Effective Cellular Transport of Ortho-Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Expression of MATE1–2, PMAT and the Function of OCTs in MCF-7 and MDA-MB-231 Cells
2.2. Inhibition of [14C]Choline Uptake in MCF-7 and MDA-MB-231 Cells
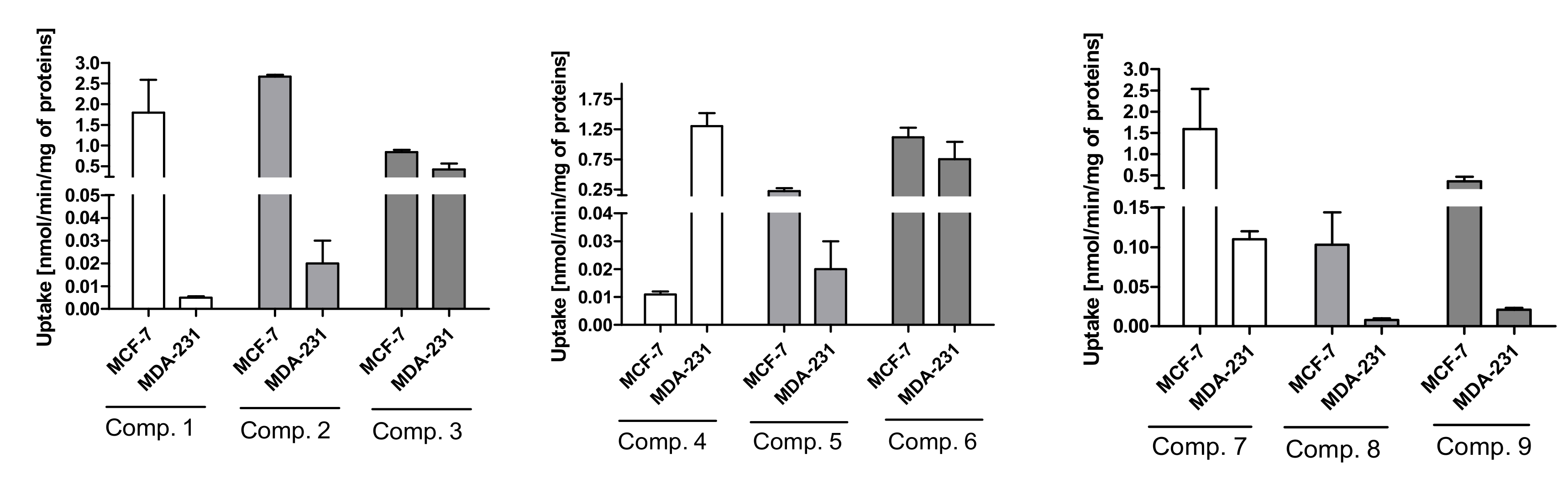
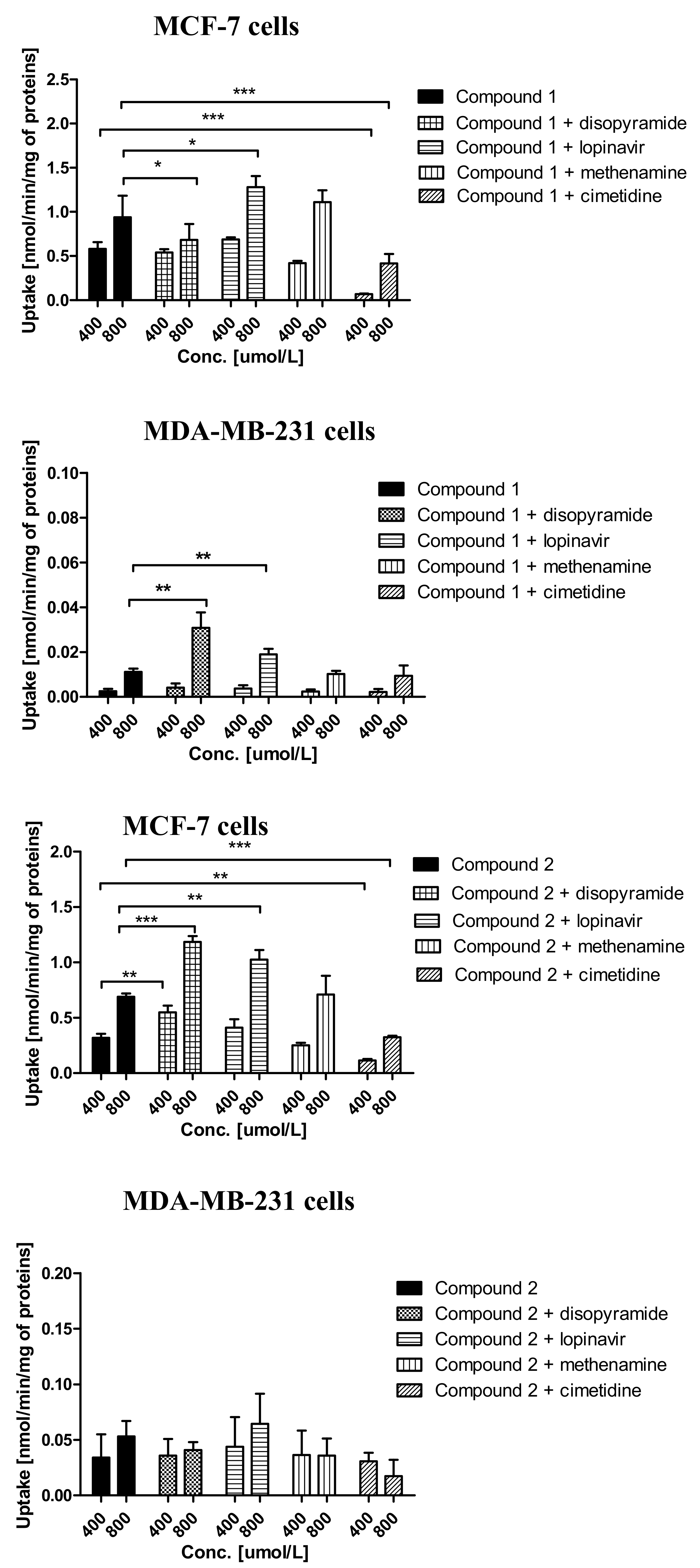
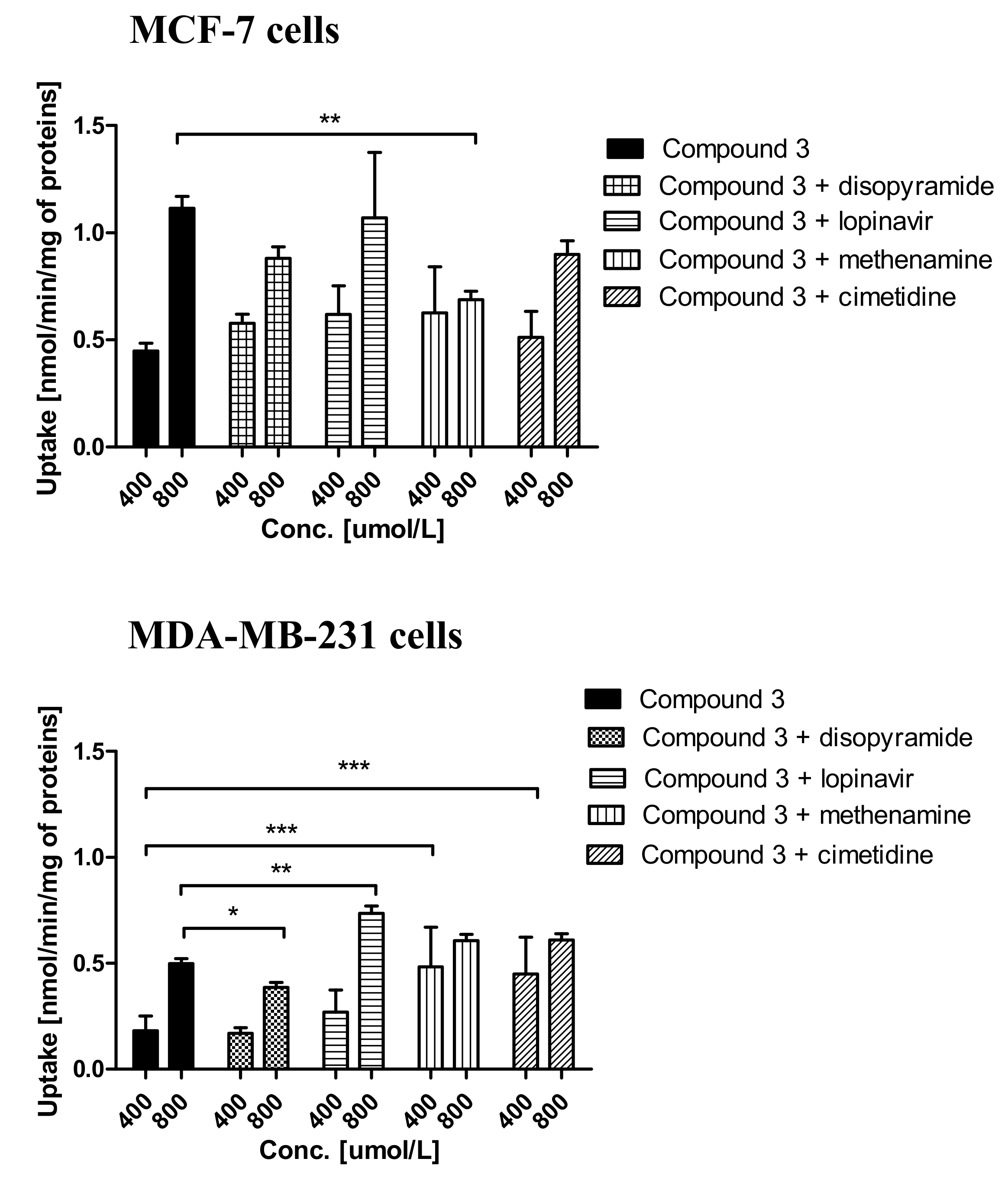
2.3. Cellular Uptake of Metformin Derivatives
2.3.1. General Characterization
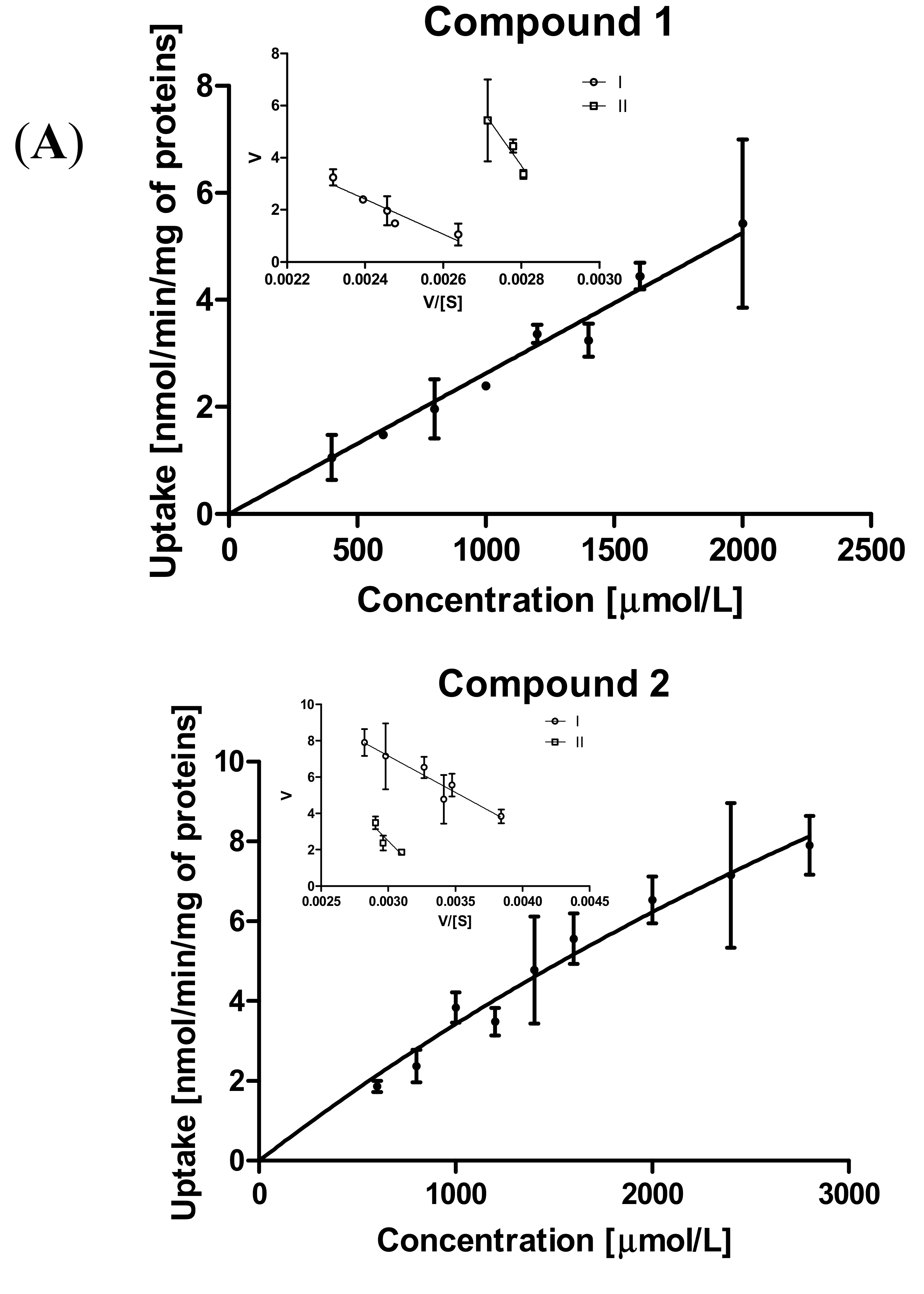
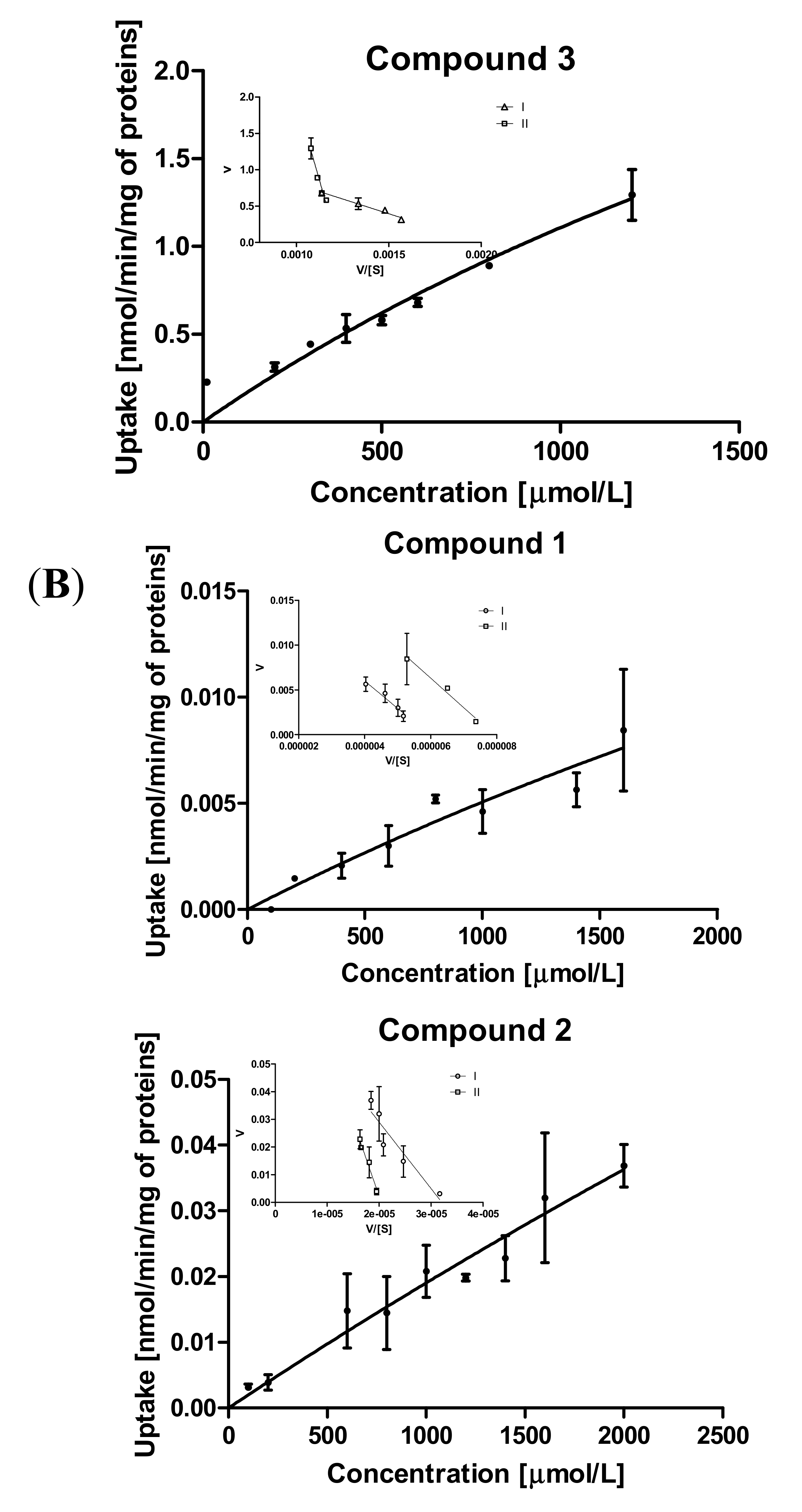
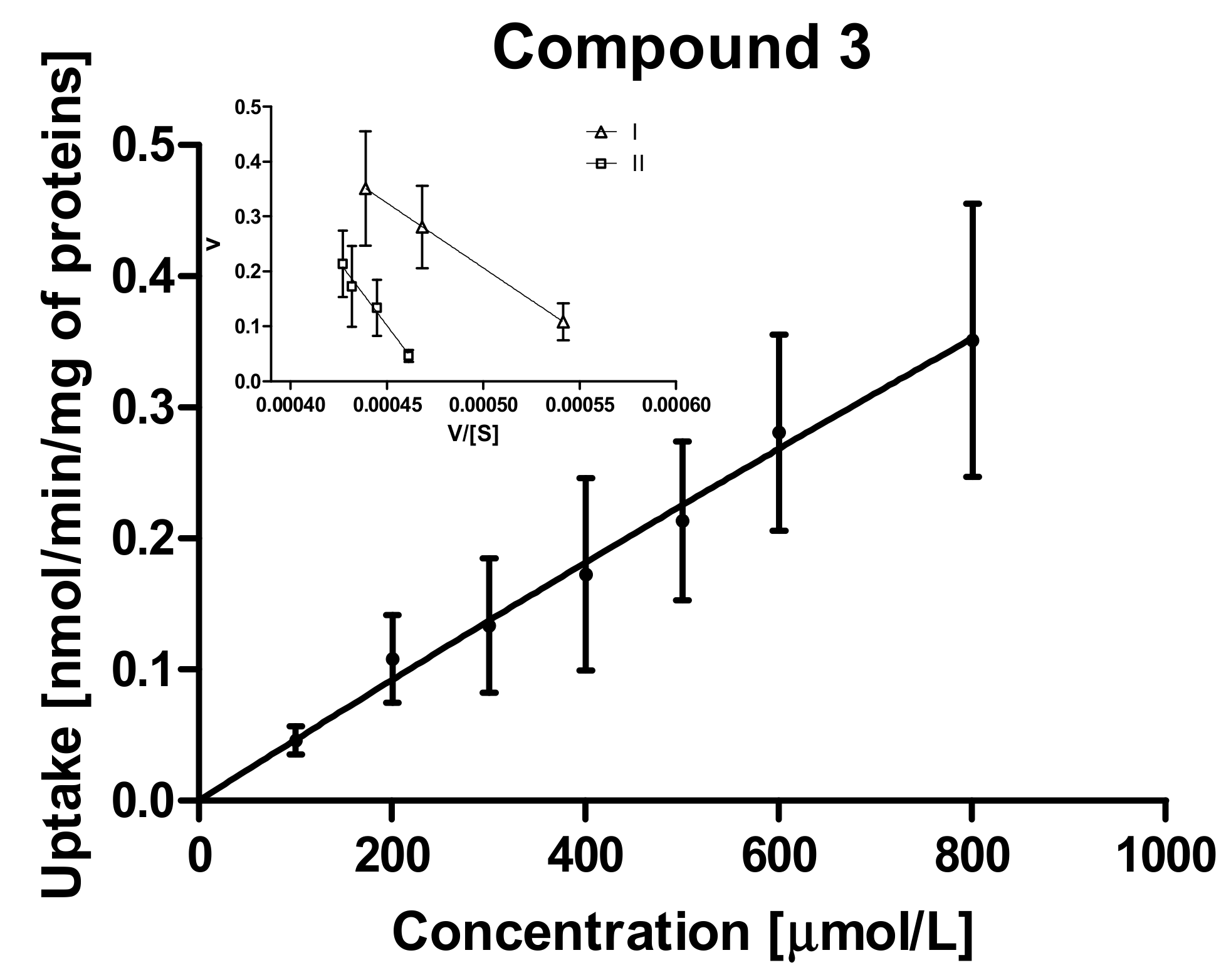
2.3.2. Kinetic Analysis of Sulfonamide Uptake in MCF-7 and MDA-MB-231 Cells
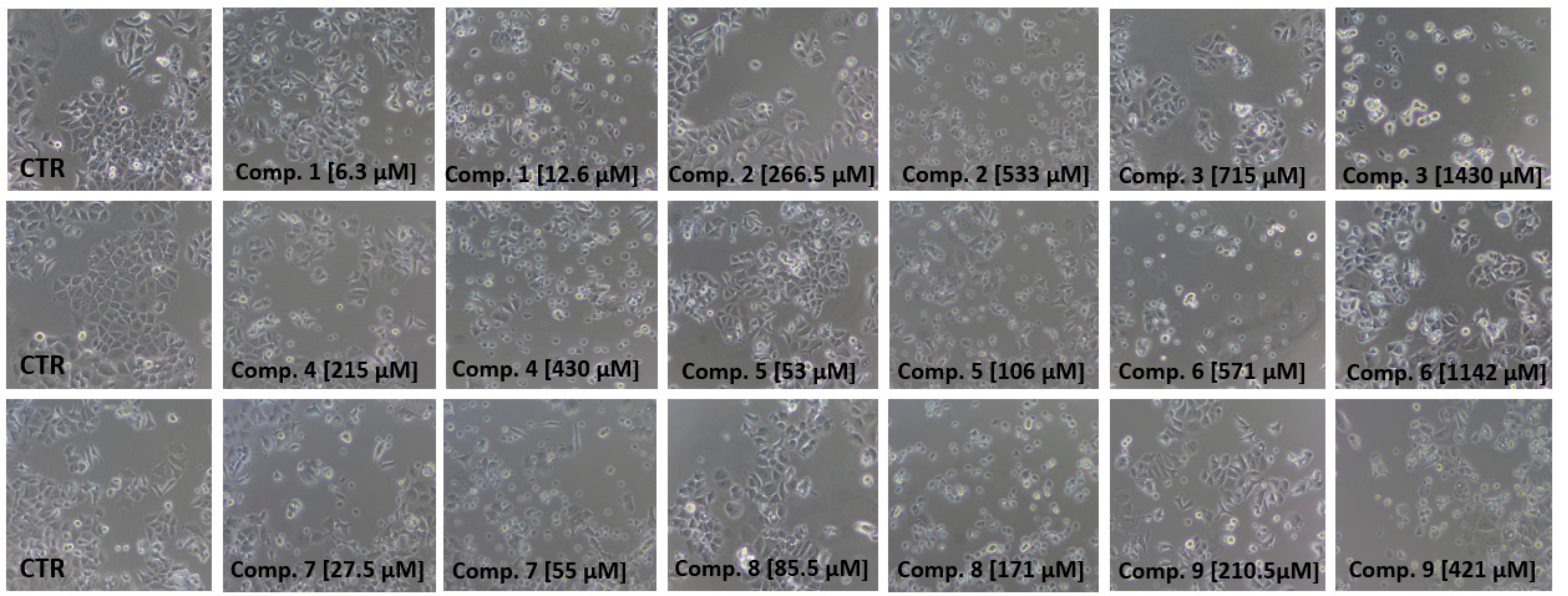
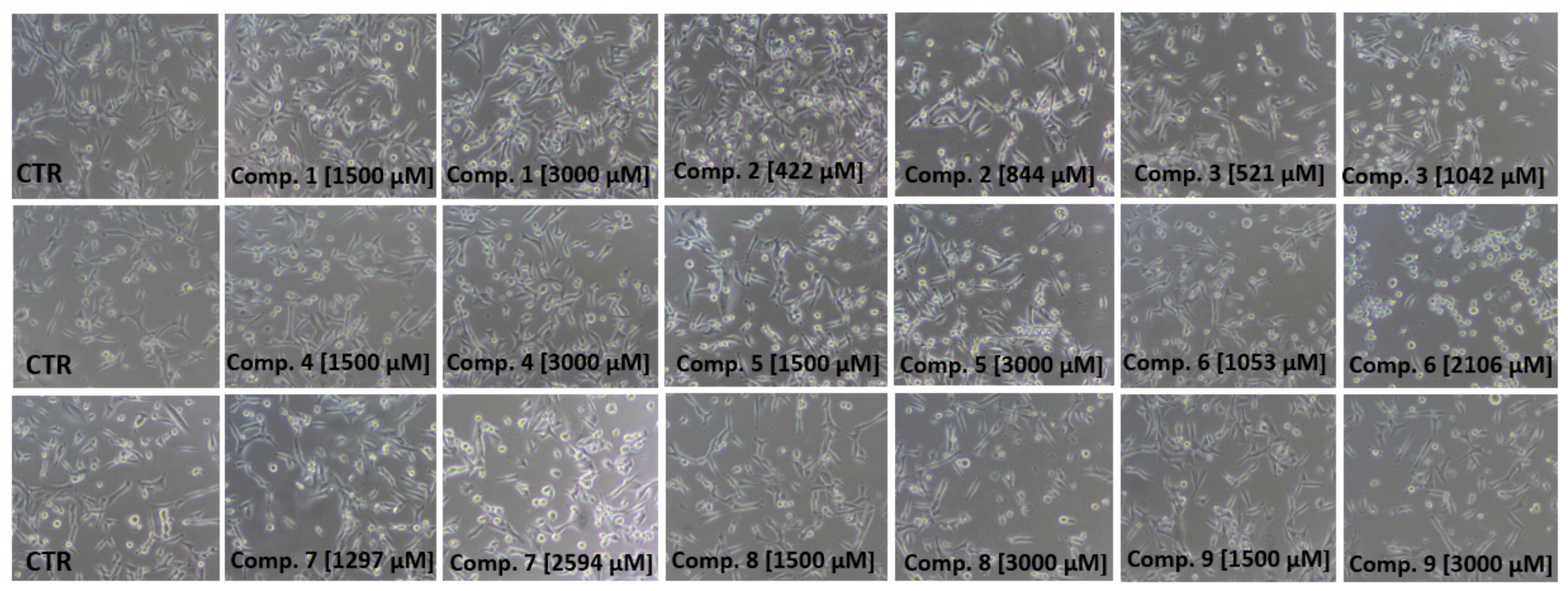
2.4. Cell Viability Assay
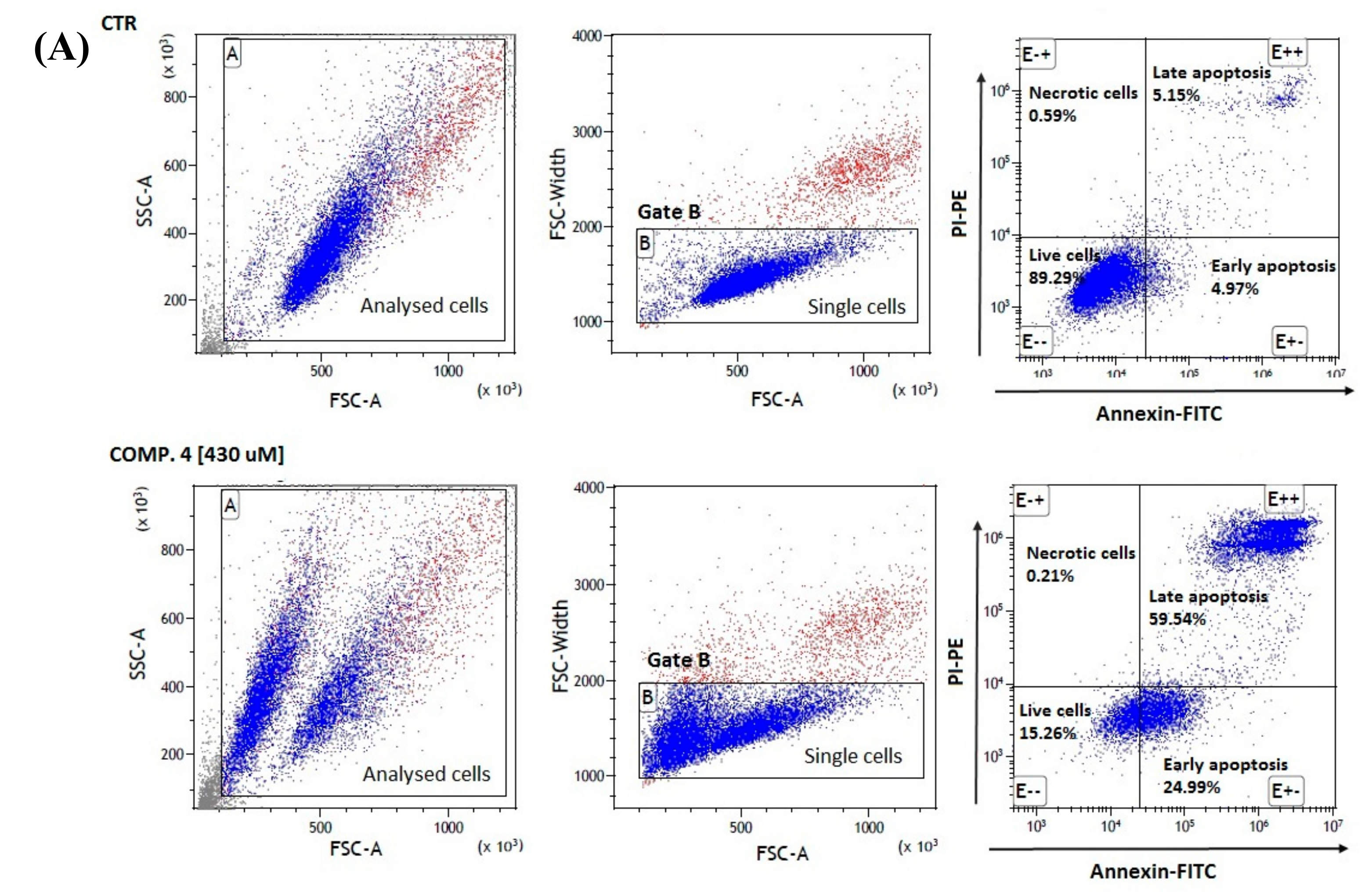
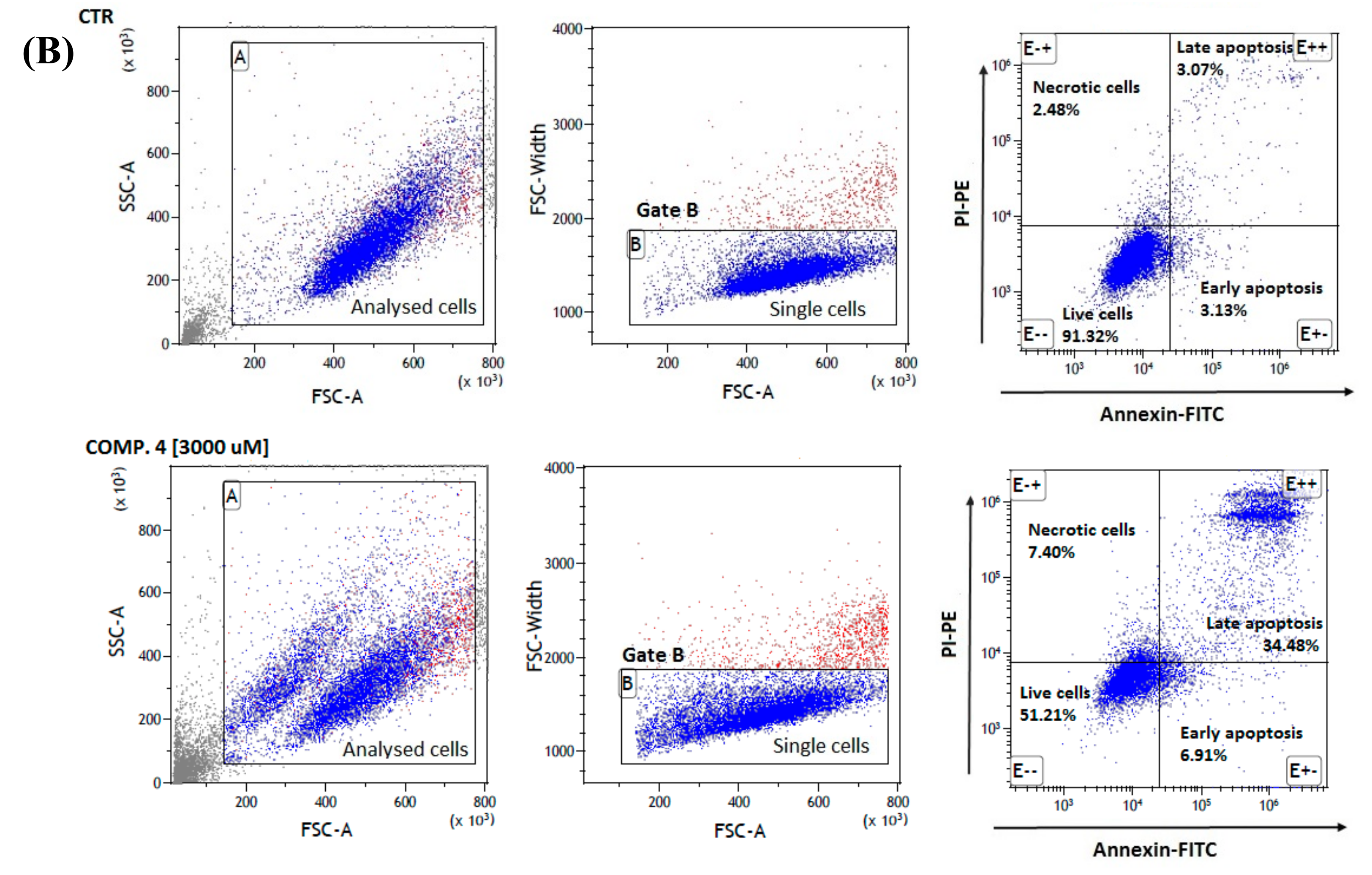
2.5. Apoptosis Assay
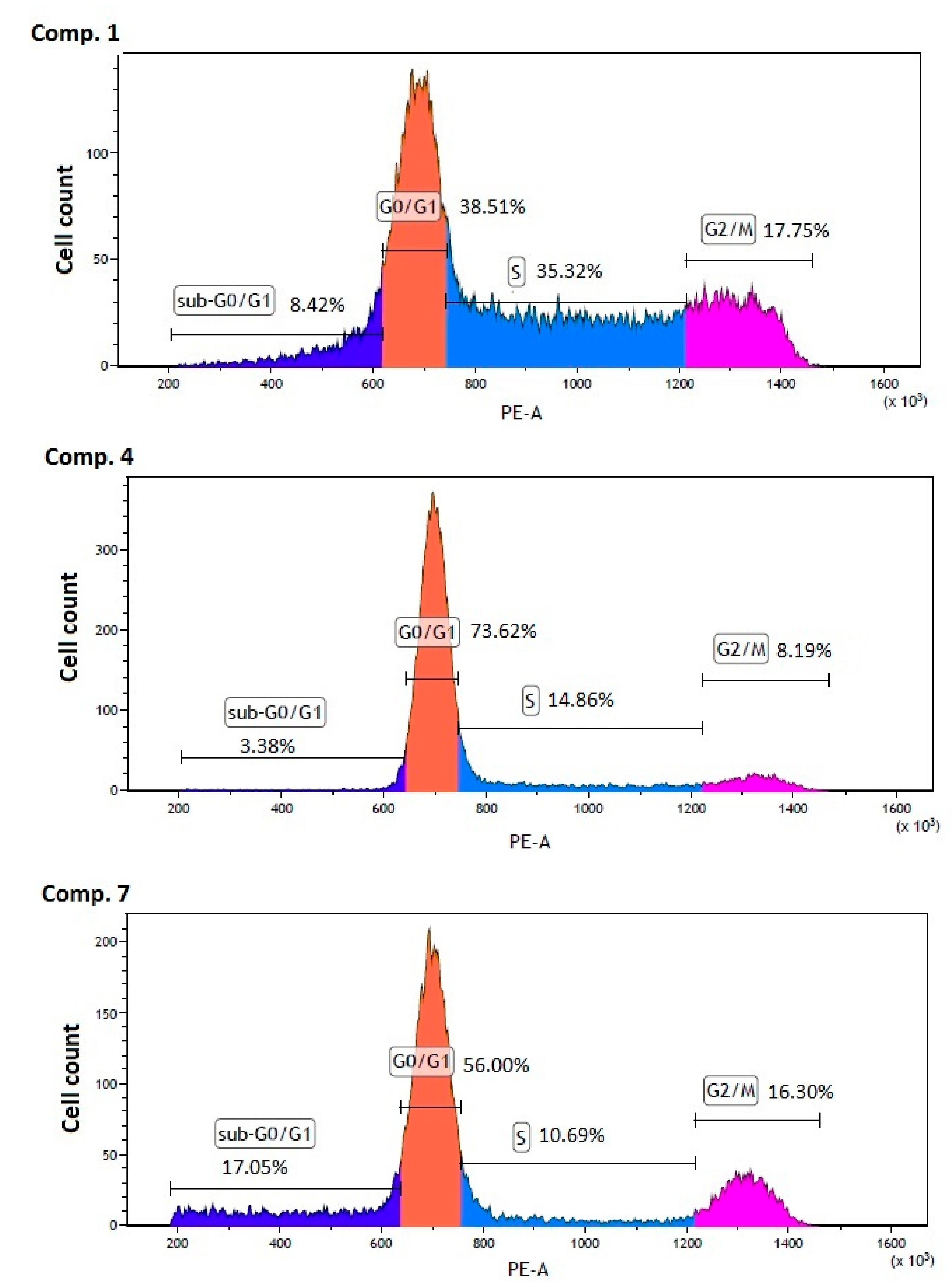
2.6. Cell Cycle Arrest
2.7. GSH:GSSG Ratio
3. Materials and Methods
3.1. Cell Culturing
3.2. Studied Compounds
3.3. PMAT and MATE1–2 Gene Expression and Function of OCTs in MCF-7 and MDA-MB-231 Cells
3.4. Inhibition of [14C]Choline Uptake
3.5. Uptake Studies
3.6. Uptake of Metformin Derivatives in the Presence of OCT and MATE Inhibitors
3.7. Cell Viability and Morphology
3.8. Cell Apoptosis Assay
3.9. Cell Cycle Analysis
3.10. GSH:GSSG Ratio
3.11. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prattichizzo, F.; Giuliani, A.; Mensà, E.; Sabbatinelli, J.; De Nigris, V.; Rippo, M.R.; La Sala, L.; Procopio, A.D.; Olivieri, F.; Ceriello, A. Pleiotropic effects of metformin: Shaping the microbiome to manage type 2 diabetes and postpone ageing. Ageing Res. Rev. 2018, 48, 87–98. [Google Scholar] [CrossRef]
- Mahmood, K.; Naeem, M.; Rahimnajjad, N.A. Metformin: The hidden chronicles of a magic drug. Eur. J. Intern. Med. 2013, 24, 20–26. [Google Scholar] [CrossRef]
- Markowicz-Piasecka, M.; Huttunen, K.M.; Mikiciuk-Olasik, E.; Mateusiak, Ł.; Sikora, J. Metformin - from anti-diabetic drug to anti-cancer drug. Acta Pol. Pharm. Drug. Res. 2018, 1, 3–18. [Google Scholar]
