Exploratory Analysis of Circulating miRNA Signatures in Atrial Fibrillation Patients Determining Potential Biomarkers to Support Decision-Making in Anticoagulation and Catheter Ablation

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
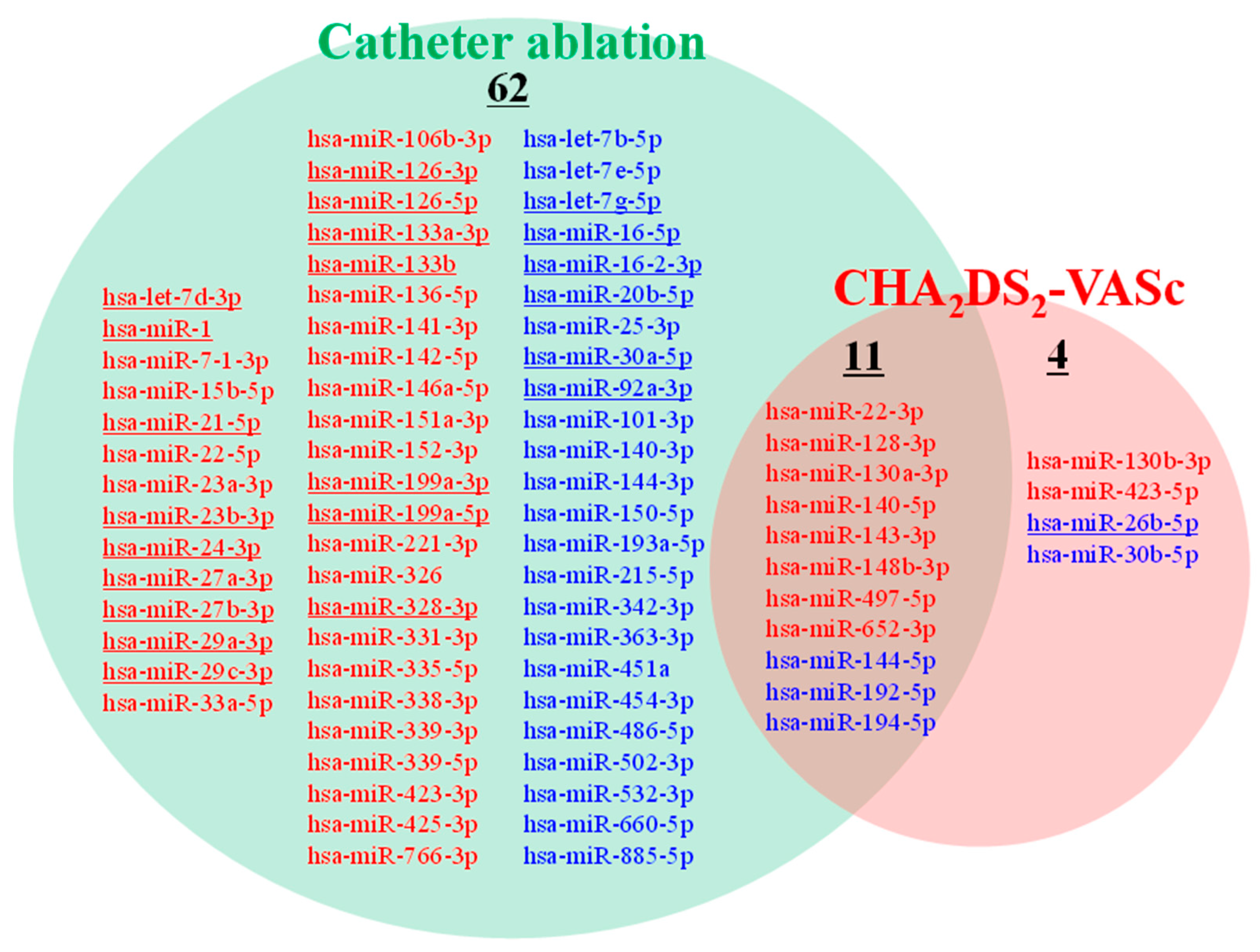
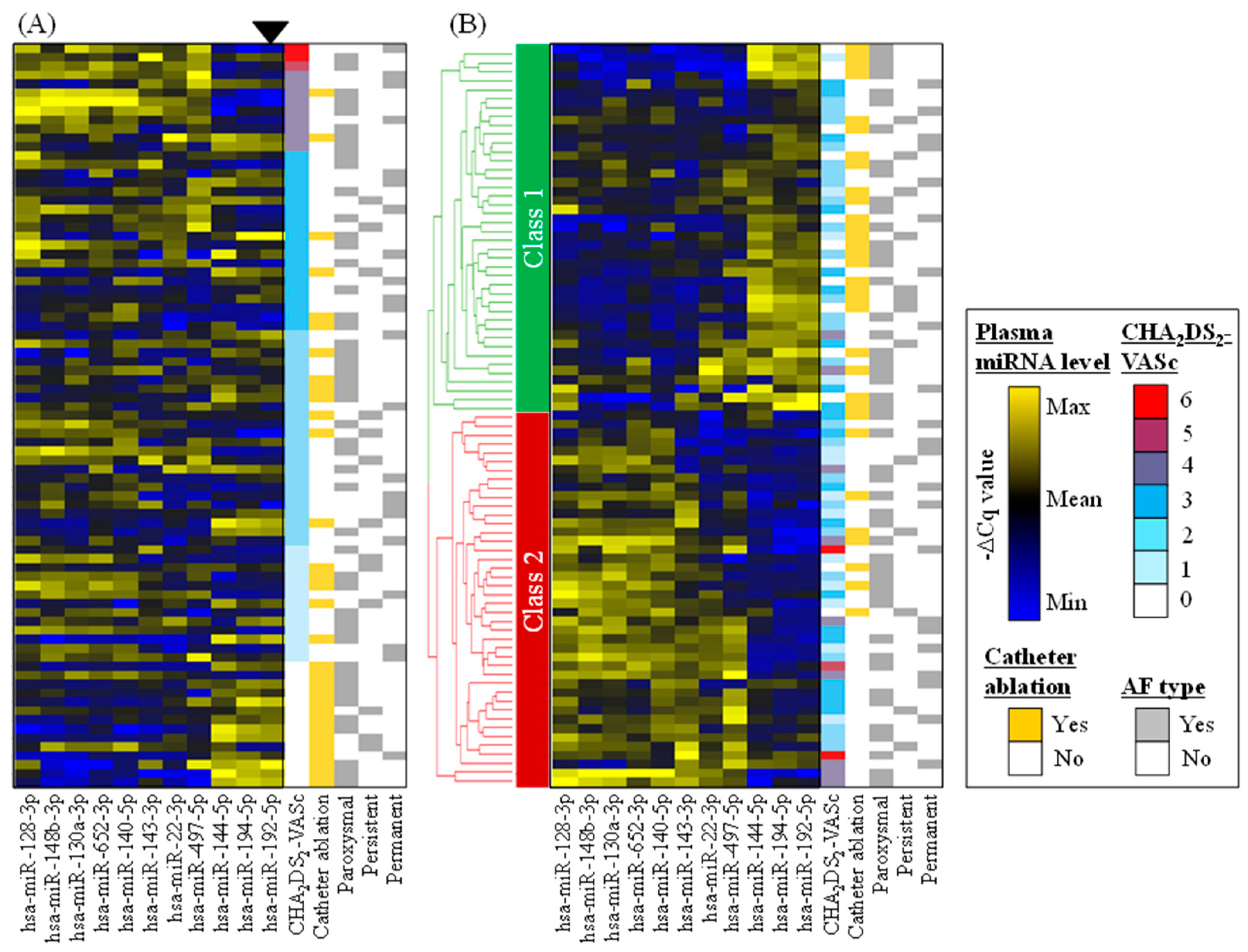
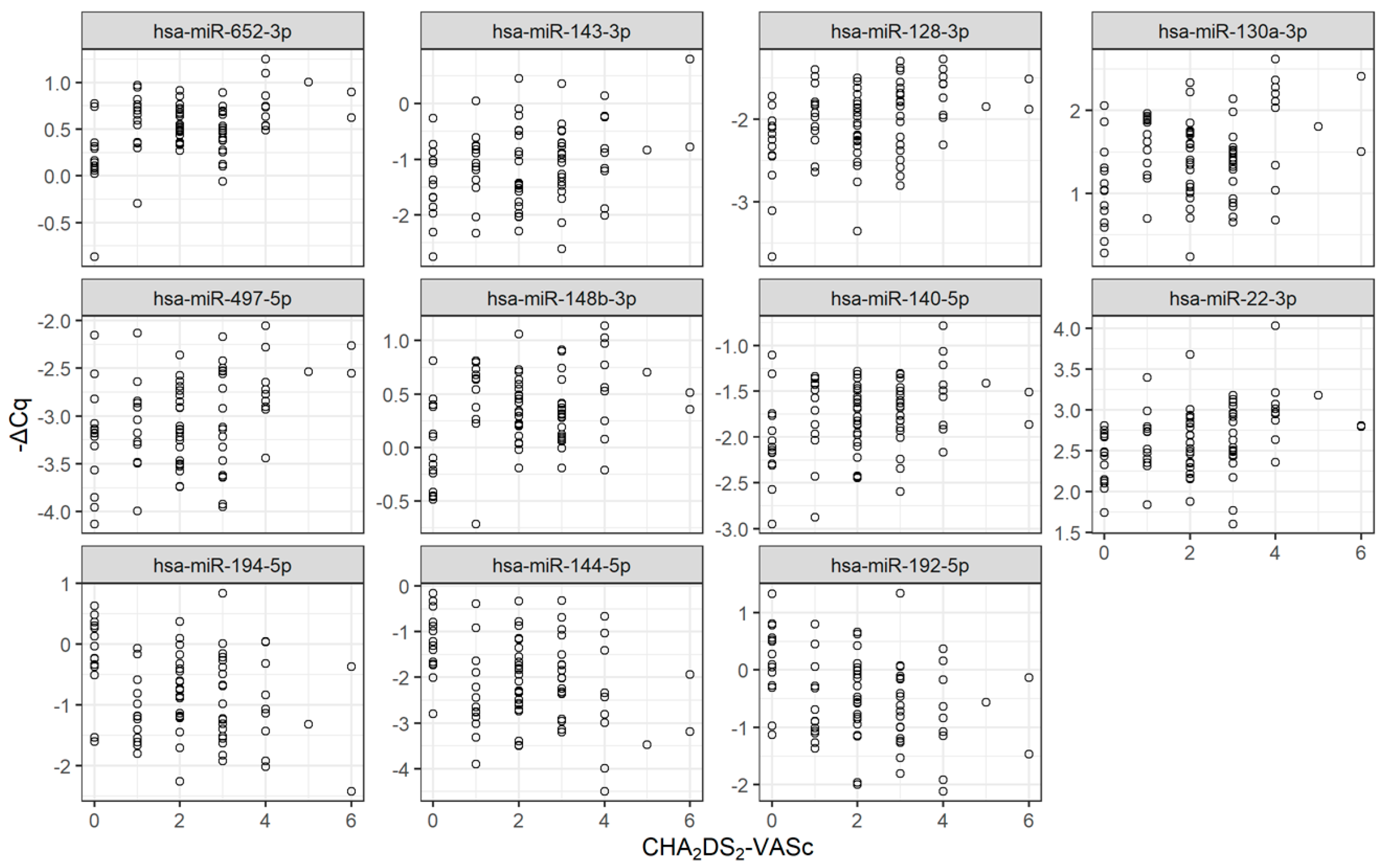
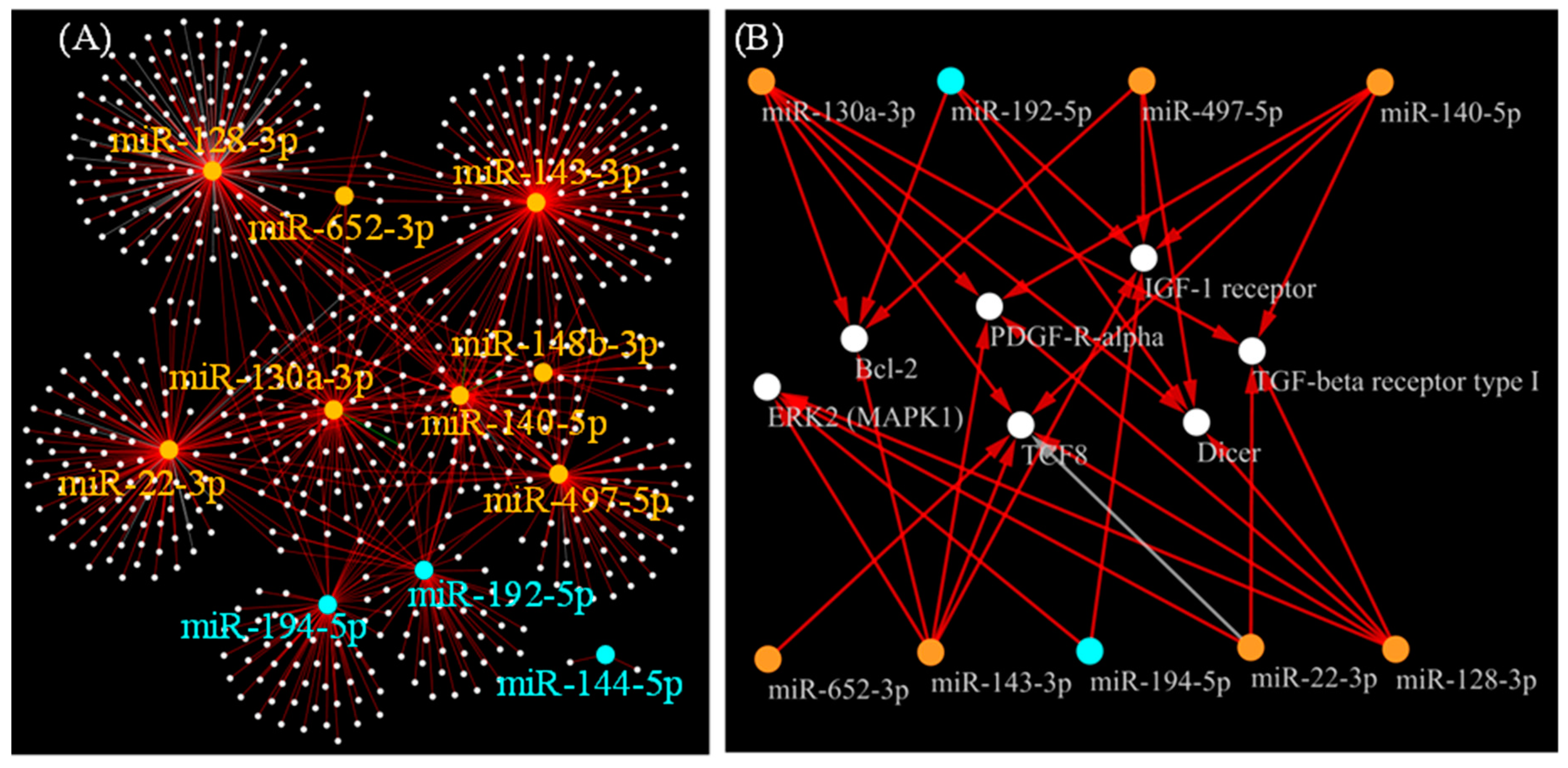
2.2. Identification of Circulating miRNAs Potentially Associated with Prognosis Risks
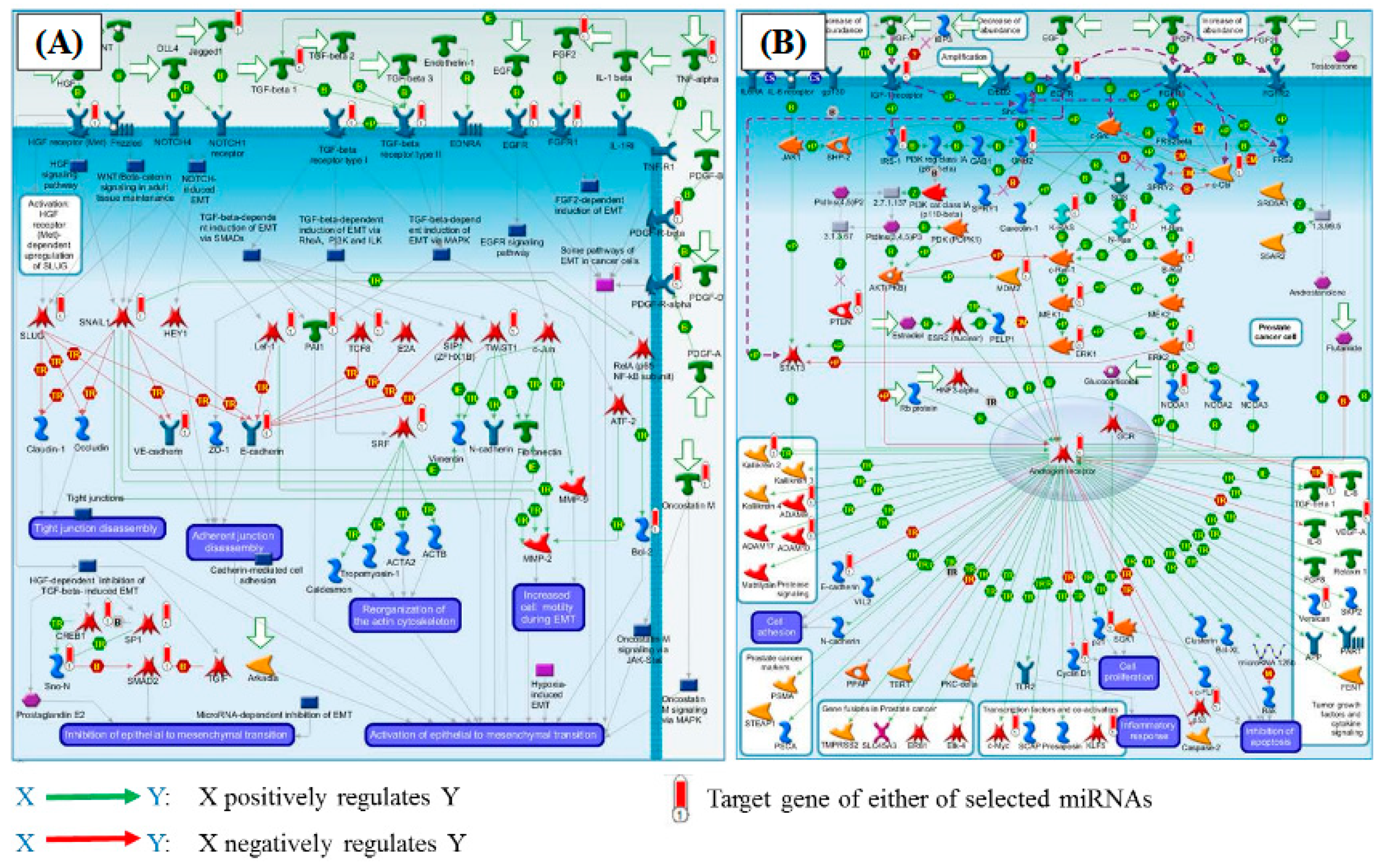
2.3. Pathway Enrichment Analysis
3. Discussion
3.1. Identification of 11 miRNAs Potentially Associated with Decision Making in AF Clinical Practice
3.2. Possible Functional Implication Relevant to 11 miRNAs
3.3. Possible Target Genes Strongly Regulated by 11 miRNAs
3.4. Clinical Implications: Classifying AF Patients Using 11 miRNAs Possibly Reflecting the Pro-Fibrotic State
3.5. Limitations
4. Materials and Methods
4.1. Ethics and Informed Consent
4.2. Study Population
4.3. RNA Extraction and miRNA Measurement
4.4. qRT-PCR Data Analysis
4.5. In Silico Functional Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | Atrial fibrillation |
| CA | Catheter ablation |
| EHRA | European Heart Rhythm Association |
| EndMT | Epithelial-to-mesenchymal transition |
| LAVI | Left atrial volume index |
| LVEF | Left ventricular ejection fraction |
| mRNA | Messenger RNA |
| miRNA | MicroRNA |
| PDGFRA | PDGF-R-alpha |
| TGFBR1 | TGF-β receptor type I |
References
- Schotten, U.; Verheule, S.; Kirchhof, P.; Goette, A. Pathophysiological Mechanisms of Atrial Fibrillation: A Translational Appraisal. Physiol. Rev. 2011, 91, 265–325. [Google Scholar] [CrossRef]
- De Jong, A.M.; Maass, A.H.; Oberdorf-Maass, S.U.; Van Veldhuisen, D.J.; Van Gilst, W.H.; Van Gelder, I.C. Mechanisms of atrial structural changes caused by stretch occurring before and during early atrial fibrillation. Cardiovasc. Res. 2010, 89, 754–765. