Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages

 , , , , , , , and
, , , , , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
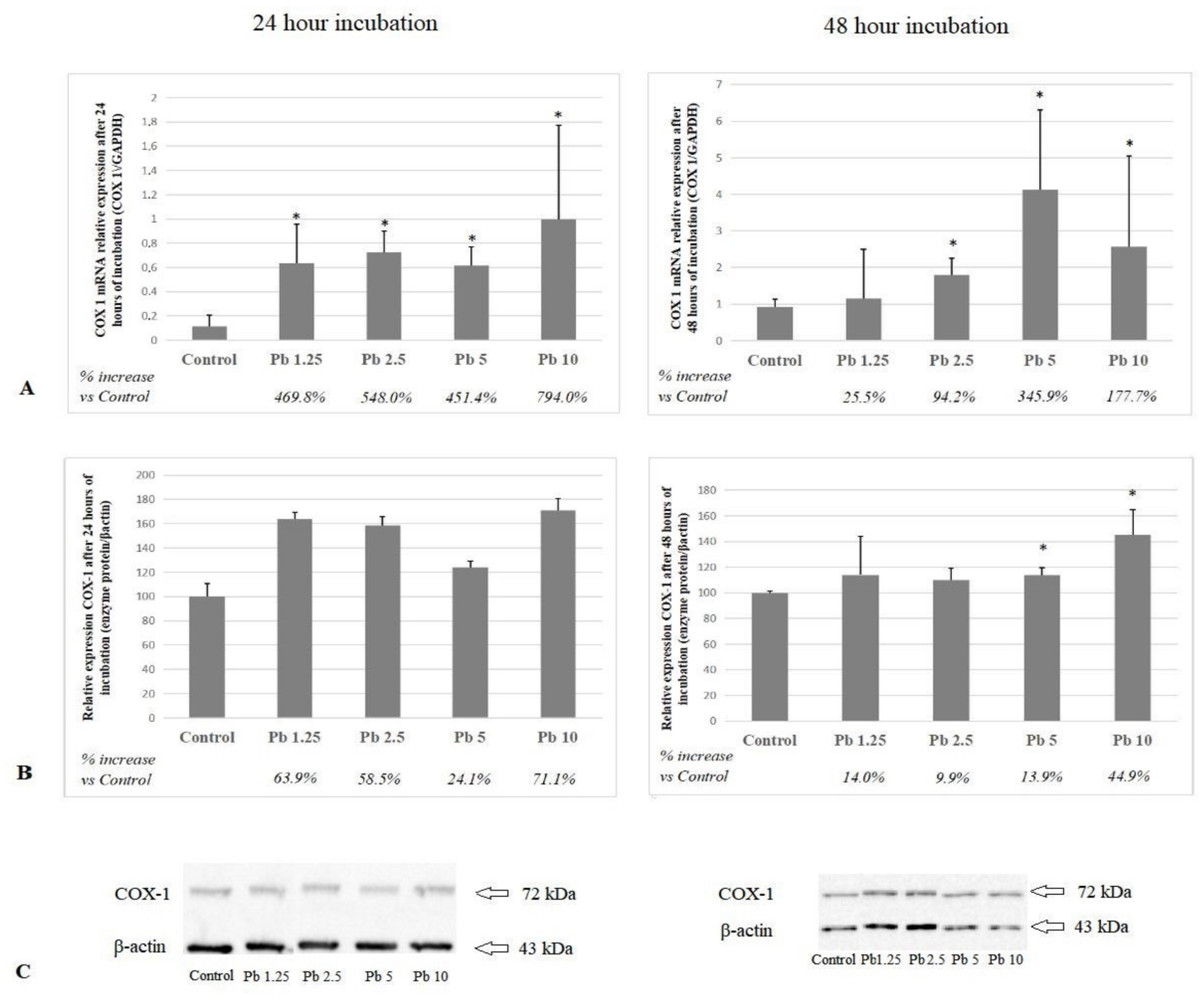
2.1. Cyclooxygenase 1
2.2. Cyclooxygenase 2
2.3. Thromboxane A2 and Prostaglandin E2
2.4. Interleukin 1β and Interleukin 6
3. Discussion
4. Materials and Methods
4.1. Materials
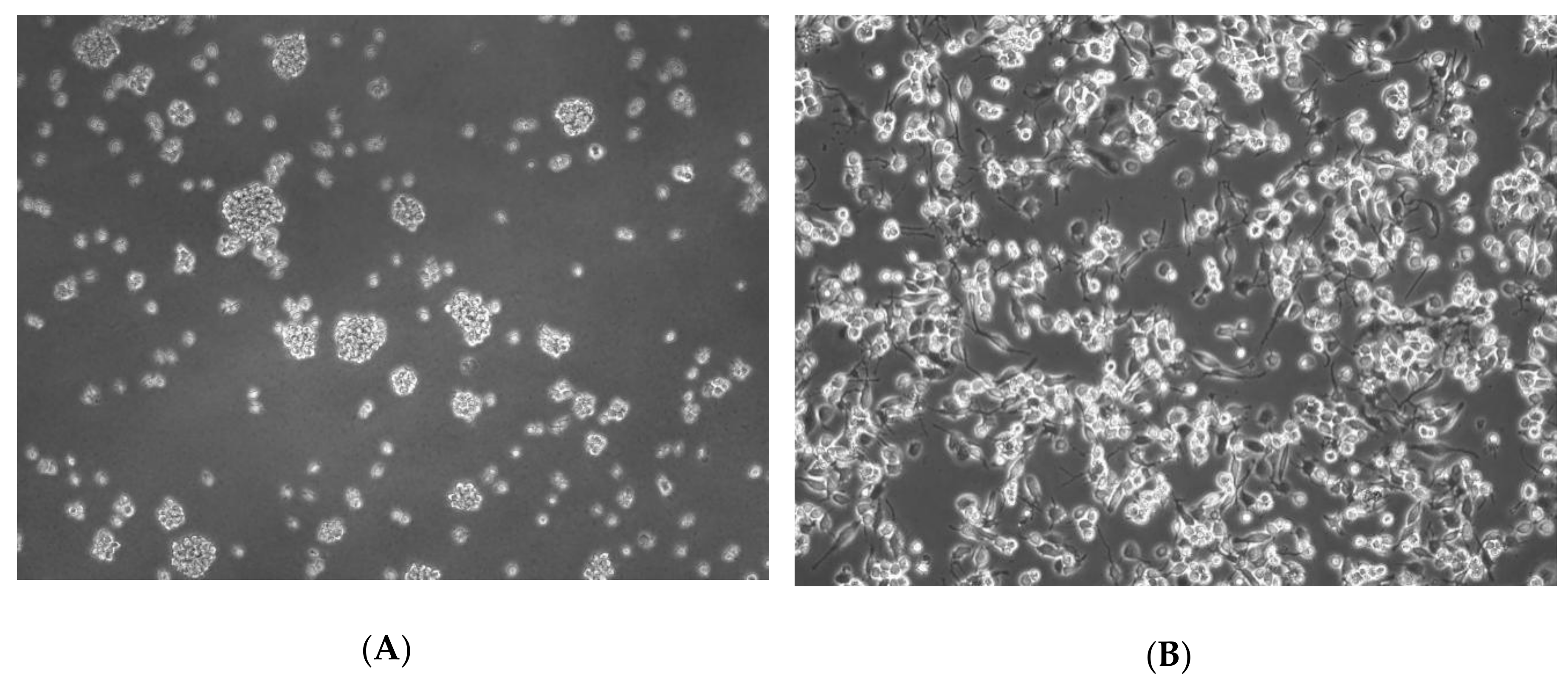
4.2. THP-1 Macrophages Experimental Model
4.3. Cell Culture and Treatment
4.4. Gene Expression
Cyclooxygenase-1 (COX-1) and Cyclooxygenase-2 (COX-2) Gene Expression Analysis by qRT-PCR
4.5. Protein Expression
Western Blotting Analysis of COX-1 and COX-2 Expression
4.6. Determination of COX-1 and COX-2 Activity
4.7. The Measurements of Interleukin 1β (IL-1β) and Interleukin 6 (IL-6) Concentration
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- United States Environmental Protection Agency (US EPA) Learn about Lead. Available online: https://www.epa.gov/lead/learn-about-lead (accessed on 1 August 2019).