- Franciosi, M.; Lucisano, G.; Lapice, E.; Strippoli, G.F.M.; Pellegrini, F.; Nicolucci, A. Metformin Therapy and Risk of Cancer in Patients with Type 2 Diabetes: Systematic Review. PLOS ONE 2013, 8, e71583. [Google Scholar] [CrossRef]
- Zakikhani, M.; Dowling, R.; Fantus, I.G.; Sonenberg, N.; Pollak, M. Metformin is an AMP Kinase-Dependent Growth Inhibitor for Breast Cancer Cells. Obstet. Gynecol. Surv. 2007, 62, 182–183. [Google Scholar] [CrossRef]
- Wahdan-Alaswad, R.S.; Cochrane, D.R.; Spoelstra, N.S.; Howe, E.; Edgerton, S.M.; Anderson, S.M.; Thor, A.D.; Richer, J.K. Metformin-induced killing of triple-negative breast cancer cells is mediated by reduction in fatty acid synthase via miRNA-193b. Horm. Cancer 2014, 5, 374–389. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.-S.; Wang, S.; Deng, A.; Liu, B.; Edgerton, S.M.; Lind, S.E.; Wahdan-Alaswad, R.; Thor, A.D. Metformin targets Stat3 to inhibit cell growth and induce apoptosis in triple-negative breast cancers. Cell Cycle 2012, 11, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Saraei, P.; Asadi, I.; Kakar, M.A.; Moradi-Kor, N. The beneficial effects of metformin on cancer prevention and therapy: a comprehensive review of recent advances. Cancer Manag. Res. 2019, 11, 3295–3313. [Google Scholar] [CrossRef] [Green Version]
- Pollak, M. Investigating Metformin for Cancer Prevention and Treatment: The End of the Beginning. Cancer Discov. 2012, 2, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Goswamic, S.; Giacominic, K.M.; Altmana, R.B.; Klein, T.E. Metformin pathways: pharmacokinetics and pharmacodynamic. Pharmacogenet. Genomics. 