Sustained Release of Decoy Wnt Receptor (sLRP6E1E2)-Expressing Adenovirus Using Gel-Encapsulation for Scar Remodeling in Pig Model

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
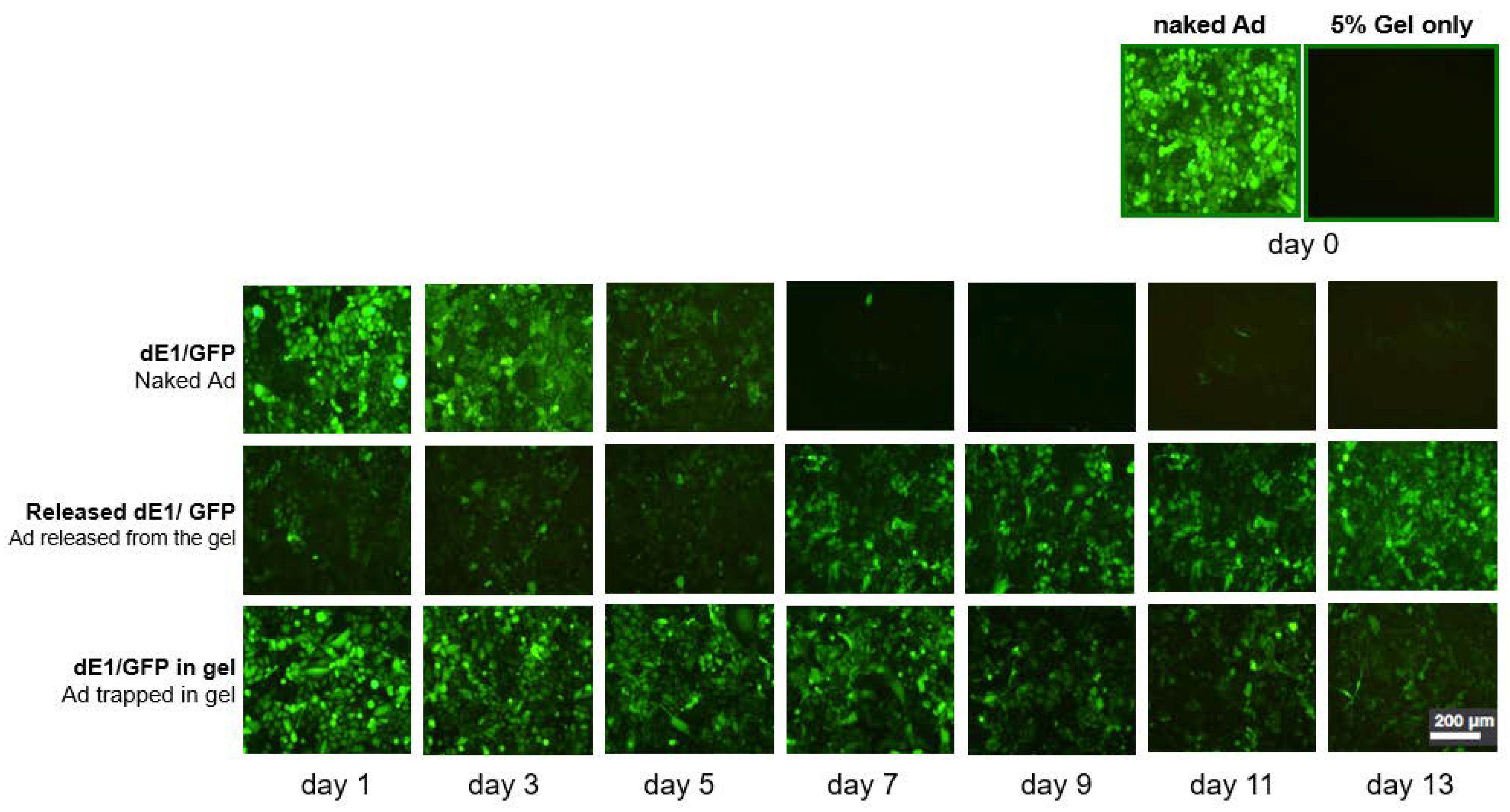
2.1. Ad Is Continuously Released from the Alginate Gel, and Gel Encapsulation Prolongs Ad Biological Activity
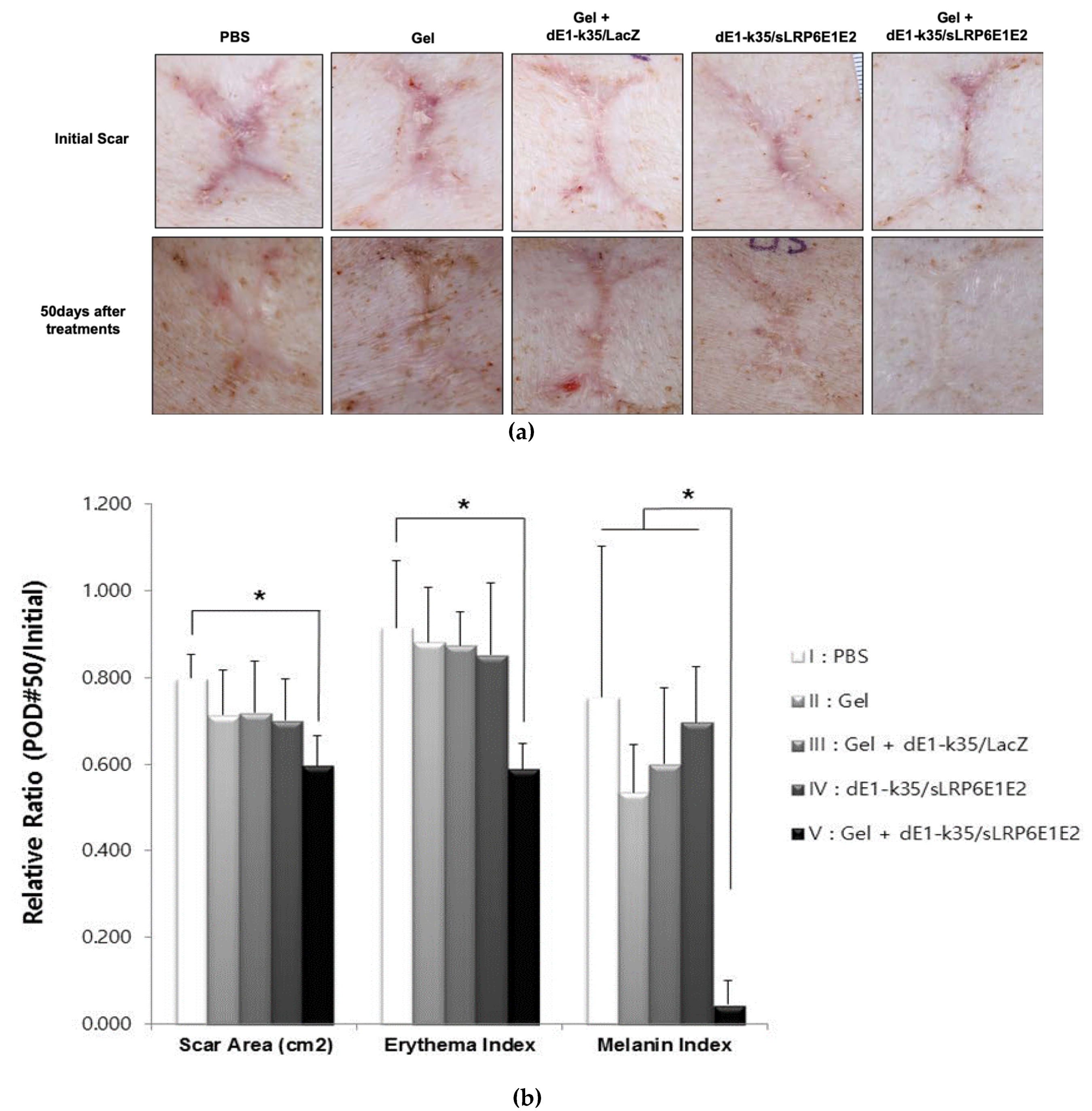
2.2. sLRP6E1E2-Expressing Ad/Gel Decreases scar Size and Color in Pig Scar Tissue
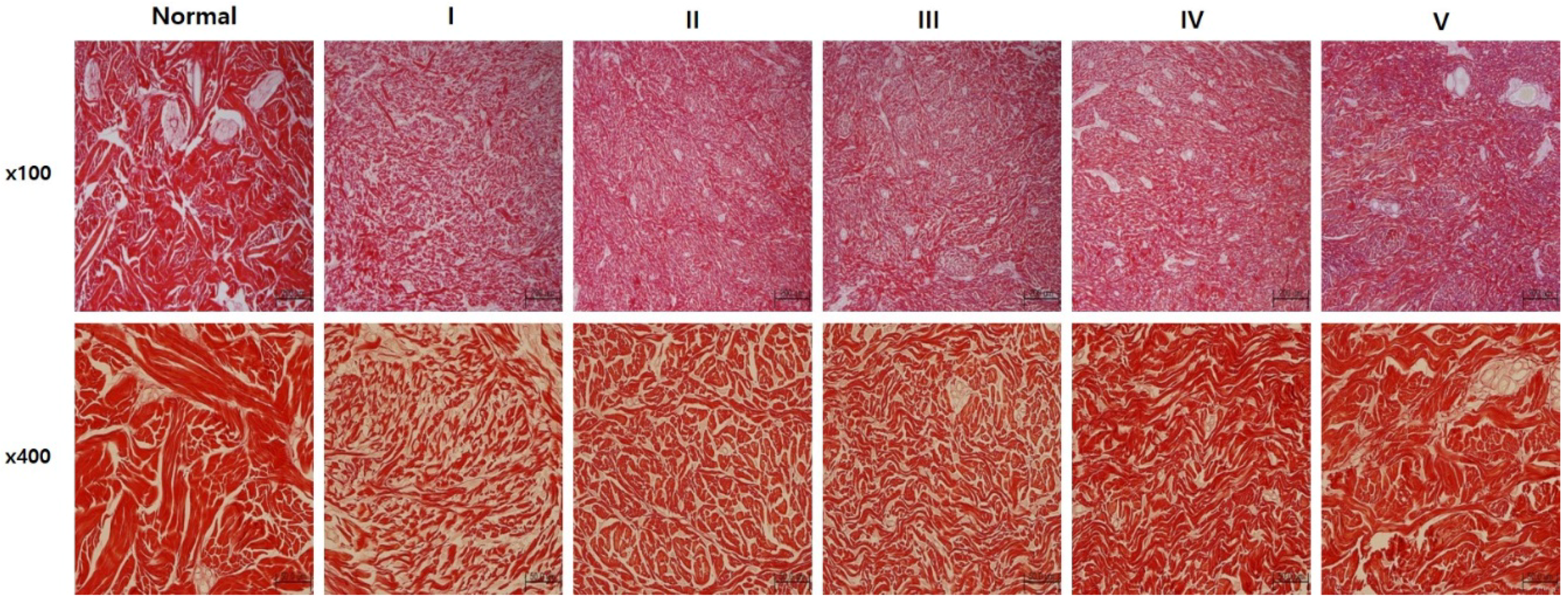
2.3. sLRP6E1E2-Expressing Ad/Gel Remodels Scar by Inducing Collagen Rearrangement
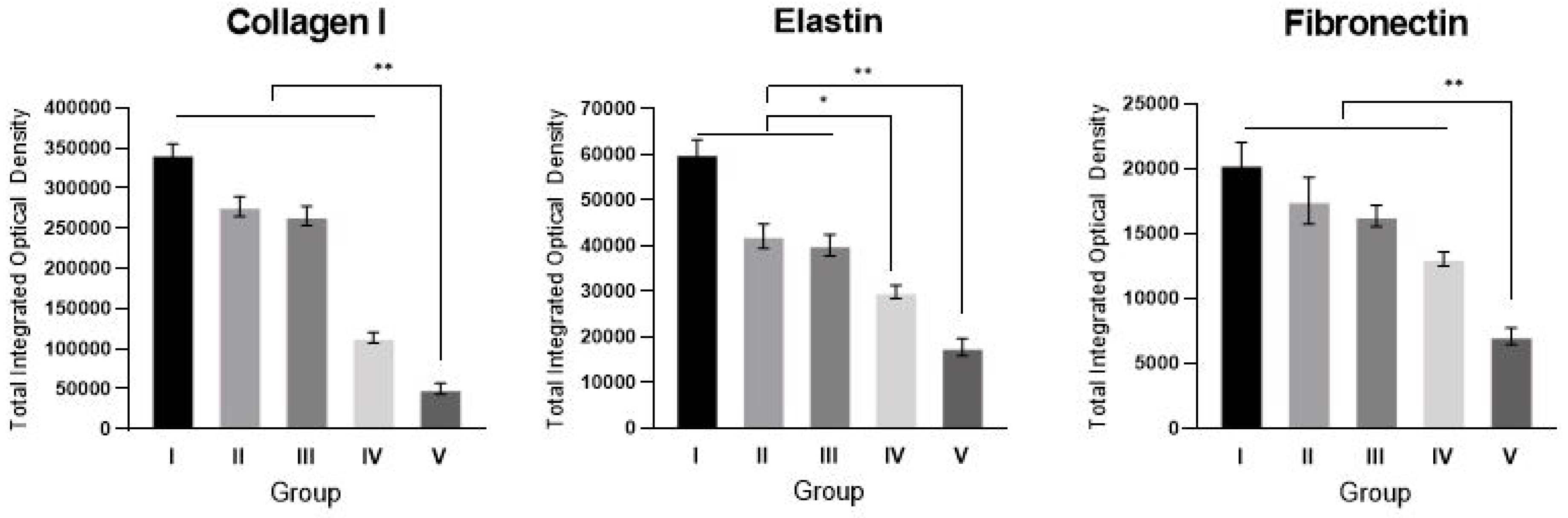
2.4. sLRP6E1E2-Expressing Ad/Gel Reduces Collagen I, Elastin, and Fibronectin Expression
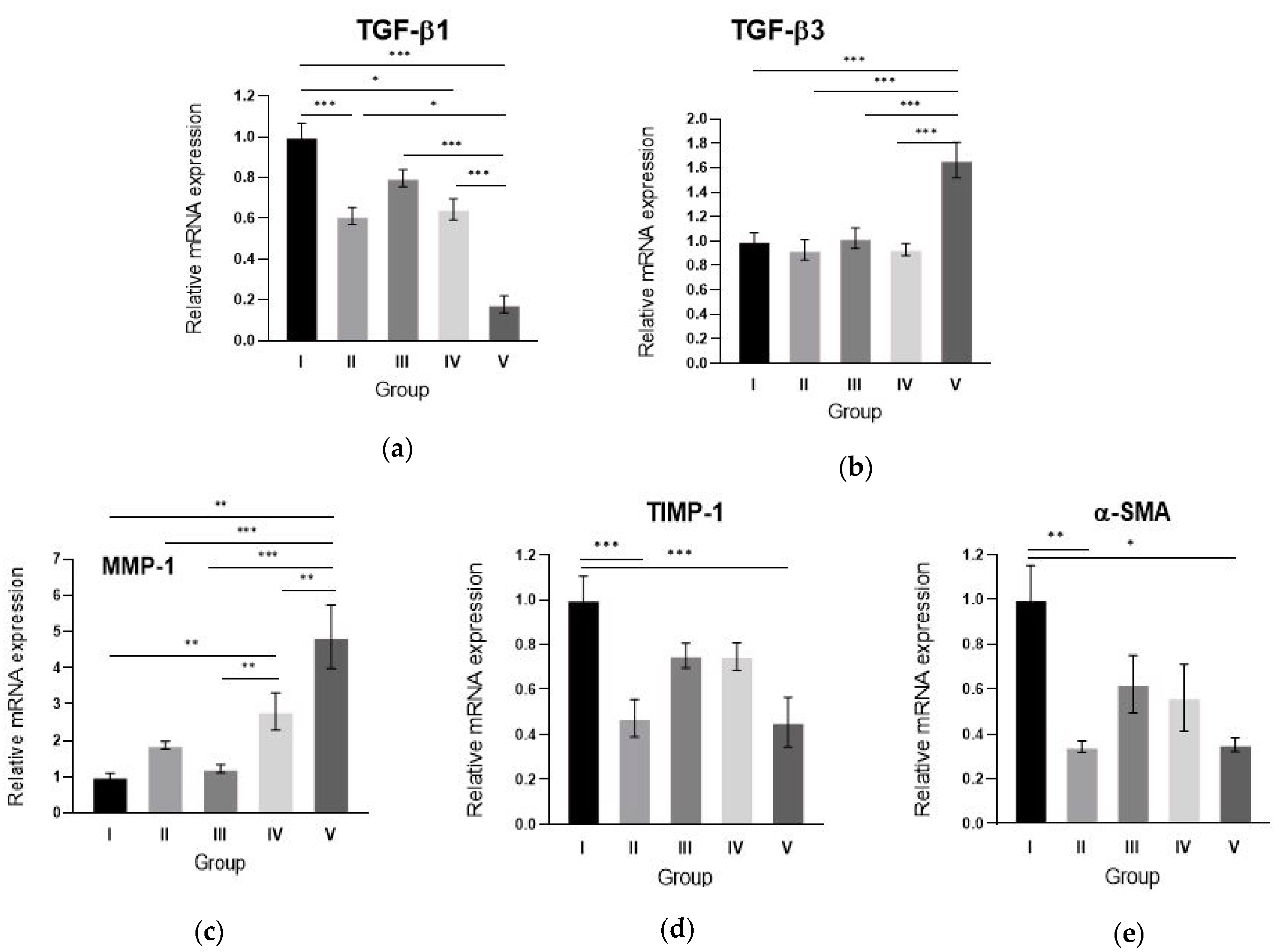
2.5. Alginate gel Encapsulating dE1-k35/sLRP6E1E2 Downregulates TGF-β1, Upregulates TGF-β3 mRNA Expression and Changes the Scar Remodeling Markers in Pig Scar Tissues
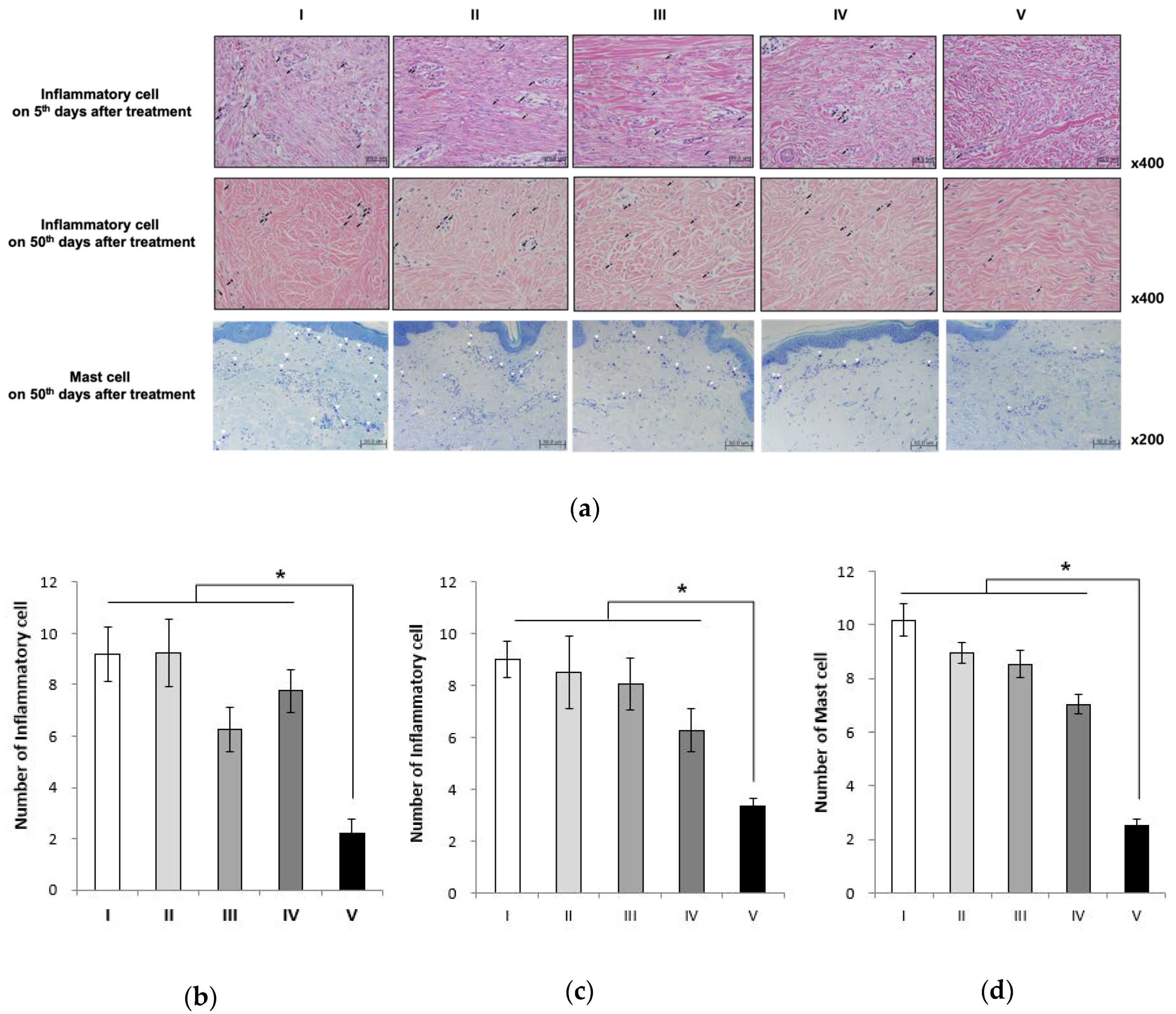
2.6. Alginate Gel Encapsulating dE1-k35/sLRP6E1E2 Decreases Inflammatory Cell Counts and Mast Cell Counts in Pig Scar Tissues
3. Discussion
4. Materials and Methods
4.1. Preparation of Ad and Alginate Gel