- Agency For Toxic Substances And Disease Registry, Case Studies In Environmental Medicine (CSEM), Lead Toxicity. 2017. Available online: https://www.atsdr.cdc.gov/csem/lead/docs/CSEM-Lead_toxicity_508.pdf (accessed on 1 August 2019).
- Baranowska-Bosiacka, I.; Kosińska, I.; Jamioł, D.; Gutowska, I.; Prokopowicz, A.; Rębacz-Maron, E.; Goschorska, M.; Olszowski, T.; Chlubek, D. Environmental Lead (Pb) Exposure Versus Fatty Acid Content in Blood and Milk of the Mother and in the Blood of Newborn Children. Biol. Trace Elem. Res. 2016, 170, 279–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Needleman, H. Lead poisoning. Annu. Rev. Med. 2004, 55, 209–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapisarda, V.; Ledda, C.; Ferrante, M.; Fiore, M.; Cocuzza, S.; Bracci, M.; Fenga, C. Blood pressure and occupational exposure to noise and lead (Pb): A cross-sectional study. Toxicol. Ind. Health. 2016, 32, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.; Liu, Y.; Buchner, V.; Tchounwou, P.B. Neurotoxic effects and biomarkers of lead exposure: a review. Rev. Environ. Health 2009, 24, 15–45. [Google Scholar] [CrossRef]
- Jakubowski, M. Low-level environmental lead exposure and intellectual impairment in children-the current concepts of risk assessment. Int. J. Occup. Med. Environ. Health. 2011, 24, 1–17. [Google Scholar] [CrossRef]
- Schwartz, J.; Otto, D. Blood lead, hearing thresholds, and neurobehavioral development in children and youth. Arch. Environ. Health 1987, 42, 153–160. [Google Scholar] [CrossRef]
- Bleecker, M.L.; Ford, D.P.; Lindgren, K.N.; Hoese, V.M.; Walsh, K.S.; Vaughan, C.G. Differential effects of lead exposure on components of verbal memory. Occup. Environ. Med. 2005, 62, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Charlet, L.; Chapron, Y.; Faller, P.; Kirsch, R.; Stone, A.T.; Baveye, P.C. Neurodegenerative diseases and exposure to the environmental metals Mn, Pb, and Hg. Coordination Chemistry Reviews 2012, 256, 2147–2163. [Google Scholar] [CrossRef]
- Chibowska, K.; Baranowska-Bosiacka, I.; Falkowska, A.; Gutowska, I.; Goschorska, M.; Chlubek, D. Effect of Lead (Pb) on Inflammatory Processes in the Brain. Int. J. Mol. Sci. 2016, 17, 2140. [Google Scholar] [CrossRef] [Green Version]
- Gąssowska, M.; Baranowska-Bosiacka, I.; Moczydłowska, J.; Frontczak-Baniewicz, M.; Gewartowska, M.; Strużyńska, L.; Gutowska, I.; Chlubek, D.; Adamczyk, A. Perinatal exposure to lead (Pb) induces ultrastructural and molecular alterations in synapses of rat offspring. Toxicology 2016, 373, 13–29. [Google Scholar] [CrossRef]
- Gąssowska, M.; Baranowska-Bosiacka, I.; Moczydłowska, J.; Tarnowski, M.; Pilutin, A.; Gutowska, I.; Strużyńska, L.; Chlubek, D.; Adamczyk, A. Perinatal exposure to lead (Pb) promotes Tau phosphorylation in the rat brain in a GSK-3β and CDK5 dependent manner: Relevance to neurological disorders. Toxicology 2016, 347–349, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Baranowska-Bosiacka, I.; Falkowska, A.; Gutowska, I.; Gąssowska, M.; Kolasa-Wołosiuk, A.; Tarnowski, M.; Chibowska, K.; Lubkowska, A.; Chlubek, D. Glycogen metabolism in brain and neurons - astrocytes metabolic cooperation can be altered by pre- and neonatal lead (Pb) exposure. Toxicology 