2012, 22, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Zi, F.; Zi, H.; Li, Y.; He, J.; Shi, Q.; Cai, Z. Metformin and cancer: An existing drug for cancer prevention and therapy. Oncol. Lett. 2017, 15, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Zielonka, J.; Ouari, O.; Lopez, M.; McAllister, D.; Boyle, K.; Barrios, C.S.; Weber, J.J.; Johnson, B.D.; Hardy, M.; et al. Mitochondria-Targeted Analogues of Metformin Exhibit Enhanced Antiproliferative and Radiosensitizing Effects in Pancreatic Cancer Cells. Cancer Res. 2016, 76, 3904–3915. [Google Scholar] [CrossRef] [Green Version]
- Markowicz-Piasecka, M.; Huttunen, J.; Sikora, J.; Huttunen, K.M. Sulfenamide derivatives can improve transporter-mediated cellular uptake of metformin andinduce cytotoxicity in human breast adenocarcinoma cell lines. Bioorg. Chem. 2019, 87, 321–334. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, Y.; Han, T.K.; Everett, R.; Thakker, D.R. Cation-selective transporters are critical to the AMPK-mediated antiproliferative effects of metformin in human breast cancer cells. Int. J. Cancer 2016, 138, 2281–2292. [Google Scholar] [CrossRef] [Green Version]
- DeCensi, A.; Puntoni, M.; Goodwin, P.J.; Cazzaniga, M.; Gennari, A.; Bonanni, B.; Gandini, S. Metformin and Cancer Risk in Diabetic Patients: A Systematic Review and Meta-analysis. Cancer Prev. Res. 2010, 3, 1451–1461. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.M.M.; A Donnelly, L.; Emslie-Smith, A.M.; Alessi, D.R.; Morris, A.D. Metformin and reduced risk of cancer in diabetic patients. BMJ 2005, 330, 1304–1305. [Google Scholar] [CrossRef] [Green Version]
- Checkley, L.A.; Rudolph, M.C.; Wellberg, E.A.; Giles, E.D.; Wahdan-Alaswad, R.S.; Houck, J.A.; Edgerton, S.M.; Thor, A.D.; Schedin, P.; Anderson, S.M.; et al. Metformin Accumulation Correlates with Organic Cation Transporter 2 Protein Expression and Predicts Mammary Tumor Regression In Vivo. Cancer Prev. Res. 2017, 10, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Štaud, F.; Cerveny, L.; Ahmadimoghaddam, D.; Ceckova, M. Multidrug and toxin extrusion proteins (MATE/SLC47); role in pharmacokinetics. Int. J. Biochem. Cell Boil. 2013, 45, 2007–2011. [Google Scholar] [CrossRef]