4.2. Biological Activity of Encapsulated Ad
4.3. Scar Formation
4.4. Injection of Ad
4.5. Therapeutic Evaluation of Scar Size and Erythema and Melanin Indices
4.6. Histology and Immunohistochemistry
4.7. Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ad | Adenoviral vector |
| ECM | Extracellular matrix |
| TGF-β | Transforming growth factor-β |
| PBS | Phosphate-buffered saline |
| GFP | Green Fluorescent Protein |
References
- Al-Attar, A.; Mess, S.; Thomassen, J.M.; Kauffman, C.L.; Davison, S.P. Keloid pathogenesis and treatment. Plast. Reconstr. Surg. 2006, 117, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Bran, G.M.; Goessler, U.R.; Hormann, K.; Riedel, F.; Sadick, H. Keloids: Current concepts of pathogenesis. Int. J. Mol. Med. 2009, 24, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.H.; Ogawa, R. Keloid research: Current status and future directions. Scars Burn Heal. 2019, 5, 2059513119868659. [Google Scholar] [CrossRef] [PubMed]
- Burd, A.; Huang, L. Hypertrophic response and keloid diathesis: Two very different forms of scar. Plast. Reconstr. Surg. 2005, 116, 150e–157e. [Google Scholar] [CrossRef]
- Atiyeh, B.S.; Costagliola, M.; Hayek, S.N. Keloid or hypertrophic scar: The controversy: Review of the literature. Ann. Plast. Surg. 2005, 54, 676–680. [Google Scholar] [CrossRef]
- Lee, W.J.; Lee, J.S.; Ahn, H.M.; Na, Y.; Yang, C.E.; Lee, J.H.; Hong, J.; Yun, C.O. Decoy wnt receptor (slrp6e1e2)-expressing adenovirus induces anti-fibrotic effect via inhibition of wnt and tgf-beta signaling. Sci. Rep. 2017, 7, 15070. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Kang, E.; Kwon, O.J.; Yun, T.J.; Park, H.K.; Kim, P.H.; Kim, S.W.; Kim, J.H.; Yun, C.O. Local sustained delivery of oncolytic adenovirus with injectable alginate gel for cancer virotherapy. Gene Ther. 2013, 20, 880–892. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, P.H.; Hwang, T.; Kwon, O.J.; Park, T.J.; Choi, S.W.; Yun, C.O.; Kim, J.H. Fabrication of cross-linked alginate beads using electrospraying for adenovirus delivery. Int. J. Pharm. 2012, 427, 417–425. [Google Scholar] [CrossRef]
- Koutsopoulos, S.; Unsworth, L.D.; Nagai, Y.; Zhang, S. Controlled release of functional proteins through designer self-assembling peptide nanofiber hydrogel scaffold. Proc. Natl. Acad. Sci. USA 2009, 106, 4623–4628. [Google Scholar] [CrossRef] [Green Version]