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.M.; Van Gelder, I.C.; Vreeswijk-Baudoin, I.; Cannon, M.V.; Van Gilst, W.H.; Maass, A.H. Atrial Remodeling Is Directly Related to End-Diastolic Left Ventricular Pressure in a Mouse Model of Ventricular Pressure Overload. PLoS ONE 2013, 8, e72651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staerk, L.; Sherer, J.A.; Ko, D.; Benjamin, E.J.; Helm, R.H. Atrial Fibrillation: Epidemiology, Pathophysiology, and Clinical Outcomes. Circ. Res. 2017, 120, 1501–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, A.; Ardeshna, D.; Minhas, S.; Cave, B.; Ibeguogu, U.; Khouzam, R. Current status of oral anticoagulant reversal strategies: A review. Ann. Transl. Med. 2019, 7, 411. [Google Scholar] [CrossRef]
- Lip, G.Y.; Nieuwlaat, R.; Pisters, R.; Lane, D.A.; Crijns, H.J. Refining clinical risk stratification for predicting stroke and thromboembolism in atrial fibrillation using a novel risk factor-based approach: The euro heart survey on atrial fibrillation. Chest 2010, 137, 263–272. [Google Scholar] [CrossRef]
- Neefs, J.; Klamer, T.A.; Krul, S.P.J.; De Groot, J.R. Should Every Patient with Atrial Fibrillation and a CHA2DS2-VASc Score of 1 Be Anticoagulated? A Systematic Review of 37,030 Patients. Cardiol. Rev. 2019, 27, 249–255. [Google Scholar] [CrossRef]
- Kirchhof, P.; Benussi, S.; Kotecha, D.; Ahlsson, A.; Atar, D.; Casadei, B.; Castellà, M.; Diener, H.-C.; Heidbuchel, H.; Hendriks, J.; et al. 2016 ESC Guidelines for the management of atrial fibrillation developed in collaboration with EACTS. Eur. Heart J. 2016, 37, 2893–2962. [Google Scholar]
- Apostolakis, S.; Guo, Y.; Lane, D.A.; Buller, H.; Lip, G.Y.H. Renal function and outcomes in anticoagulated patients with non-valvular atrial fibrillation: The AMADEUS trial. Eur. Heart J. 2013, 34, 3572–3579. [Google Scholar] [CrossRef] [Green Version]
- Asad, Z.U.A.; Yousif, A.; Khan, M.S.; Al-Khatib, S.M.; Stavrakis, S. Catheter Ablation Versus Medical Therapy for Atrial Fibrillation. Circ. Arrhythmia Electrophysiol. 2019, 12, e007414. [Google Scholar] [CrossRef]
- Walsh, K.; Marchlinski, F.E. Catheter ablation for atrial fibrillation: Current patient selection and outcomes. Expert Rev. Cardiovasc. Ther. 2018, 16, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Ardhianto, P.; Yuniadi, Y. Biomarkers of atrial fibrillation: Which one is a true marker? Cardiol. Res. Pract. 2019, 2019, 8302326. [Google Scholar] [CrossRef]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santulli, G.; Iaccarino, G.; DeLuca, N.; Trimarco, B.; Condorelli, G. Atrial fibrillation and microRNAs. Front. Physiol. 