- Sjögren, E.; Lennernas, H.; Andersson, T.B.; Gråsjö, J.; Bredberg, U. The Multiple Depletion Curves Method Provides Accurate Estimates of Intrinsic Clearance (CLint), Maximum Velocity of the Metabolic Reaction (Vmax), and Michaelis Constant (Km): Accuracy and Robustness Evaluated through Experimental Data and Monte Carlo Simulations. Drug Metab. Dispos. 2008, 37, 47–58. [Google Scholar]
- Krisnamurti, D.G.B.; Louisa, M.; Anggraeni, E.; Wanandi, S.I. Drug Efflux Transporters Are Overexpressed in Short-Term Tamoxifen-Induced MCF7 Breast Cancer Cells. Adv. Pharmacol. Sci. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Chang, F.-W.; Fan, H.-C.; Liu, J.-M.; Fan, T.-P.; Jing, J.; Yang, C.-L.; Hsu, R.-J. Estrogen Enhances the Expression of the Multidrug Transporter Gene ABCG2—Increasing Drug Resistance of Breast Cancer Cells through Estrogen Receptors. Int. J. Mol. Sci. 2017, 18, 163. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Uray, I.P.; Mazumdar, A.; Mayer, J.A.; Brown, P.H. SLC22A5/OCTN2 expression in breast cancer is induced by estrogen via a novel intronic estrogen-response element (ERE). Breast Cancer Res. Treat. 2012, 134, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Czuba, L.; Hillgren, K.M.; Swaan, P. Post-translational modifications of transporters. Pharmacol. Ther. 2018, 192, 88–99. [Google Scholar] [CrossRef]
- Markowicz-Piasecka, M.; Sikora, J.; Zajda, A.; Huttunen, K.M. Novel halogenated sulfonamide biguanides with anti-coagulation properties. Bioorganic Chem. 2020, 94, 103444. [Google Scholar] [CrossRef]
- Olszewska, P.; Cal, D.; Zagórski, P.; Mikiciuk-Olasik, E. A novel trifluoromethyl 2-phosphonopyrrole analogue inhibits human cancer cell migration and growth by cell cycle arrest at G1 phase and apoptosis. Eur. J. Pharmacol. 2020, 871, 172943. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Torres, G.C.; Baltiérrez-Hoyos, R.; Andrade-Jorge, E.; Villa-Treviño, S.; Trujillo-Ferrara, J.G.; Vásquez-Garzón, V.R. Cytotoxicity, Oxidative Stress, Cell Cycle Arrest, and Mitochondrial Apoptosis after Combined Treatment of Hepatocarcinoma Cells with Maleic Anhydride Derivatives and Quercetin. Oxidative Med. Cell. Longev. 2017, 2017, 1–16. [Google Scholar] [CrossRef]