2017, 390, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Baranowska-Bosiacka, I.; Gutowska, I.; Chlubek, D. Cyclooxygenase pathways. Acta Biochim. Pol. 2014, 61, 639–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.A.; Iadarola, M.; Yang, H.Y.; Dionne, R.A. Expression of COX-1 and COX-2 in a clinical model of acute inflammation. J. Pain 2007, 8, 349–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurumbail, R.G.; Kiefer, J.R.; Marnett, L.J. Cyclooxygenase enzymes: catalysis and inhibition. Curr. Opin. Struct. Biol. 2001, 11, 752–760. [Google Scholar] [CrossRef]
- Lin, W.; Li, Z. Blueberries inhibit cyclooxygenase-1 and cyclooxygenase-2 activity in human epithelial ovarian cancer. Oncol. Lett. 2017, 13, 4897–4904. [Google Scholar] [CrossRef] [Green Version]
- Dubois, R.N.; Abramson, S.B.; Crofford, L.; Gupta, R.A.; Simon, L.S.; Van De Putte, L.B.; Lipsky, P.E. Cyclooxygenase in biology and disease. FASEB J. 1998, 12, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Morita, I. Distinct functions of COX-1 and COX-2. Prostaglandins Other Lipid Mediat. 2002, 68–69, 165–175. [Google Scholar] [CrossRef]
- Park, Y.K.; Hong, K.; Jang, B.C. Transcriptional and translational regulation of COX-2 expression by cadmium in C6 glioma cells. Int. J. Mol. Med. 2012, 30, 960–966. [Google Scholar] [CrossRef] [Green Version]
- Lasa, M.; Mahtanil, K.R.; Fich, A.; Brewer, G.; Saklatvala, J.; Clark, A.R. Regulation of cyclooxygenase 2 mRNA stability by the mitogen-activated protein kinase p38 signaling cascade. Mol. Cell. Biol. 2000, 20, 4265–4274. [Google Scholar] [CrossRef] [Green Version]
- Asmuss, M.; Mullenders, L.H.; Hartwig, A. Interference by toxic metal compounds with isolated zinc finger DNA repair proteins. Toxicol. Lett. 2000, 112–113, 227–231. [Google Scholar] [CrossRef]
- Ramesh, G.T.; Manna, S.K.; Aggarwal, B.B.; Jadhav, A.L. Lead exposure activates nuclear factor kappa B, activator protein-1, c-Jun N-terminal kinase and caspases in the rat brain. Toxicol. Lett. 2001, 123, 195–207. [Google Scholar] [CrossRef]
- Gry, M.; Rimini, R.; Strömberg, S.; Asplund, A.; Pontén, F.; Uhlén, M.; Nilsson, P. Correlations between RNA and protein expression profiles in 23 human cell lines. BMC Genomics 2009, 10, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Du, K.; Cai, Q.; Ma, L.; Jiao, Z.; Tan, J.; Xu, Z.; Li, J.; Luo, W.; Chen, J.; et al. Lead induces COX-2 expression in glial cells in a NFAT-dependent, AP-1/NFκB-independent manner. Toxicology 2014, 325, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Curran, B.; O’Connor, J.J. The pro-inflammatory cytokine interleukin-18 impairs long-term potentiation and NMDA receptor-mediated transmission in the rat hippocampus in vitro. Neuroscience 2001, 108, 83–90. [Google Scholar] [CrossRef]
- Chou, Y.H.; Woon, P.Y.; Huang, W.C.; Shiurba, R.; Tsai, Y.T.; Wang, Y.S.; Hsieh, T.J.; Chang, W.C.; Chuang, H.Y.; Chang, W.C. Divalent lead cations induce cyclooxygenase-2 gene expression by epidermal growth factor receptor/nuclear factor-kappa B signaling in A431carcinoma cells. Toxicology Letters 2011, 203, 147–153. [Google Scholar] [CrossRef]