2014, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Lu, Y.; Yang, B. MicroRNAs and atrial fibrillation: New fundamentals. Cardiovasc. Res. 2010, 89, 710–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lucia, C.; Komici, K.; Borghetti, G.; Femminella, G.D.; Bencivenga, L.; Cannavo, A.; Corbi, G.; Ferrara, N.; Houser, S.R.; Koch, W.J.; et al. microRNA in Cardiovascular Aging and Age-Related Cardiovascular Diseases. Front. Med. 2017, 4, 74. [Google Scholar]
- Lopez, J.P.; Diallo, A.; Cruceanu, C.; Fiori, L.M.; LaBoissière, S.; Guillet, I.; Fontaine, J.; Ragoussis, J.; Benes, V.; Turecki, G.; et al. Biomarker discovery: Quantification of microRNAs and other small non-coding RNAs using next generation sequencing. BMC Med. Genom. 2015, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Brase, J.C.; Wuttig, D.; Kuner, R.; Sueltmann, H. Serum microRNAs as non-invasive biomarkers for cancer. Mol. Cancer 2010, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Komal, S.; Yin, J.-J.; Wang, S.-H.; Huang, C.-Z.; Tao, H.-L.; Dong, J.-Z.; Han, S.-N.; Zhang, L. MicroRNAs: Emerging biomarkers for atrial fibrillation. J. Cardiol. 2019, 74, 475–482. [Google Scholar] [CrossRef]
- Van den Berg, N.W.E.; Kawasaki, M.; Berger, W.R.; Neefs, J.; Meulendijks, E.; Tijsen, A.J.; de Groot, J.R. MicroRNAs in Atrial Fibrillation: From Expression Signatures to Functional Implications. Cardiovasc. Drugs Ther. 2017, 31, 345–365. [Google Scholar] [CrossRef]
- Liang, Y.; Ridzon, D.; Wong, L.; Chen, C. Characterization of microRNA expression profiles in normal human tissues. BMC Genom. 2007, 8, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.A.; Lip, G.Y. Use of the CHA(2)DS(2)-VASc and HAS-BLED scores to aid decision making for thromboprophylaxis in nonvalvular atrial fibrillation. Circulation 2012, 126, 860–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burstein, B.; Nattel, S. Atrial Fibrosis: Mechanisms and Clinical Relevance in Atrial Fibrillation. J. Am. Coll. Cardiol. 2008, 51, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.H.; Yeh, Y.H.; Lee, J.L.; Hsu, Y.J.; Kuo, C.T.; Chen, W.J. Transforming growth factor-beta-mediated CD44/STAT3 signaling contributes to the development of atrial fibrosis and fibrillation. Basic Res. Cardiol. 2017, 112, 58. [Google Scholar] [CrossRef] [PubMed]