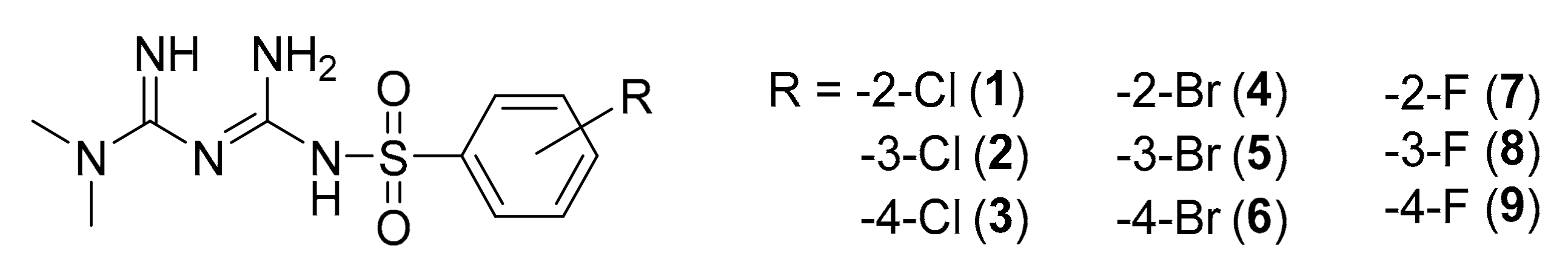{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 MCF-7 (μmol/L) | IC50 MDA-MB-231 (μmol/L) |
|---|---|---|
| 1 | 1053 ± 14.2 | 2135 ± 14.0 |
| 2 | 2923 ± 9.1 | 4449 ± 18.1 |
| 3 | NE | 3757.1 ± 124.4 |
| 4 | 1670 ± 8.5 | 919.60 ± 13.0 |
| 5 | 888.5 ± 11.4 | 593.1 ± 15.0 |
| 6 | 2062.5 ± 58.4 | 2489.9 ± 211.1 |
| 7 | 6354 ± 12.9 | 5149 ± 22.0 |
| 8 | 7195 ± 5.3 | 2284 ± 14.0 |
| 9 | 1057 ± 7.5 | 1383 ± 14.0 |
| Kinetic Parameters of Prodrug Uptakes | |||||
|---|---|---|---|---|---|
| MCF-7 | MDA-MB-231 Cells | ||||
| Compound | Transp./Efficacy (mL/(min∙mg)) | Km (μmol/L) | Vmax (nmol/min/mg) | Km (μmol/L) | Vmax (nmol/min/mg) |
| 1 | I | 6719.0 ± 981.9 | 18.53 ± 2.41 | 3070.0 ± 561.6 | 0.0182 ± 0.0026 |
| Vmax/Km # | 0.00276 | 0.000006 | |||
| II | 21130.0 ± 7726 | 62.84 ± 21.37 | 3298.0 ± 625.0 | 0.0261 ± 0.0040 | |
| Vmax/Km # | 0.00297 | 0.000008 | |||
| 2 | I | 4014.0 ± 695.8 | 19.2 ± 2.31 | 2404.0 ± 353.3 | 0.077 ± 0.008 |
| Vmax/Km # | 0.00478 | 0.00003 | |||
| II | 7474.0 ± 1821.0 | 24.91 ± 5.44 | 5475.0 ± 730.0 | 0.111 ± 0.016 | |
| Vmax/Km # | 0.00333 | 0.00002 | |||
| 3 | I | 812.7 ± 86.50 | 1.614 ± 0.11 | 2366.0 ± 479.9 | 1.390 ± 0.23 |
| Vmax/Km # | 0.0019 | 0.0006 | |||
| II | 8814 ± 791.3 | 10.76 ± 0.88 | 4666.0 ± 932.4 | 2.20 ± 0.41 | |
| Vmax/Km # | 0.0012 | 0.0005 | |||
| 4 | I | 1540.0 ± 220.3 | 0.031 ± 0.002 | 5516.0 ± 748.2 | 10.27 ± 1.18 |
| Vmax/Km # | 0.0000201 | 0.0018 | |||
| II | 8367.0 ± 1425.0 | 0.153 ± 0.022 | 11570.0 ± 2436 | 21.44 ± 4.03 | |
| Vmax/Km # | 0.0000183 | 0.0018 | |||
| 5 | I | 3616.0 ± 619.2 | 1.635 ± 0.21 | 467.7 ± 108.8 | 0.038 ± 0.004 |
| Vmax/Km # | 0.00045 | 0.000081 | |||
