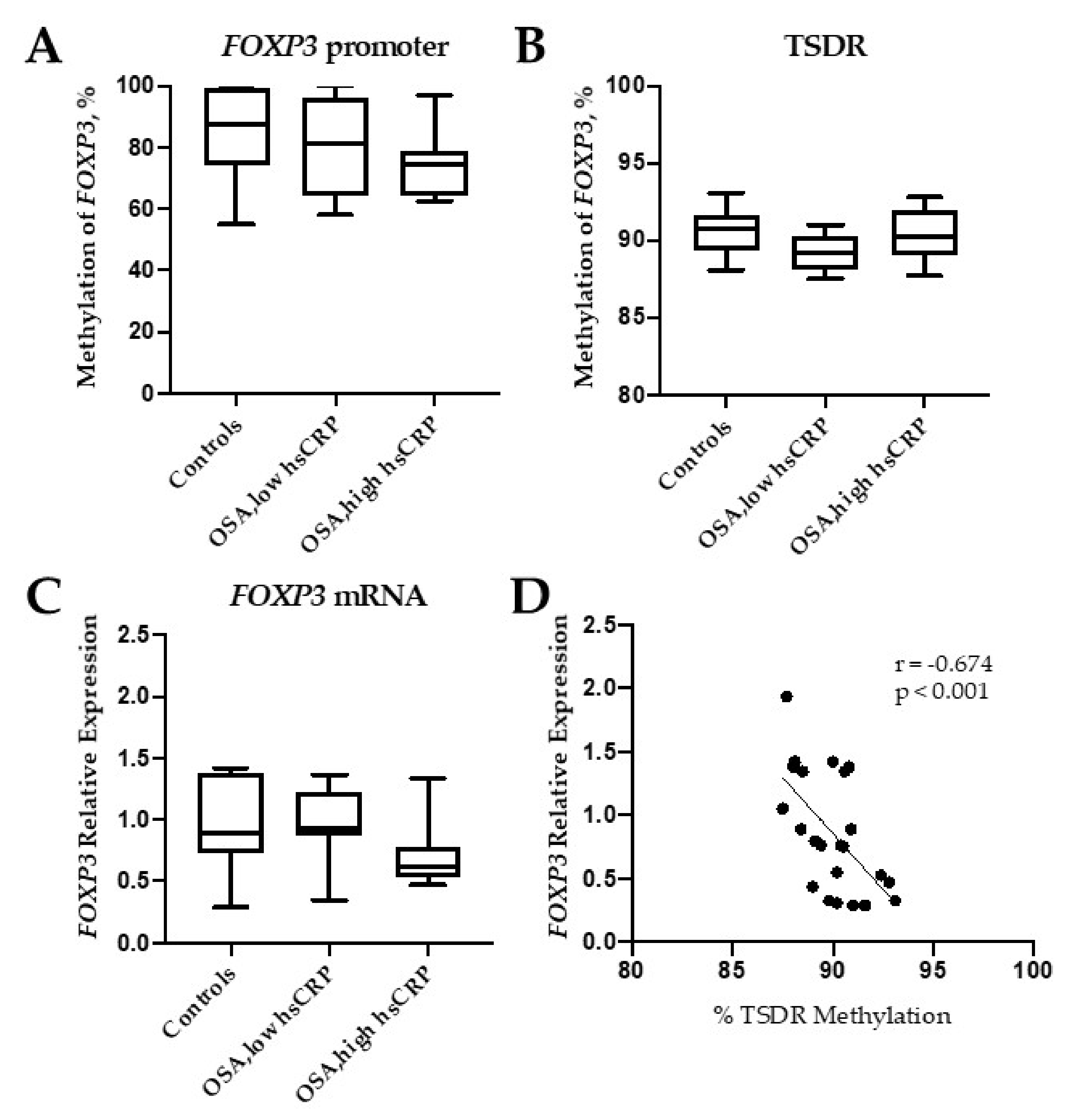
2.2. FOXP3 DNA Methylation Analysis
2.3. FOXP3 Protein and Gene Expression
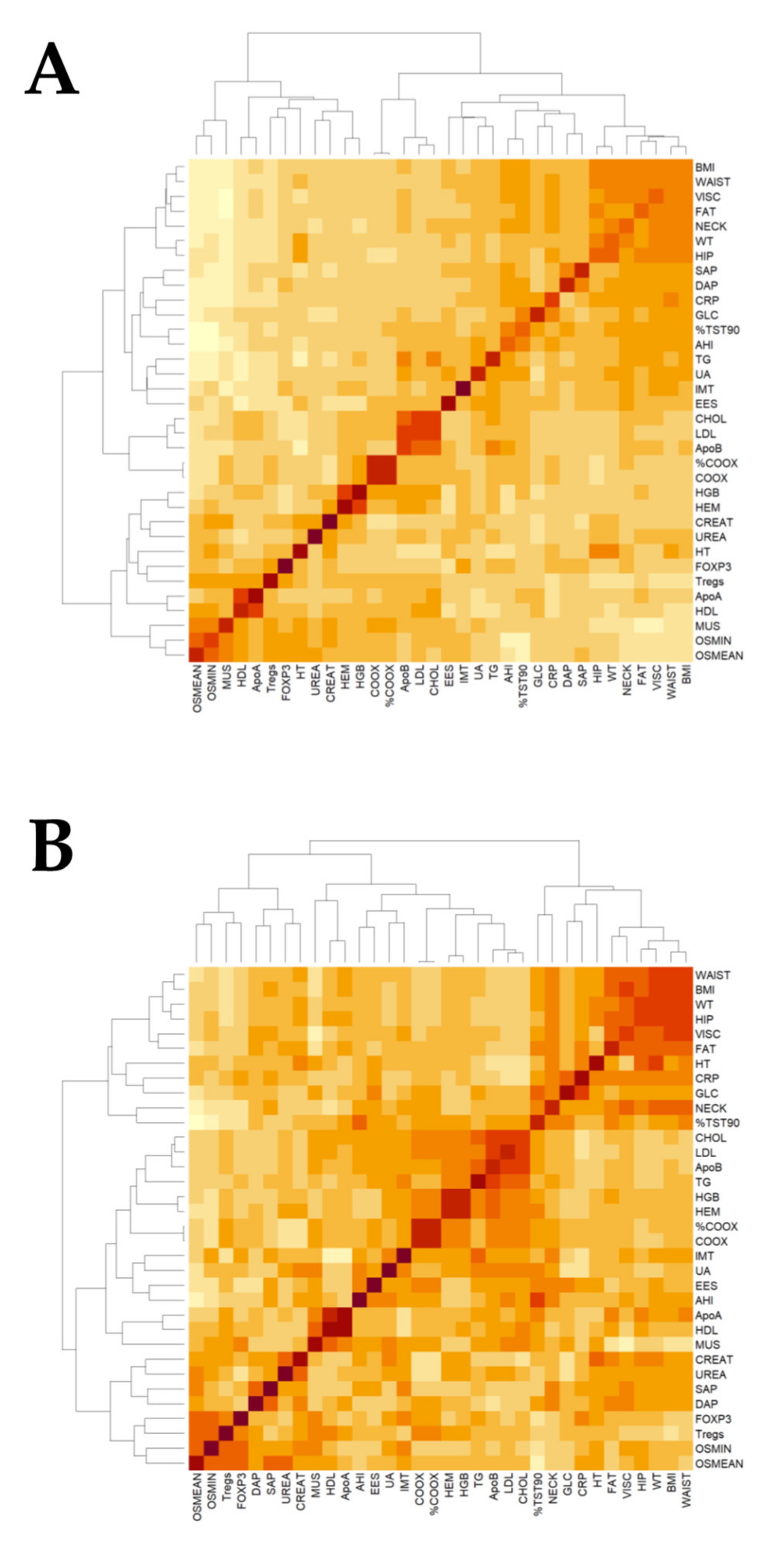
2.4. Association of FOXP3 mRNA Expression with Clinical Data
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Clinical Assessment
4.3. Flow Cytometry
4.4. FOXP3 DNA Methylation Analyses
4.5. FOXP3 Gene Expression
4.6. Quantification of FOXP3 Protein
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| OSA | Obstructive sleep apnea |
| FOXP3 | Forkhead Box P3 |
| AHI | Apnea-Hypopnea Index |
| IMT | Intima-media thickness of the common carotid artery |
References
- Young, T.; Peppard, P.E.; Gottlieb, D.J. Epidemiology of obstructive sleep apnea: A population health perspective. Am. J. Respir. Crit. Care Med. 2002, 165, 1217–1239. [Google Scholar] [CrossRef] [PubMed]
- Marin, J.M.; Carrizo, S.J.; Vicente, E.; Agusti, A.G. Long-term cardiovascular outcomes in men with obstructive sleep apnoea-hypopnoea with or without treatment with continuous positive airway pressure: An observational study. Lancet 2005, 365, 1046–1053. [Google Scholar] [CrossRef]
- Gottlieb, D.J.; Yenokyan, G.; Newman, A.B.; O’Connor, G.T.; Punjabi, N.M.; Quan, S.F.; Redline, S.; Resnick, H.E.; Tong, E.K.; Diener-West, M.; et al. Prospective study of obstructive sleep apnea and incident coronary heart disease and heart failure: The sleep heart health study. Circulation 2010, 122, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Yokoe, T.; Minoguchi, K.; Matsuo, H.; Oda, N.; Minoguchi, H.; Yoshino, G.; Hirano, T.; Adachi, M. Elevated levels of C-reactive protein and interleukin-6 in patients with obstructive sleep apnea syndrome are decreased by nasal continuous positive airway pressure. Circulation 2003, 107, 1129–1134. [Google Scholar] [CrossRef] [Green Version]
- Ikonomidis, I.; Stamatelopoulos, K.; Lekakis, J.; Vamvakou, G.D.; Kremastinos, D.T. Inflammatory and non-invasive vascular markers: The multimarker approach for risk stratification in coronary artery disease. Atherosclerosis 2008, 199, 3–11. [Google Scholar] [CrossRef]