- Daccarett, M.; Badger, T.J.; Akoum, N.; Burgon, N.S.; Mahnkopf, C.; Vergara, G.; Kholmovski, E.; McGann, C.J.; Parker, D.; Brachmann, J.; et al. Association of Left Atrial Fibrosis Detected by Delayed-Enhancement Magnetic Resonance Imaging and the Risk of Stroke in Patients with Atrial Fibrillation. J. Am. Coll. Cardiol. 2011, 57, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Sekiguchi, A.; Sagara, K.; Tanabe, H.; Takamura, M.; Kaneko, S.; Aizawa, T.; Fu, L.-T.; Yamashita, T. Endothelial–mesenchymal transition in human atrial fibrillation. J. Cardiol. 2017, 69, 706–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, L.; Du, X.; You, T.; Sun, L.; Li, W.; Xiao, L.; Lu, H.; Wang, W.; Li, X. Reciprocal enhancement of thrombosis by endothelial-to-mesenchymal transition induced by iliac vein compression. Life Sci. 2019, 233, 116659. [Google Scholar] [CrossRef]
- Magnani, J.W.; Moser, C.B.; Murabito, J.M.; Sullivan, L.; Wang, N.; Ellinor, P.T.; Vasan, R.S.; Benjamin, E.J.; Coviello, A.D. Association of sex hormones, aging, and atrial fibrillation in men: The Framingham Heart Study. Circ. Arrhythmia Electrophysiol. 2014, 7, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Glueck, C.J.; Wang, P. Testosterone therapy, thrombosis, thrombophilia, cardiovascular events. Metabolism 2014, 63, 989–994. [Google Scholar] [CrossRef]
- Dzeshka, M.S.; Lip, G.Y.; Snezhitskiy, V.; Shantsila, E. Cardiac Fibrosis in Patients with Atrial Fibrillation: Mechanisms and Clinical Implications. J. Am. Coll. Cardiol. 2015, 66, 943–959. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, G.; Manabe, I.; Tsushima, K.; Fujiu, K.; Oishi, Y.; Imai, Y.; Maemura, K.; Miyagishi, M.; Higashi, Y.; Kondoh, H.; et al. DeltaEF1 mediates TGF-beta signaling in vascular smooth muscle cell differentiation. Dev. Cell 2006, 11, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, E.; Vidal, J.-S.; Funalot, B.; Brunel, N.; Viollet, C.; Seux, M.-L.; Treluyer, J.-M.; Epelbaum, J.; Le Bouc, Y.; Hanon, O. Insulin-Like Growth Factor I, Insulin-like Growth factor Binding Protein 3, and Atrial Fibrillation in the Elderly. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2013, 69, 1025–1032. [Google Scholar] [CrossRef] [Green Version]