| II | 6846.0 ± 712.9 | 2.167 ± 0.196 | 1071 ± 204.0 | 0.098 ± 0.011 | |
| Vmax/Km # | 0.000316 | 0.000092 | |||
| 6 | I | 307.6 ± 101.8 | 0.7898 ± 0.12 | 1934.0 ± 222.8 | 2.644 ± 0.22 |
| Vmax/Km # | 0.00257 | 0.00137 | |||
| II | 8933.0 ± 1099 | 13.64 ± 1.55 | 2110.0 ± 172.4 | 2.091 ± 0.15 | |
| Vmax/Km # | 0.00153 | 0.00099 | |||
| 7 | I | 2134.0 ± 814.5 | 5.303 ± 1.334 | 1535.0 ± 298.4 | 0.262 ± 0.035 |
| Vmax/Km # | 0.00248 | 0.00017 | |||
| II | 6141.0 ± 4603 | 22.43 ± 12.78 | 2250.0 ± 189.7 | 0.443 ± 0.026 | |
| Vmax/Km # | 0.00365 | 0.0002 | |||
| 8 | I | 2837.0 ± 778.2 | 0.830 ± 0.171 | 1235.0 ± 1313.0 | 0.049 ± 0.025 |
| Vmax/Km # | 0.000292 | 0.00004 | |||
| II | NE | NE | 3383.0 ± 2133 | 0.045 ± 0.021 | |
| Vmax/Km # | 0.00194 | 0.000013 | |||
| 9 | I | NE | NE | 1880.0 ± 428.6 | 0.097 ± 0.015 |
| Vmax/Km # | NE | 0.00002 | |||
| II | NE | NE | 2219.0 ± 617.7 | 0.053 ± 0.012 | |
| Vmax/Km # | NE | 0.000024 | |||
| COMPOUND |
MCF-7 Cells (µmol/L) |
MDA-MB-231 Cells (µmol/L) |
|---|---|---|
| 1 | 12.6 ± 1.2 | 69.25% # |
| 2 | 533.2 ± 87.4 | 843.9 ± 135.0 |
| 3 | 1430 ± 135.0 | 1042 ± 134.0 |
| 4 | 429.7 ± 119.0 | 56.37% # |
| 5 | 106.2 ± 12.3 | 49.81% # |
| 6 | 1142 ± 139.0 | 2106 ± 137.0 |
| 7 | 55.05 ± 2.74 | 2594 ± 150 |
| 8 | 171.1 ± 13.6 | 67.12% # |
| 9 | 421.6 ± 32.8 | 70.91% # |
| Compound [μmol/L] | Single Cells in Gate B [%] 1 | Living Cells [E−−] [%] 2 | Necrotic Cells [E−+] [%] 2 | Early Apoptotic [E+−] [%] 2 | Late Apoptotic [E++] [%] 2 |
|---|---|---|---|---|---|
| Control MCF-7 | 87.90 ± 0.87 | 88.59 ± 0.96 | 0.93 ± 0.33 | 5.84 ± 0.80 | 4.64 ± 0.57 |
| Comp. 1 [6.5 μmol/L] | 77.51 ± 0.92 *** | 32.82 ± 4.50 *** | 0.67 ± 0.08 | 27.59 ± 1.03 *** | 38.93 ± 3.46 *** |
| Comp. 1 [13 μmol/L] | 82.66 ± 0.68 *** | 29.96 ± 6.41 *** | 0.41 ± 0.02 ** | 35.79 ± 3.78 *** | 33.84 ± 3.17 *** |
| Comp. 4 [215 μmol/L] | 78.90 ± 0.11 *** | 34.17 ± 3.88 *** | 0.45 ± 0.15 ** | 30.69 ± 3.01 *** | 34.69 ± 1.51 *** |