- Watson, J.A.; Watson, C.J.; Mccann, A.; Baugh, J. Epigenetics, the epicenter of the hypoxic response. Epigenetics 2010, 5, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Stenvinkel, P.; Karimi, M.; Johansson, S.; Axelsson, J.; Suliman, M.; Lindholm, B.; Heimbürger, O.; Barany, P.; Alvestrand, A.; Nordfors, L.; et al. Impact of inflammation on epigenetic DNA methylation - A novel risk factor for cardiovascular disease? J. Intern. Med. 2007, 261, 488–499. [Google Scholar] [CrossRef]
- Klingenberg, R.; Gerdes, N.; Badeau, R.M.; Gisterå, A.; Strodthoff, D.; Ketelhuth, D.F.J.; Lundberg, A.M.; Rudling, M.; Nilsson, S.K.; Olivecrona, G.; et al. Depletion of FOXP3+ regulatory T cells promotes hypercholesterolemia and atherosclerosis. J. Clin. Invest. 2013, 123, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bhattacharjee, R.; Khalyfa, A.; Kheirandish-Gozal, L.; Capdevila, O.S.; Wang, Y.; Gozal, D. DNA Methylation in Inflammatory Genes among Children with Obstructive Sleep Apnea. Am. J. Respir. Crit. Care Med. 2012, 185, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Liu, H.; Zhang, G.; Li, P.; Wang, Z.; Huang, S.; Yang, Q.; Li, Y. The treg/th17 imbalance in patients with obstructive sleep apnoea syndrome. Mediators Inflamm. 2012, 2012, 815308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Chen, T.-W.; Su, M.-C.; Chen, C.-J.; Chen, K.-D.; Liou, C.-W.; Tang, P.; Wang, T.-Y.; Chang, J.-C.; Wang, C.-C.; et al. Whole Genome DNA Methylation Analysis of Obstructive Sleep Apnea: IL1R2,NPR2, AR, SP140 Methylation and Clinical Phenotype. Sleep 2016, 39, 743–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, J.M.; Artal, J.; Martin, T.; Carrizo, S.J.; Andres, M.; Martin-Burriel, I.; Bolea, R.; Sanz, A.; Varona, L.; Godino, J.; et al. Epigenetics modifications and Subclinical Atherosclerosis in Obstructive Sleep Apnea: The EPIOSA study. BMC Pulm. Med. 2014, 14, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bégin, P.; Schulze, J.; Baron, U.; Olek, S.; Bauer, R.N.; Passerini, L.; Baccheta, R.; Nadeau, K.C. Human in vitro induced T regulatory cells and memory T cells share common demethylation of specific FOXP3 promoter region. Clin. Transl. Allergy 2015, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, K.; McDonald-Hyman, C.; Noth, E.M.; Pratt, B.; Hammond, S.K.; Balmes, J.; Tager, I. Ambient air pollution impairs regulatory T-cell function in asthma. J. Allergy Clin. Immunol. 