- Saber, H.; Himali, J.J.; Beiser, A.; Shoamanesh, A.; Pikula, A.; Roubenoff, R.; Romero, J.R.; Kase, C.S.; Vasan, R.S.; Seshadri, S. Serum Insulin-Like Growth Factor 1 and the Risk of Ischemic Stroke: The Framingham Study. Stroke 2017, 48, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Goette, A.; Staack, T.; Röcken, C.; Arndt, M.; Geller, J.C.; Huth, C.; Ansorge, S.; Klein, H.U.; Lendeckel, U. Increased expression of extracellular signal-regulated kinase and angiotensin-converting enzyme in human atria during atrial fibrillation. J. Am. Coll. Cardiol. 2000, 35, 1669–1677. [Google Scholar] [CrossRef] [Green Version]
- Diao, S.-L.; Xu, H.-P.; Zhang, B.; Ma, B.-X.; Liu, X.-L. Associations of MMP-2, BAX, and Bcl-2 mRNA and Protein Expressions with Development of Atrial Fibrillation. Med. Sci. Monit. 2016, 22, 1497–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, S.; Vitacolonna, A.; Bonzano, A.; Comoglio, P.M.; Crepaldi, T. ERK: A Key Player in the Pathophysiology of Cardiac Hypertrophy. Int. J. Mol. Sci. 2019, 20, 2164. [Google Scholar] [CrossRef] [Green Version]
- Mutlak, M.; Schlesinger-Laufer, M.; Haas, T.; Shofti, R.; Ballan, N.; Lewis, Y.E.; Zuler, M.; Zohar, Y.; Caspi, L.H.; Kehat, I.; et al. Extracellular signal-regulated kinase (ERK) activation preserves cardiac function in pressure overload induced hypertrophy. Int. J. Cardiol. 2018, 270, 204–213. [Google Scholar] [CrossRef]
- Mayorga, M.; Bahi, N.; Ballester, M.; Comella, J.X.; Sanchís, D.; Junutula, J.R.; Schonteich, E.; Wilson, G.M.; Peden, A.A.; Scheller, R.H.; et al. Bcl-2 Is a Key Factor for Cardiac Fibroblast Resistance to Programmed Cell Death. J. Biol. Chem. 2004, 279, 34882–34889. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, M.; Xu, L.; Liu, J.; Wang, D.; Li, Q.; Wang, L.; Li, P.; Chen, S.; Liu, T. Expression of Bcl-2 and microRNAs in cardiac tissues of patients with dilated cardiomyopathy. Mol. Med. Rep. 2016, 15, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-F.; Murchison, E.P.; Tang, R.; Callis, T.E.; Tatsuguchi, M.; Deng, Z.; Rojas, M.; Hammond, S.M.; Schneider, M.D.; Selzman, C.H.; et al. Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure. Proc. Natl. Acad. Sci. USA 2008, 105, 2111–2116. [Google Scholar] [CrossRef] [Green Version]