- Simões, M.R.; Aguado, A.; Fiorim, J.; Silveira, E.A.; Azevedo, B.F.; Toscano, C.M.; Zhenyukh, O.; Briones, A.M.; Alonso, M.J.; Vassallo, D.V.; et al. MAPK pathway activation by chronic lead-exposure increases vascular reactivity through oxidative stress/cyclooxygenase-2-dependent pathways. Toxicol. Appl. Pharmacol. 2015, 283, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Aid, S.; Caracciolo, L.; Minami, S.S.; Niikura, T.; Matsuoka, Y.; Turner, R.S.; Mattson, M.P.; Bosetti, F. Cyclooxygenase-1 inhibition reduces amyloid pathology and improves memory deficits in a mouse model of Alzheimer’s disease. J. Neurochem. 2013, 124, 59–68. [Google Scholar] [CrossRef]
- Xiang, Z.; Ho, L.; Yemul, S.; Zhao, Z.; Qing, W.; Pompl, P.; Kelley, K.; Dang, A.; Qing, W.; Teplow, D.; et al. Cyclooxygenase-2 Promotes Amyloid Plaque Deposition in a Mouse Model of Alzheimer’s Disease Neuropathology. Gene Expr. 2002, 10, 271–278. [Google Scholar] [CrossRef]
- Hoozemans, J.J.; Rozemuller, J.M.; van Haastert, E.S.; Veerhuis, R.; Eikelenboom, P. Cyclooxygenase-1 and -2 in the different stages of Alzheimer’s disease pathology. Curr. Pharm. Des. 2008, 14, 1419–1427. [Google Scholar] [CrossRef]
- Teismann, P.; Tieu, K.; Choi, D.K.; Wu, D.C.; Naini, A.; Hunot, S.; Vila, M.; Jackson-Lewis, V.; Przedborski, S. Cyclooxygenase-2 is instrumental in Parkinson’s disease neurodegeneration. Proc. Natl. Acad. Sci. USA 2003, 100, 5473–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, A.L.; Leenders, K.L. Cyclooxygenase and neuroinflammation in Parkinson’s disease neurodegeneration. Curr. Neuropharmacol. 2010, 8, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Bueno, B.; Serrats, J.; Sawchenko, P.E. Cerebrovascular cyclooxygenase-1 expression, regulation, and role in hypothalamic-pituitary-adrenal axis activation by inflammatory stimuli. J. Neurosci. 2009, 29, 12970–12981. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Aid, S.; Choi, U.; Bosetti, F. Cyclooxygenases-1 and -2 differentially modulate leukocyte recruitment into the inflamed brain. Pharmacogenomics J. 2010, 10, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matousek, S.B.; Hein, A.M.; Shaftel, S.S.; Olschowka, J.A.; Kyrkanides, S.; O’Banion, M.K. Cyclooxygenase-1 mediates prostaglandin E(2) elevation and contextual memory impairment in a model of sustained hippocampal interleukin-1beta expression. J. Neurochem. 2010, 114, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargahi, L.; Nasiraei-Moghadam, S.; Abdi, A.; Khalaj, L.; Moradi, F.; Ahmadiani, A. Cyclooxygenase (COX)-1 activity precedes the COX-2 induction in Aβ-induced neuroinflammation. J. Mol. Neurosci. 2011, 45, 10–21. [Google Scholar] [CrossRef]
- Hoozemans, J.J.; Rozemuller, A.J.; Janssen, I.; De Groot, C.J.; Veerhuis, R.; Eikelenboom, P. Cyclooxygenase expression in microglia and neurons in Alzheimer’s disease and control brain. Acta. Neuropathol. 2001, 101, 2–8. [Google Scholar] [CrossRef]
- Yermakova, A.V.; Rollins, J.; Callahan, L.M.; Rogers, J.; O’Banion, M.K. Cyclooxygenase-1 in human Alzheimer and control brain: quantitative analysis of expression by microglia and CA3 hippocampal neurons. J. Neuropathol. Exp. Neurol. 1999, 58, 1135–1146. [Google Scholar] [CrossRef] [Green Version]