2010, 126, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Brunst, K.J.; Leung, Y.-K.; Ryan, P.H.; Khurana Hershey, G.K.; Levin, L.; Ji, H.; LeMasters, G.K.; Ho, S.-M. Forkhead box protein 3 (FOXP3) hypermethylation is associated with diesel exhaust exposure and risk for childhood asthma. J. Allergy Clin. Immunol. 2013, 131, 592–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Wang, Q.; Sun, X.H.; Liu, R.Z.; Shu, Y.; Kanekura, T.; Huang, J.H.; Li, Y.P.; Wang, J.C.; Zhao, M.; et al. DNA hypermethylation of the forkhead box protein 3 (FOXP3) promoter in CD4+ T cells of patients with systemic sclerosis. Br. J. Dermatol. 2014, 171, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Garcia, M.A.; Lyu, S.C.; Bucayu, R.; Kohli, A.; Ishida, S.; Berglund, J.P.; Tsai, M.; Maecker, H.; O’Riordan, G.; et al. Peanut oral immunotherapy results in increased antigen-induced regulatory T-cell function and hypomethylation of forkhead box protein 3 (FOXP3). J. Allergy Clin. Immunol. 2014, 133, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.-L.; Gozal, D.; Wang, Y.; Bandla, H.P.R.; Bhattacharjee, R.; Kulkarni, R.; Kheirandish-Gozal, L. Alterations in Circulating T-Cell Lymphocyte Populations in Children with Obstructive Sleep Apnea. Sleep 2013, 36, 913–922. [Google Scholar] [CrossRef] [Green Version]
- O’Driscoll, D.M.; Landry, S.A.; Pham, J.; Young, A.; Sands, S.A.; Hamilton, G.S.; Edwards, B.A. The physiological phenotype of obstructive sleep apnea differs between Caucasian and Chinese patients. Sleep 2019, 42. [Google Scholar] [CrossRef]
- Young, T.; Skatrud, J.; Peppard, P.E. Risk Factors for Obstructive Sleep Apnea in Adults. JAMA 2004, 291, 2013. [Google Scholar] [CrossRef]