- Japanese Circulation Society Joint Working Group. JCS/JHRS Guideline on Non-Pharmacotherapy of Cardiac Arrhythmias. 2018. (In Japanese). Available online: http://www.j-circ.or.jp/guideline/pdf/JCS2018_kurita_nogami.pdf (accessed on 12 March 2020).
- January, C.T.; Wann, L.S.; Alpert, J.S.; Calkins, H.; Cigarroa, J.E.; Cleveland, J.C.; Conti, J.B.; Ellinor, P.T.; Ezekowitz, M.D.; Field, M.E.; et al. 2014 AHA/ACC/HRS guideline for the management of patients with atrial fibrillation: A report of the American College of Cardiology/American Heart Association Task Force on practice guidelines and the Heart Rhythm Society. Circulation 2014, 130, e199–e267. [Google Scholar] [CrossRef]
- Lee, G.; Sparks, P.B.; Morton, J.B.; Kistler, P.M.; Vohra, J.K.; Medi, C.; Rosso, R.; Teh, A.; Halloran, K.; Kalman, J.M. Low Risk of Major Complications Associated with Pulmonary Vein Antral Isolation for Atrial Fibrillation: Results of 500 Consecutive Ablation Procedures in Patients with Low Prevalence of Structural Heart Disease From a Single Center. J. Cardiovasc. Electrophysiol. 2010, 22, 163–168. [Google Scholar]
- Rostock, T.; Salukhe, T.V.; Steven, D.; Drewitz, I.; Hoffmann, B.A.; Bock, K.; Servatius, H.; Müllerleile, K.; Sultan, A.; Gosau, N.; et al. Long-term single- and multiple-procedure outcome and predictors of success after catheter ablation for persistent atrial fibrillation. Heart Rhythm 2011, 8, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.N.; Zhang, Z.L.; Li, Z.; Zhang, C.; Li, H.; Wang, J.L.; Wang, J.; Gu, Z.C. Identification of microRNA biomarkers in atrial fibrillation: A protocol for systematic review and bioinformatics analysis. Medicine 2019, 98, e16538. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009, 10, R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiagen, Guidelines for Profiling Biofluid miRNAs. Available online: https://www.qiagen.com/us/-/media/project/qiagen/qiagen-home/documents/content-worlds/liquid-biopsy/guidelines-for-profiling-biofluid-mirnas.pdf (accessed on 11 March 2020).