- Kaur, C.; Rathnasamy, G.; Ling, E.A. Biology of Microglia in the Developing Brain. J. Neuropathol. Exp. Neurol. 2017, 76, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Calvello, R.; Lofrumento, D.D.; Perrone, M.G.; Cianciulli, A.; Salvatore, R.; Vitale, P.; De Nuccio, F.; Giannotti, L.; Nicolardi, G.; Panaro, M.A.; et al. Highly Selective Cyclooxygenase-1 Inhibitors P6 and Mofezolac Counteract Inflammatory State both In Vitro and In Vivo Models of Neuroinflammation. Front. Neurol. 2017, 8, 251. [Google Scholar] [CrossRef] [Green Version]
- Saliba, S.W.; Marcotegui, A.R.; Fortwängler, E.; Ditrich, J.; Perazzo, J.C.; Muñoz, E.; de Oliveira, A.C.P.; Fiebich, B.L. AM404, paracetamol metabolite, prevents prostaglandin synthesis in activated microglia by inhibiting COX activity. J. Neuroinflammation 2017, 14, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, L.; Purohit, D.; Haroutunian, V.; Luterman, J.D.; Willis, F.; Naslund, J.; Buxbaum, J.D.; Mohs, R.C.; Aisen, P.S.; Pasinetti, G.M. Neuronal cyclooxygenase 2 expression in the hippocampal formation as a function of the clinical progression of Alzheimer disease. Arch. Neurol. 2001, 58, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Pieroni, C.; Winger, D.; Purohit, D.P.; Aisen, P.S.; Pasinetti, G.M. Regional distribution of cyclooxygenase-2 in the hippocampal formation in Alzheimer’s disease. J. Neurosci. Res. 1999, 57, 295–303. [Google Scholar] [CrossRef]
- Pasinetti, G.M.; Aisen, P.S. Cyclooxygenase-2 expression is increased in frontal cortex of Alzheimer’s disease brain. Neuroscience 1998, 87, 319–324. [Google Scholar] [CrossRef]
- Shanmugam, N.; Gaw Gonzalo, I.T.; Natarajan, R. Molecular mechanisms of high glucose-induced cyclooxygenase-2 expression in monocytes. Diabetes 2004, 53, 795–802. [Google Scholar] [CrossRef] [Green Version]
- Miao, F.; Gonzalo, I.G.; Lanting, L.; Natarajan, R. In vivo chromatin remodeling events leading to inflammatory gene transcription under diabetic conditions. J. Biol. Chem. 2004, 279, 18091–18097. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Knowles, S.O.; Donaldson, W.E. Lead disrupts eicosanoid metabolism, macrophage function, and disease resistance in birds. Biol. Trace Elem. Res. 1997, 60, 13–26. [Google Scholar] [CrossRef]
- Lee, J.J.; Battles, A.H. Lead toxicity via arachidonate signal transduction to growth responses in the splenic macrophage. Environ. Res. 1994, 67, 209–219. [Google Scholar] [CrossRef]
- Flohé, S.B.; Brüggemann, J.; Herder, C.; Goebel, C.; Kolb, H. Enhanced proinflammatory response to endotoxin after priming of macrophages with lead ions. J. Leukoc. Biol. 2002, 71, 417–424. [Google Scholar]
- Miyahara, T.; Komiyama, H.; Miyanishi, A.; Matsumoto, M.; Xue-Ya, W.T.M.; Takata, S.; Nagai, M.; Kozuka, H.; Yokoyama, K.; Kanamoto, Y. Effects of lead on osteoclast-like cell formation in mouse bone marrow cell cultures. Calcified Tissue International 1994, 54, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer. 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z. The use of THP-1 cells as a model for mimicking the function and regulation of monocytes and macrophages in the vasculature. Atherosclerosis 2012, 221, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J.; Miller, K.R.; Lopes, K.O.; Njie, E. Microglial degeneration in the aging brain - bad news for neurons? Front. Biosci. 2008, 13, 3423–3438. [Google Scholar] [CrossRef] [Green Version]