- Hochberg, Z.; Feil, R.; Constancia, M.; Fraga, M.; Junien, C.; Carel, J.C.; Boileau, P.; Le Bouc, Y.; Deal, C.L.; Lillycrop, K.; et al. Child health, developmental plasticity, and epigenetic programming. Endocr. Rev. 2011, 32, 159–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.A.; Chen, N.H.; Huang, C.G.; Lin, S.W.; Fang, T.J.; Li, H.Y. Patients with severe obstructive sleep apnea syndrome and elevated high-sensitivity C-reactive protein need priority treatment. Otolaryngol. Head Neck Surg. 2010, 143, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.E.; Lu, A.T.; Chen, B.H.; Hernandez, D.G.; Singleton, A.B.; Ferrucci, L.; Bandinelli, S.; Salfati, E.; Manson, J.A.E.; Quach, A.; et al. Menopause accelerates biological aging. Proc. Natl. Acad. Sci. USA 2016, 113, 9327–9332. [Google Scholar] [CrossRef] [Green Version]
- Bacon, E.R.; Mishra, A.; Wang, Y.; Desai, M.K.; Yin, F.; Brinton, R.D. Neuroendocrine aging precedes perimenopause and is regulated by DNA methylation. Neurobiol. Aging 2019, 74, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Voskuhl, R.R.; Sawalha, A.H.; Itoh, Y. Sex chromosome contributions to sex differences in multiple sclerosis susceptibility and progression. Mult. Scler. 2018, 24, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Oxygen homeostasis. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 336–361. [Google Scholar] [CrossRef] [PubMed]
- Clambey, E.T.; McNamee, E.N.; Westrich, J.A.; Glover, L.E.; Campbell, E.L.; Jedlicka, P.; De Zoeten, E.F.; Cambier, J.C.; Stenmark, K.R.; Colgan, S.P.; et al. Hypoxia-inducible factor-1 alpha-dependent induction of FoxP3 drives regulatory T-cell abundance and function during inflammatory hypoxia of the mucosa. Proc. Natl. Acad. Sci. USA 2012, 109, E2784–E2793. [Google Scholar] [CrossRef] [Green Version]
- Johns, M.W. A new method for measuring daytime sleepiness: The Epworth sleepiness scale. Sleep 1991, 14, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Berry, R.B.; Budhiraja, R.; Gottlieb, D.J.; Gozal, D.; Iber, C.; Kapur, V.K.; Marcus, C.L.; Mehra, R.; Parthasarathy, S.; Quan, S.F.; et al. Rules for scoring respiratory events in sleep: Update of the 2007 AASM manual for the scoring of sleep and associated events. J. Clin. Sleep Med. 2012, 8, 597–619. [Google Scholar] [CrossRef] [Green Version]