- Yun, I.S.; Kang, E.; Ahn, H.M.; Kim, Y.O.; Rah, D.K.; Roh, T.S.; Lee, W.J.; Yun, C.O. Effect of relaxin expression from an alginate gel-encapsulated adenovirus on scar remodeling in a pig model. Yonsei Med. J. 2019, 60, 854–863. [Google Scholar] [CrossRef]
- Young, W.G.; Worsham, M.J.; Joseph, C.L.; Divine, G.W.; Jones, L.R. Incidence of keloid and risk factors following head and neck surgery. JAMA Facial Plast. Surg. 2014, 16, 379–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ud-Din, S.; Bayat, A. New insights on keloids, hypertrophic scars, and striae. Dermatol. Clin. 2014, 32, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Shih, B.; Garside, E.; McGrouther, D.A.; Bayat, A. Molecular dissection of abnormal wound healing processes resulting in keloid disease. Wound Repair Regen. 2010, 18, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Drugging wnt signalling in cancer. EMBO J. 2012, 31, 2737–2746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, S.B.; Russell, J.D.; Trupin, K.M.; Gayden, A.E.; Opalenik, S.R.; Nanney, L.B.; Broquist, A.H.; Raju, L.; Williams, S.M. Epigenetically altered wound healing in keloid fibroblasts. J. Investig. Dermatol. 2010, 130, 2489–2496. [Google Scholar] [CrossRef] [Green Version]