- Godard, P.; van Eyll, J. Pathway analysis from lists of microRNAs: Common pitfalls and alternative strategy. Nucleic Acids Res. 2015, 43, 3490–3497. [Google Scholar] [CrossRef] [PubMed]
- Kiyosawa, N.; Manabe, S.; Yamoto, T.; Sanbuissho, A. Practical Application of Toxicogenomics for Profiling Toxicant-Induced Biological Perturbations. Int. J. Mol. Sci. 2010, 11, 3397–3412. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Morishima, Y.; Takita, A.; Yagi, N.; Otsuka, T.; Arita, T.; Yamashita, T. Responses of prothrombin time and activated partial thromboplastin time to edoxaban in Japanese patients with non-valvular atrial fibrillation: Characteristics of representative reagents in Japan (CVI ARO 7). Heart Vessels 2019, 34, 2011–2020. [Google Scholar] [CrossRef]
- Suzuki, S.; Morishima, Y.; Takita, A.; Otsuka, T.; Yagi, N.; Arita, T.; Yamashita, T. Association between plasma concentration of edoxaban determined by direct and indirect methods in Japanese patients with non-valvular atrial fibrillation (CVI ARO 7). Heart Vessels 2020, 35, 409–416. [Google Scholar] [CrossRef]
- Blondal, T.; Nielsen, S.J.; Baker, A.; Andreasen, D.; Mouritzen, P.; Teilum, M.W.; Dahlsveen, I.K. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods 2013, 59, S1–S6. [Google Scholar] [CrossRef]
- Costa Sa, A.C.C.; Madsen, H.; Brown, J.R. Shared Molecular Signatures Across Neurodegenerative Diseases and Herpes Virus Infections Highlights Potential Mechanisms for Maladaptive Innate Immune Responses. Sci. Rep. 2019, 9, 8795. [Google Scholar] [CrossRef] [PubMed]
- Godard, P.; Page, M. PCAN: Phenotype consensus analysis to support disease-gene association. BMC Bioinform. 2016, 17, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Arat, S.; Magid-Slav, M.; Brown, J.R. Meta-analysis of human gene expression in response to Mycobacterium tuberculosis infection reveals potential therapeutic targets. BMC Syst. Biol. 2018, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Risk Scores | n = 83 | % |
|---|---|---|
| Gender | ||
| Male | 61 | 73.5% |
| Female | 22 | 26.5% |
| Age (mean ± SD) | ||
| Male | 67.4 ± 11.4 | |
| Female | 74.0 ± 7.2 | |
| AF type | ||
| Paroxysmal | 48 | 57.8% |
| Persistent | 13 | 15.7% |
| Permanent | 22 | 26.5% |
| Number of Patients Underwent Catheter Ablation | ||
| Paroxysmal | 22 | 26.5% |
| Persistent | 9 | 10.8% |
| Permanent | 1 | 1.2% |
| CHADS2 Score | ||
| 0 | 25 | 30.1% |
| 1 | 33 | 39.8% |
| 2 | 15 | 18.1% |
| 3 | 7 | 8.4% |
| 4 | 3 | 3.6% |
| >5 | 0 | 0% |
| CHA2DS2-VASc Score | ||
| 0 | 14 | 16.9% |
| 1 | 13 | 15.7% |
| 2 | 24 | 28.9% |
| 3 | 20 | 24.1% |
| 4 | 9 | 10.8% |
| 5 | 1 | 1.2% |
| 6 | 2 | 2.4% |
| >7 | 0 | 0% |
| HAS-BLED Score | ||
| 0 | 17 | 20.5% |
| 1 | 25 | 30.1% |
| 2 | 30 | 36.1% |
| 3 | 11 | 13.3% |
| >4 | 0 | 0% |
| Total (n = 33) | Paroxysmal AF (n = 22) | Persistent AF (n = 10) | Permanent AF (n = 1) | |
|---|---|---|---|---|
| Age, years | 61.3 ± 10.7 | 62.3 ± 11.1 | 59.7 ± 10.3 | 56 |
| Male, n (%) | 28 (85) | 19 (86) | 8 (80) | 1 |
| CHA2DS2-VASc, points | 1 (0–2) | 1 (0–2) | 1 (0–2) | 0 |
| EHRA Score | ||||
| 1 | 12 (36) | 6 (27) | 5 (50) | 1 |
| 2 | 16 (49) | 12 (55) | 4 (40) | 0 |
| 3 | 2 (6) | 1 (5) | 1 (10) | 0 |
| 4 | 3 (9) | 3 (14) | 0 (0) | 0 |
| Mitral regurgitation, n (%) | 0 (0) | 0 (0) | 0 (0) | 0 |
| Mitral stenosis, n (%) | 0 (0) | 0 (0) | 0 (0) | 0 |
| Aortic regurgitation, n (%) | 0 (0) | 0 (0) | 0 (0) | 0 |
| Aortic stenosis, n (%) | 1 (3) | 1 (5) | 0 (0) | 0 |
| LVEF, % | 63.3 ± 8.8 | 63.9 ± 9.9 | 61.3 ± 5.8 | 72 |
| LAVI, mL/m2 | 39.3 ± 17.7 | 38.8 ± 20.4 | 37.7 ± 10.9 | 61 |
| Recurrence within 1 year | 1 (3) | 1 (5) | 0 (0) | 0 |
| Pathways | Number of Mapped Genes | Number of Genes in the Pathway | q-Value |
|---|---|---|---|
| Development_regulation of epithelial-to-mesenchymal transition (EMT) | 26 | 64 | 7.8 × 10−20 |
| Androgen receptor activation and downstream signaling in prostate cancer | 31 | 110 | 8.0 × 10−19 |
| Development_TGF-β receptor signaling | 23 | 52 | 8.0 × 10−19 |
| Dual role of TGF-β 1 in HCC | 17 | 24 | 1.7 × 10−18 |
| Development_YAP/TAZ-mediated co-regulation of transcription | 23 | 56 | 3.8 × 10−18 |
| TGF-β signaling via SMADs in breast cancer | 21 | 47 | 1.8 × 10−17 |
| Main genetic and epigenetic alterations in lung cancer | 21 | 48 | 2.7 × 10−17 |
| Cell cycle_Regulation of G1/S transition (part 1) | 19 | 38 | 5.4 × 10−17 |