- Kapur, V.K.; Auckley, D.H.; Chowdhuri, S.; Kuhlmann, D.C.; Mehra, R.; Ramar, K.; Harrod, C.G. Clinical practice guideline for diagnostic testing for adult obstructive sleep apnea: An American academy of sleep medicine clinical practice guideline. J. Clin. Sleep Med. 2017, 13, 479–504. [Google Scholar] [CrossRef]
- Muntendam, P.; McCall, C.; Sanz, J.; Falk, E.; Fuster, V. High-Risk Plaque Initiative The BioImage Study: Novel approaches to risk assessment in the primary prevention of atherosclerotic cardiovascular disease—study design and objectives. Am. Heart J. 2010, 160, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Jarauta, E.; Mateo-Gallego, R.; Bea, A.; Burillo, E.; Calmarza, P.; Civeira, F. Carotid Intima-Media Thickness in Subjects With No Cardiovascular Risk Factors. Rev. Española Cardiol. (English Ed.) 2010, 63, 97–102. [Google Scholar] [CrossRef]
- Li, L.-C. Designing PCR Primer for DNA Methylation Mapping. In PCR Primer Design; Humana Press: Totowa, NJ, USA, 2007; pp. 370–383. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable | Controls | OSA | OSA (low hsCRP) | OSA (high hsCRP) |
|---|---|---|---|---|
| Individuals, n | 7 | 16 | 8 | 8 |
| Age (years) | 41.9 ± 6.9 | 43.2 ± 9.3 | 45.1 ± 9.6 | 42.4 ± 9.2 |
| BMI (kg/m2) | 29.8 ± 2.8 | 31.6 ± 4.0 | 30.7 ± 3.5 | 32.7 ± 4.4 |
| SBP (mmHg) | 123 ± 9 | 133 ± 15 * | 132 ± 17 | 133 ± 19 |
| DBP (mmHg) | 69 ± 9 | 82 ± 11 * | 81 ± 14 | 82 ± 13 |
| AHI (events/hour) | 2.2 ± 2.5 ** | 67.2 ± 20.3 | 65.1 ± 23.7 | 69.2 ± 17.6 |
| Nadir SaO2 | 89.6 ± 2.4 ** | 73.2 ± 8.1 | 71.4 ± 7.7 | 75.1 ± 8.5 |
| CT90, % | 0.1 ± 0.3 ** | 45.2 ± 24.7 | 43.2 ± 25.1 | 43.3 ± 25.7 |
| ESS | 9.7 ± 6.1 | 12.6 ± 3.5 | 12.7 ± 3.9 | 12.6 ± 2.8 |
| hsCRP (mg/dl) | 0.08 ± 0.09 * | 0.35 ± 0.25 | 0.14 ± 0.09 | 0.56 ± 0.17 *** |
| ApoA (mg/dl) | 140.9 ± 21.1 | 141.4 ± 20.3 | 140.2 ± 25.3 | 144.3 ± 25.3 |
| ApoB (mg/dl) | 96.9 ± 24.4 | 114.1 ± 29.3 | 112.3 ± 29.9 | 117.6 ± 32.2 |
| IMT (mm) | 0.54 ± 0.09 * | 0.66 ± 0.12 | 0.61 ± 0.09 | 0.70 ± 0.15 |
| Tregs (% CD4+ cells) | 6.7 ± 1.5 | 7.1 ± 1.5 | 6.5 ± 1.1 | 7.6 ± 1.7 |