- Goschorska, M.; Baranowska-Bosiacka, I.; Gutowska, I.; Tarnowski, M.; Piotrowska, K.; Metryka, E.; Safranow, K.; Chlubek, D. Effect of acetylcholinesterase inhibitors donepezil and rivastigmine on the activity and expression of cyclooxygenases in a model of the inflammatory action of fluoride on macrophages obtained from THP-1 monocytes. Toxicology 2018, 406–407, 9–20. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- An, C.; Shi, Y.; Li, P.; Hu, X.; Yu, G.; Stetler, R.A.; Leak, R.K.; Gao, Y.; Sun, B.L.; Zheng, P.; et al. Molecular dialogs between the ischemic brain and the peripheral immune system: dualistic roles in injury and repair. Prog. Neurobiol. 2014, 115, 6–24. [Google Scholar] [CrossRef] [Green Version]
- Olszowski, T.; Baranowska-Bosiacka, I.; Gutowska, I.; Piotrowska, K.; Mierzejewska, K.; Korbecki, J.; Kurzawski, M.; Tarnowski, M.; Chlubek, D. The Effects of Cadmium at Low Environmental Concentrations on THP-1 Macrophage Apoptosis. Int. J. Mol. Sci. 2015, 16, 21410–21427. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Baranowska-Bosiacka, I.; Gutowska, I.; Piotrowska, K.; Chlubek, D. Cyclooxygenase-1 as the main source of proinflammatory factors after sodium orthovanadate treatment. Biol. Trace Elem. Res. 2015, 163, 103–111. [Google Scholar] [CrossRef] [Green Version]
- CDC. Low Level Lead Exposure Harms Children: A Renewed Call for Primary Prevention. Report of the Advisory Committee on Childhood Lead Poisoning Prevention of the Centers for Disease Control and Prevention. 2012. Available online: https://www.cdc.gov/nceh/lead/acclpp/final_document_030712.pdf (accessed on 25 July 2019).
- Nandi, A.; Chandil, D.; Lechesal, R.; Pryor, S.C.; McLaughlin, A.; Bonventre, J.A.; Flynnx, K.; Weeks, B.S. Bifenthrin causes neurite retraction in the absence of cell death: a model for pesticide associated neurodegeneration. Med. Sci. Monit. 2006, 12, BR169–BR173. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metryka, E.; Kupnicka, P.; Kapczuk, P.; Simińska, D.; Tarnowski, M.; Goschorska, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages. Int. J. Mol. Sci. 2020, 21, 2254. https://doi.org/10.3390/ijms21062254
Metryka E, Kupnicka P, Kapczuk P, Simińska D, Tarnowski M, Goschorska M, Gutowska I, Chlubek D, Baranowska-Bosiacka I. Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages. International Journal of Molecular Sciences. 2020; 21(6):2254. https://doi.org/10.3390/ijms21062254
Chicago/Turabian StyleMetryka, Emilia, Patrycja Kupnicka, Patrycja Kapczuk, Donata Simińska, Maciej Tarnowski, Marta Goschorska, Izabela Gutowska, Dariusz Chlubek, and Irena Baranowska-Bosiacka. 2020. "Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages" International Journal of Molecular Sciences 21, no. 6: 2254. https://doi.org/10.3390/ijms21062254
APA StyleMetryka, E., Kupnicka, P., Kapczuk, P., Simińska, D., Tarnowski, M., Goschorska, M., Gutowska, I., Chlubek, D., & Baranowska-Bosiacka, I. (2020). Lead (Pb) as a Factor Initiating and Potentiating Inflammation in Human THP-1 Macrophages. International Journal of Molecular Sciences, 21(6), 2254. https://doi.org/10.3390/ijms21062254