| Comp. 4 [430 μmol/L] | 83.19 ± 0.40 ** | 16.84 ± 1.93 *** | 0.29 ± 0.09 *** | 22.71 ± 2.11 *** | 60.16 ± 2.21 *** |
| Comp. 7 [27.5 μmol/L] | 78.97 ± 0.51 *** | 39.85 ± 5.67 *** | 0.57 ± 0.03 * | 23.59 ± 1.57 *** | 35.98 ± 4.64 *** |
| Comp. 7 [55 μmol/mL] | 79.48 ± 0.82*** | 31.88 ± 4.77 *** | 0.54 ± 0.01 * | 24.35 ± 2.19 *** | 43.24 ± 4.09 *** |
| Control MDA-MB-231 | 93.49 ± 0.47 | 90.68 ± 1.37 | 2.27 ± 0.33 | 3.75 ± 1.39 | 3.31 ± 0.32 |
| Comp. 1 [1500 μmol/L] | 94.34 ± 0.26 | 81.55 ± 1.65 * | 2.74 ± 0.98 | 7.66 ± 0.06 *** | 8.06 ± 0.81 *** |
| Comp. 1 [3000 μmol/L] | 91.85 ± 0.37 ** | 53.83 ± 3.87 *** | 4.81 ± 0.60 | 9.96 ± 0.92 *** | 31.41 ± 3.46 *** |
| Comp. 4 [1500 μmol/L] | 93.37 ± 0.34 | 72.64 ± 1.76 *** | 6.31 ± 0.26 * | 7.74 ± 0.91 *** | 13.30 ± 0.66 *** |
| Comp. 4 [3000 μmol/L] | 90.59 ± 0.70 *** | 49.49 ± 2.31 *** | 7.15 ± 1.45 ** | 6.85 ± 0.34 ** | 36.51 ± 3.36 *** |
| Comp. 7 [1300 μmol/L] | 93.75 ± 0.41 | 68.26 ± 3.51 *** | 4.06 ± 1.75 | 8.78 ± 0.63 *** | 18.89 ± 2.67 *** |
| Comp. 7 [2600 μmol/mL] | 91.60 ± 0.48 ** | 50.40 ± 4.95 *** | 7.66 ± 2.93 ** | 6.39 ± 0.16 ** | 35.54 ± 2.67 *** |
| Compound [µmol/L] | Sub G0/G1 [%] | G0/G1 [%] | S [%] | G2/M [%] |
|---|---|---|---|---|
| Control MCF-7 | 1.65 ± 0.19 | 45.56 ± 0.68 | 33.63 ± 0.02 | 19.20 ± 0.90 |
| Comp. 1 [13 µmol/L] | 8.28 ± 1.04 *** | 38.76 ± 0.80 ** | 35.45 ± 0.23 ** | 17.54 ± 0.48 * |
| Comp. 4 [430 µmol/L] | 2.98 ± 0.40 | 73.69 ± 0.26 *** | 15.16 ± 0.54 *** | 8.21 ± 0.03 *** |
| Comp. 7 [55 µmol/L] | 17.85 ± 2.08 *** | 56.48 ± 3.34 *** | 10.34 ± 0.74 *** | 15.43 ± 0.93 *** |
| Control MDA-MB-231 | 1.96 ± 0.11 | 50.54 ± 0.86 | 28.88 ± 0.45 | 18.50 ± 0.45 |
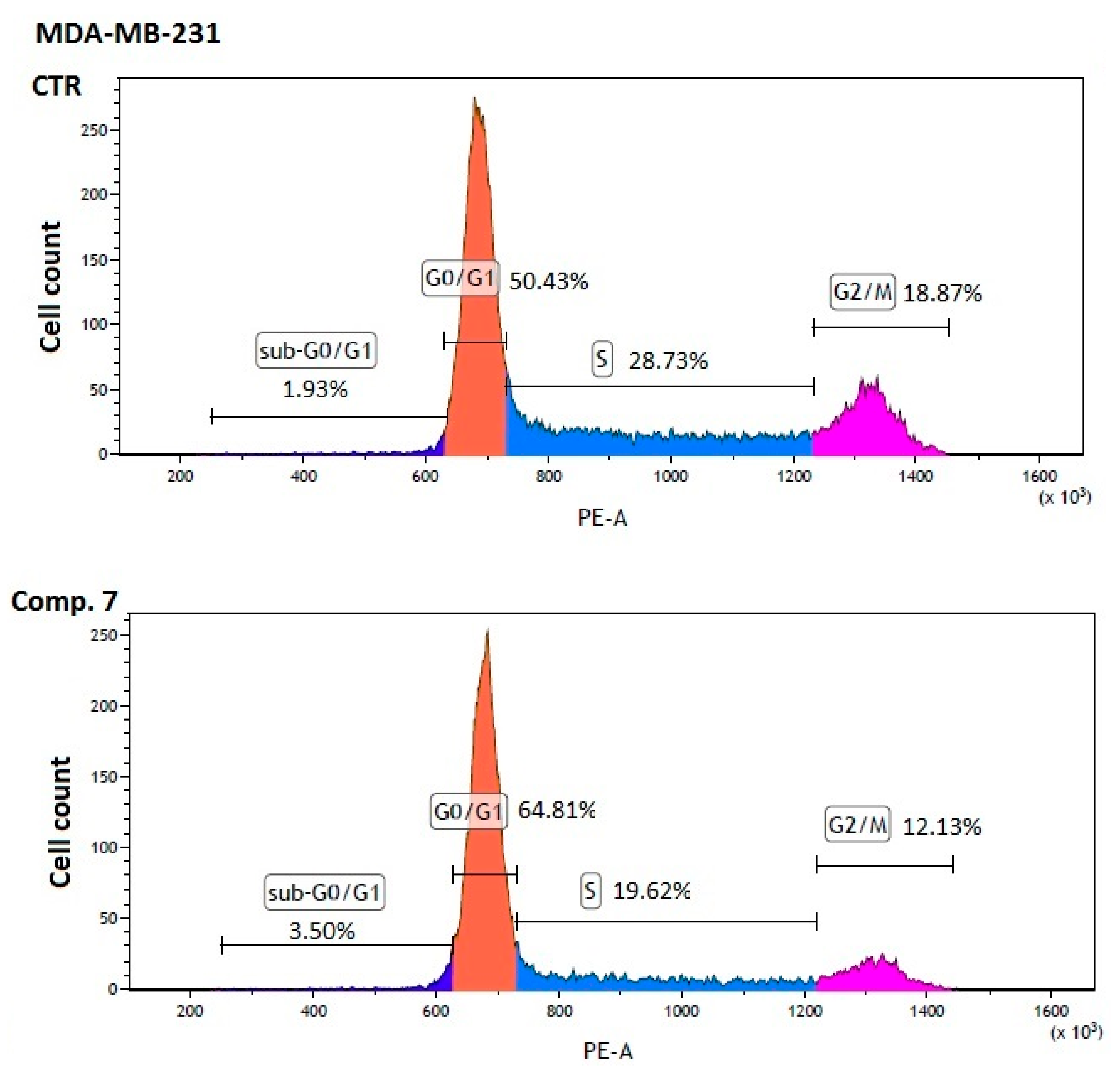
| Comp. 7 [2600 µmol/L] | 3.41 ± 0.08 ** | 64.31 ± 1.13 *** | 19.99 ± 0.97 ** | 12.24 ± 0.10 ** |
| MCF-7 | MDA-MB-231 | |||||
|---|---|---|---|---|---|---|
| GSH [nmol/1 mln Cells] | GSSG [nmol/1 mln Cells] | GSH:GSSG Ratio | GSH [nmol/1 mln Cells] | GSSG [nmol/1 mln Cells] | GSH:GSSG Ratio | |