| Variable | Controls | Mild-moderate OSA | Severe OSA |
|---|---|---|---|
| Individuals, n | 31 | 19 | 55 |
| Age (years) | 41.5 ± 8.4 | 45.3 ± 6.8 | 44.1±8.9 |
| BMI (kg/m2) | 27.2 ± 3.1 | 30.9 ± 4.6 | 32.2 ±5.2 *** |
| AHI (events/hour) | 2.0 ± 2.9 | 20.5 ± 6.5 * | 57.7 ± 19.9 *** § |
| ESS | 10.3 ± 4.7 | 8.8 ± 4.5 | 11.1 ± 4.6 |
| SBP (mmHg) | 122 ± 10 | 128 ± 10 | 132 ± 15 ** |
| DBP (mmHg) | 73 ± 10 | 80 ± 8* | 83 ± 11 ** |
| hsCRP (mg/dl) | 0.13 ± 0.12 | 0.26 ± 0.28 | 0.40 ± 0.34 * |
| ApoA (mg/dl) | 142.9 ± 22.1 | 152.2 ± 25.4 | 143.3 ± 22.5 |
| ApoB (mg/dl) | 98.8 ± 23.9 | 105.6 ± 27.4 | 117.8 ± 27.8 * |
| TREG | 7.1 ± 1.9 | 7.2 ± 1.5 | 7.1 ± 1.5 |
| IMT (mm) | 0.56 ± 0.09 | 0.61 ± 0.15 | 0.65 ± 0.13 * |
| Subclinical atherosclerosis, n (%) | 5 (16.1) | 4 (21.0) | 20 (36.3) **§ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanz-Rubio, D.; Sanz, A.; Varona, L.; Bolea, R.; Forner, M.; Gil, A.V.; Cubero, P.; Marin-Oto, M.; Martin-Burriel, I.; Marin, J.M. Forkhead Box P3 Methylation and Expression in Men with Obstructive Sleep Apnea. Int. J. Mol. Sci. 2020, 21, 2233. https://doi.org/10.3390/ijms21062233
Sanz-Rubio D, Sanz A, Varona L, Bolea R, Forner M, Gil AV, Cubero P, Marin-Oto M, Martin-Burriel I, Marin JM. Forkhead Box P3 Methylation and Expression in Men with Obstructive Sleep Apnea. International Journal of Molecular Sciences. 2020; 21(6):2233. https://doi.org/10.3390/ijms21062233
Chicago/Turabian StyleSanz-Rubio, David, Arianne Sanz, Luis Varona, Rosa Bolea, Marta Forner, Ana V. Gil, Pablo Cubero, Marta Marin-Oto, Inmaculada Martin-Burriel, and Jose M. Marin. 2020. "Forkhead Box P3 Methylation and Expression in Men with Obstructive Sleep Apnea" International Journal of Molecular Sciences 21, no. 6: 2233. https://doi.org/10.3390/ijms21062233
APA StyleSanz-Rubio, D., Sanz, A., Varona, L., Bolea, R., Forner, M., Gil, A. V., Cubero, P., Marin-Oto, M., Martin-Burriel, I., & Marin, J. M. (2020). Forkhead Box P3 Methylation and Expression in Men with Obstructive Sleep Apnea. International Journal of Molecular Sciences, 21(6), 2233. https://doi.org/10.3390/ijms21062233