- Bastakoty, D.; Young, P.P. Wnt/beta-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. Faseb J. 2016, 30, 3271–3284. [Google Scholar] [CrossRef]
- Lee, W.J.; Choi, I.K.; Lee, J.H.; Kim, Y.O.; Yun, C.O. A novel three-dimensional model system for keloid study: Organotypic multicellular scar model. Wound Repair Regen. 2013, 21, 155–165. [Google Scholar] [CrossRef]
- Carre, A.L.; James, A.W.; MacLeod, L.; Kong, W.; Kawai, K.; Longaker, M.T.; Lorenz, H.P. Interaction of wingless protein (wnt), transforming growth factor-beta1, and hyaluronan production in fetal and postnatal fibroblasts. Plast. Reconstr. Surg. 2010, 125, 74–88. [Google Scholar] [CrossRef]
- Sato, M. Upregulation of the wnt/beta-catenin pathway induced by transforming growth factor-beta in hypertrophic scars and keloids. Acta Dermato-Venereol. 2006, 86, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Xiao, L.; Sun, L.; Liu, F. Wnt/beta-catenin signaling: A promising new target for fibrosis diseases. Physiol. Res. 2012, 61, 337–346. [Google Scholar]
- Satterwhite, D.J.; Neufeld, K.L. Tgf-beta targets the wnt pathway components, apc and beta-catenin, as mv1lu cells undergo cell cycle arrest. Cell Cycle (Georgetown, Tex.) 2004, 3, 1069–1073. [Google Scholar] [CrossRef] [Green Version]
- Logan, C.Y.; Nusse, R. The wnt signaling pathway in development and disease. Annu. Rev. Cell Dev. Biol. 2004, 20, 781–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, S.; Poon, R.; Yu, C.; Khoury, M.; Shenker, R.; Fish, J.; Alman, B.A. Prolonged beta-catenin stabilization and tcf-dependent transcriptional activation in hyperplastic cutaneous wounds. Lab. Investig. 2005, 85, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Ries, C. Cytokine functions of timp-1. Cell Mol. Life Sci. 2014, 71, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Roldan, V.; Garcia, M.; Monmeneu, J.; de Burgos, F.G.; Lip, G.Y.; Marin, F. Matrix metalloproteinase-1 and its inhibitor, timp-1, in systolic heart failure: Relation to functional data and prognosis. J. Intern. Med. 2007, 262, 385–392. [Google Scholar] [CrossRef]