| TGF-β 1-mediated induction of EMT in normal and asthmatic airway epithelium | 20 | 44 | 6.1 × 10−17 |
| Mechanisms of resistance to EGFR inhibitors in lung cancer | 20 | 45 | 9.6 × 10−17 |
| Ligand-independent activation of androgen receptor in prostate cancer | 23 | 67 | 2.1 × 10−16 |
| Inhibition of TGF-β signaling in lung cancer | 17 | 31 | 3.5 × 10−16 |
| Activation of TGF-β signaling in pancreatic cancer | 16 | 27 | 5.9 × 10−16 |
| K-RAS signaling in pancreatic cancer | 19 | 44 | 1.0 × 10−15 |
| Main pathways of Schwann cells transformation in neurofibromatosis type 1 | 24 | 80 | 1.0 × 10−15 |
| TGF-β signaling via kinase cascades in breast cancer | 21 | 58 | 1.3 × 10−15 |
| Role of microRNAs in cell migration, survival and angiogenesis in colorectal cancer | 22 | 67 | 2.4 × 10−15 |
| Stromal-epithelial interaction in prostate cancer | 18 | 42 | 7.0 × 10−14 |
| EGFR family signaling in pancreatic cancer | 22 | 75 | 3.2 × 10−14 |
| Role of microRNAs in cell proliferation in colorectal cancer | 21 | 69 | 6.2 × 10−14 |
| Pathways | Number of Mapped Genes | Number of Genes in the Pathway | q-Value |
|---|---|---|---|
| Androgen receptor activation and downstream signaling in prostate cancer | 11 | 110 | 2.1× 10−7 |
| TGF-β signaling via kinase cascades in breast cancer | 8 | 58 | 2.9 × 10−6 |
| Mitogenic action of ESR1 (membrane) in breast cancer | 7 | 47 | 9.7 × 10−6 |
| Main genetic and epigenetic alterations in lung cancer | 7 | 48 | 9.7 × 10−6 |
| IGF-1 signaling in multiple myeloma | 7 | 50 | 1.0 × 10−5 |
| SHH signaling in melanoma | 6 | 33 | 1.3 × 10−5 |
| IGF family, invasion and metastasis in colorectal cancer | 6 | 33 | 1.3 × 10−5 |
| Aberrant B-Raf signaling in melanoma progression | 7 | 55 | 1.3 × 10−5 |
| Signal transduction_AKT signaling | 6 | 43 | 5.2 × 10−5 |
| Mechanisms of drug resistance in SCLC | 7 | 70 | 5.2 × 10−5 |
| Development_Ligand-independent activation of ESR1 and ESR2 | 6 | 44 | 5.2 × 10−5 |
| Stem cells_stimulation of differentiation of mouse embryonic fibroblasts into adipocytes by extracellular factors | 7 | 71 | 5.2 × 10−5 |
| Immune response_IL-2 signaling via ERK, PI3K, and PLC-γ | 7 | 73 | 5.8 × 10−5 |
| Neuroendocrine transdifferentiation in prostate cancer | 6 | 48 | 7.5 × 10−5 |
| The role of PTEN and PI3K signaling in melanoma | 6 | 50 | 9.0 × 10−5 |
| Anti-apoptotic action of ErbB2 in breast cancer | 6 | 51 | 9.5 × 10−5 |
| Stem cells role of growth factors in the maintenance of embryonic stem cell pluripotency | 6 | 53 | 1.1 × 10−4 |
| DNA damage_Brca1 as a transcription regulator | 5 | 30 | 1.2 × 10−4 |
| Cell adhesion_ECM remodeling | 6 | 55 | 1.3 × 10−4 |
| Putative role of estrogen and androgen receptor signaling in progression of lung cancer | 6 | 58 | 1.6 × 10−4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiyosawa, N.; Watanabe, K.; Morishima, Y.; Yamashita, T.; Yagi, N.; Arita, T.; Otsuka, T.; Suzuki, S. Exploratory Analysis of Circulating miRNA Signatures in Atrial Fibrillation Patients Determining Potential Biomarkers to Support Decision-Making in Anticoagulation and Catheter Ablation. Int. J. Mol. Sci. 2020, 21, 2444. https://doi.org/10.3390/ijms21072444
Kiyosawa N, Watanabe K, Morishima Y, Yamashita T, Yagi N, Arita T, Otsuka T, Suzuki S. Exploratory Analysis of Circulating miRNA Signatures in Atrial Fibrillation Patients Determining Potential Biomarkers to Support Decision-Making in Anticoagulation and Catheter Ablation. International Journal of Molecular Sciences. 2020; 21(7):2444. https://doi.org/10.3390/ijms21072444
Chicago/Turabian StyleKiyosawa, Naoki, Kenji Watanabe, Yoshiyuki Morishima, Takeshi Yamashita, Naoharu Yagi, Takuto Arita, Takayuki Otsuka, and Shinya Suzuki. 2020. "Exploratory Analysis of Circulating miRNA Signatures in Atrial Fibrillation Patients Determining Potential Biomarkers to Support Decision-Making in Anticoagulation and Catheter Ablation" International Journal of Molecular Sciences 21, no. 7: 2444. https://doi.org/10.3390/ijms21072444
APA StyleKiyosawa, N., Watanabe, K., Morishima, Y., Yamashita, T., Yagi, N., Arita, T., Otsuka, T., & Suzuki, S. (2020). Exploratory Analysis of Circulating miRNA Signatures in Atrial Fibrillation Patients Determining Potential Biomarkers to Support Decision-Making in Anticoagulation and Catheter Ablation. International Journal of Molecular Sciences, 21(7), 2444. https://doi.org/10.3390/ijms21072444