| CTR | 147.1 ± 9.6 | 7.0 ± 0.9 | 21.9 | 204.3 ± 21.0 | 17.9 ± 1.3 | 11.4 |
| Ascorbic acid [200 µM] | 153.1 ± 5.4 | 6.0 ± 0.4 | 25.3 | 249.6 ± 20.8 | 17.9 ± 1.5 | 13.9 |
| AAPH [10 mM] | 117.6 ± 7.8 | 12.8 ± 0.5 | 8.9 | 102.7 ± 3.2 | 14.3 ± 1.9 | 6.9 |
| Comp. 1 [1/2 × IC50] | 167.5 ± 5.1 | 7.4 ± 1.8 | 22.6 | 242.0 ± 47.0 | 19.9 ± 2.0 | 12.1 |
| Comp. 1 [1/4 × IC50] | 183.1 ± 8.9 | 9.1 ± 3.9 | 20.1 | 238.0 ± 4.3 | 19.5 ± 2.2 | 12.2 |
| Comp. 4 [1/2 × IC50] | 148.5 ± 4.8 | 6.2 ± 0.7 | 24.0 | 242.1 ± 13.7 | 19.6 ± 1.4 | 12.3 |
| Comp. 4 [1/4 × IC50] | 165.4 ± 9.6 | 8.6 ± 0.7 | 19.2 | 250.2 ± 12.1 | 19.9 ± 2.3 | 12.5 |
| Comp. 7 [1/2 × IC50] | 169.0 ± 6.0 | 10.6 ± 0.4 | 15.9 | 248.7 ± 13.3 | 21.9 ± 2.9 | 11.3 |
| Comp. 7 [1/4 × IC50] | 166.8 ± 12.8 | 7.9 ± 0.9 | 21.2 | 265.0 ± 15.9 | 22.5 ± 0.9 | 11.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markowicz-Piasecka, M.; Komeil, I.; Huttunen, J.; Sikora, J.; Huttunen, K.M. Effective Cellular Transport of Ortho-Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells. Int. J. Mol. Sci. 2020, 21, 2389. https://doi.org/10.3390/ijms21072389
Markowicz-Piasecka M, Komeil I, Huttunen J, Sikora J, Huttunen KM. Effective Cellular Transport of Ortho-Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells. International Journal of Molecular Sciences. 2020; 21(7):2389. https://doi.org/10.3390/ijms21072389
Chicago/Turabian StyleMarkowicz-Piasecka, Magdalena, Ibrahim Komeil, Johanna Huttunen, Joanna Sikora, and Kristiina M. Huttunen. 2020. "Effective Cellular Transport of Ortho-Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells" International Journal of Molecular Sciences 21, no. 7: 2389. https://doi.org/10.3390/ijms21072389
APA StyleMarkowicz-Piasecka, M., Komeil, I., Huttunen, J., Sikora, J., & Huttunen, K. M. (2020). Effective Cellular Transport of Ortho-Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells. International Journal of Molecular Sciences, 21(7), 2389. https://doi.org/10.3390/ijms21072389