- Ogawa, R. Keloid and hypertrophic scars are the result of chronic inflammation in the reticular dermis. Int. J. Mol. Sci. 2017, 18, 606. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Hur, M.W.; Lee, S.K.; Choi, W.I.; Kwon, Y.G.; Yun, C.O. A novel slrp6e1e2 inhibits canonical wnt signaling, epithelial-to-mesenchymal transition, and induces mitochondria-dependent apoptosis in lung cancer. PLoS ONE 2012, 7, e36520. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.J.; Zhang, S.N.; Choi, I.K.; Kim, J.S.; Yun, C.O. Strengthening of antitumor immune memory and prevention of thymic atrophy mediated by adenovirus expressing il-12 and gm-csf. Gene Ther. 2012, 19, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.N.; Choi, I.K.; Huang, J.H.; Yoo, J.Y.; Choi, K.J.; Yun, C.O. Optimizing dc vaccination by combination with oncolytic adenovirus coexpressing il-12 and gm-csf. Mol. Ther. 2011, 19, 1558–1568. [Google Scholar] [CrossRef]
- Yun, I.S.; Lee, W.J.; Rah, D.K.; Kim, Y.O.; Park, B.Y. Skin color analysis using a spectrophotometer in asians. Skin Res. Technol. 2010, 16, 311–315. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-E.; Choi, S.; Lee, J.H.; Kang, E.H.; Ahn, H.M.; Roh, T.S.; Yun, C.-O.; Lee, W.J. Sustained Release of Decoy Wnt Receptor (sLRP6E1E2)-Expressing Adenovirus Using Gel-Encapsulation for Scar Remodeling in Pig Model. Int. J. Mol. Sci. 2020, 21, 2242. https://doi.org/10.3390/ijms21062242
Yang C-E, Choi S, Lee JH, Kang EH, Ahn HM, Roh TS, Yun C-O, Lee WJ. Sustained Release of Decoy Wnt Receptor (sLRP6E1E2)-Expressing Adenovirus Using Gel-Encapsulation for Scar Remodeling in Pig Model. International Journal of Molecular Sciences. 2020; 21(6):2242. https://doi.org/10.3390/ijms21062242
Chicago/Turabian StyleYang, Chae-Eun, Sewoon Choi, Ju Hee Lee, Eun Hye Kang, Hyo Min Ahn, Tai Suk Roh, Chae-Ok Yun, and Won Jai Lee. 2020. "Sustained Release of Decoy Wnt Receptor (sLRP6E1E2)-Expressing Adenovirus Using Gel-Encapsulation for Scar Remodeling in Pig Model" International Journal of Molecular Sciences 21, no. 6: 2242. https://doi.org/10.3390/ijms21062242
APA StyleYang, C. -E., Choi, S., Lee, J. H., Kang, E. H., Ahn, H. M., Roh, T. S., Yun, C. -O., & Lee, W. J. (2020). Sustained Release of Decoy Wnt Receptor (sLRP6E1E2)-Expressing Adenovirus Using Gel-Encapsulation for Scar Remodeling in Pig Model. International Journal of Molecular Sciences, 21(6), 2242. https://doi.org/10.3390/ijms21062242