Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro


Abstract
:1. Introduction
2. Results and Discussion
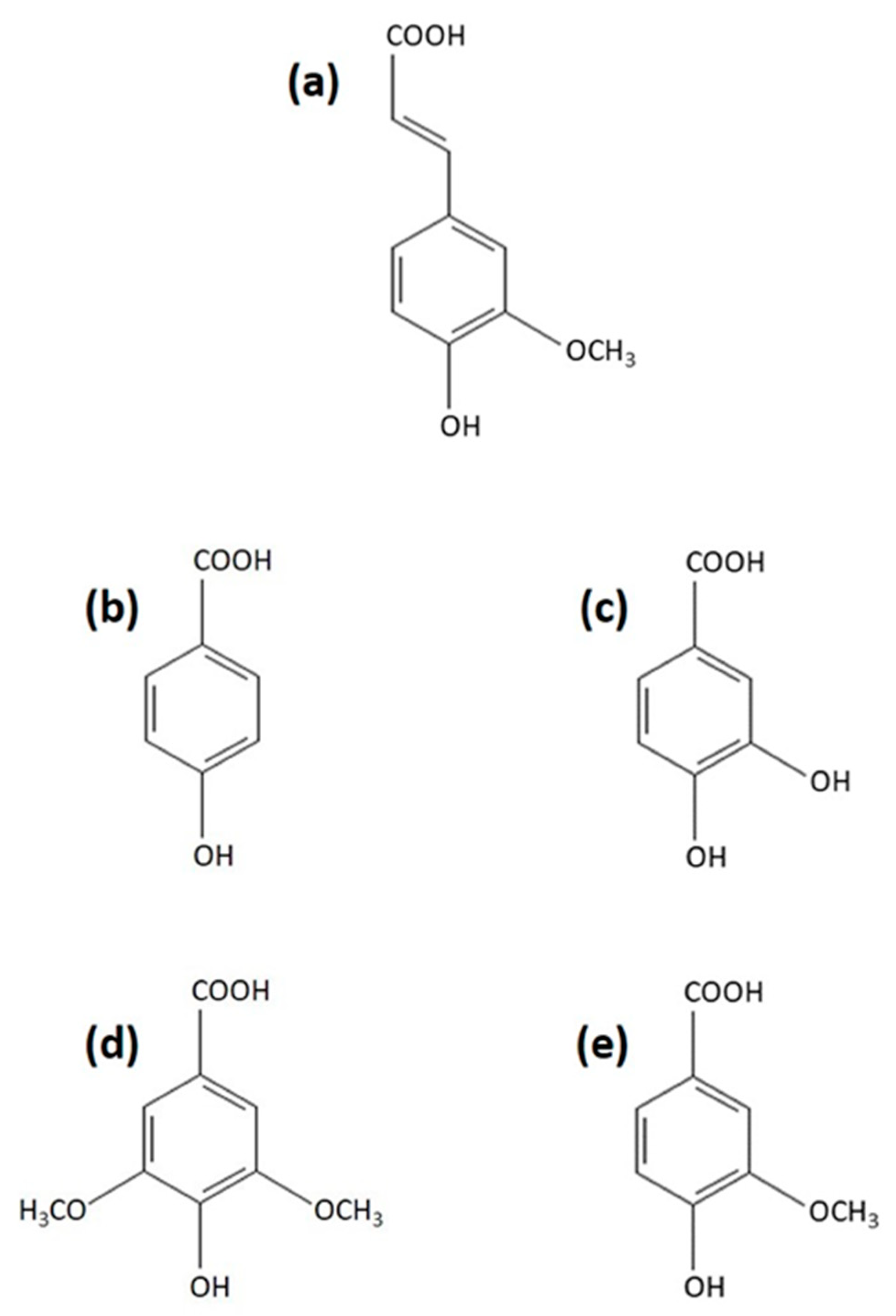
2.1. Comparison of the Effect of Various Phenolic Acids on CHTL Activity of T. versicolor Proteasomes
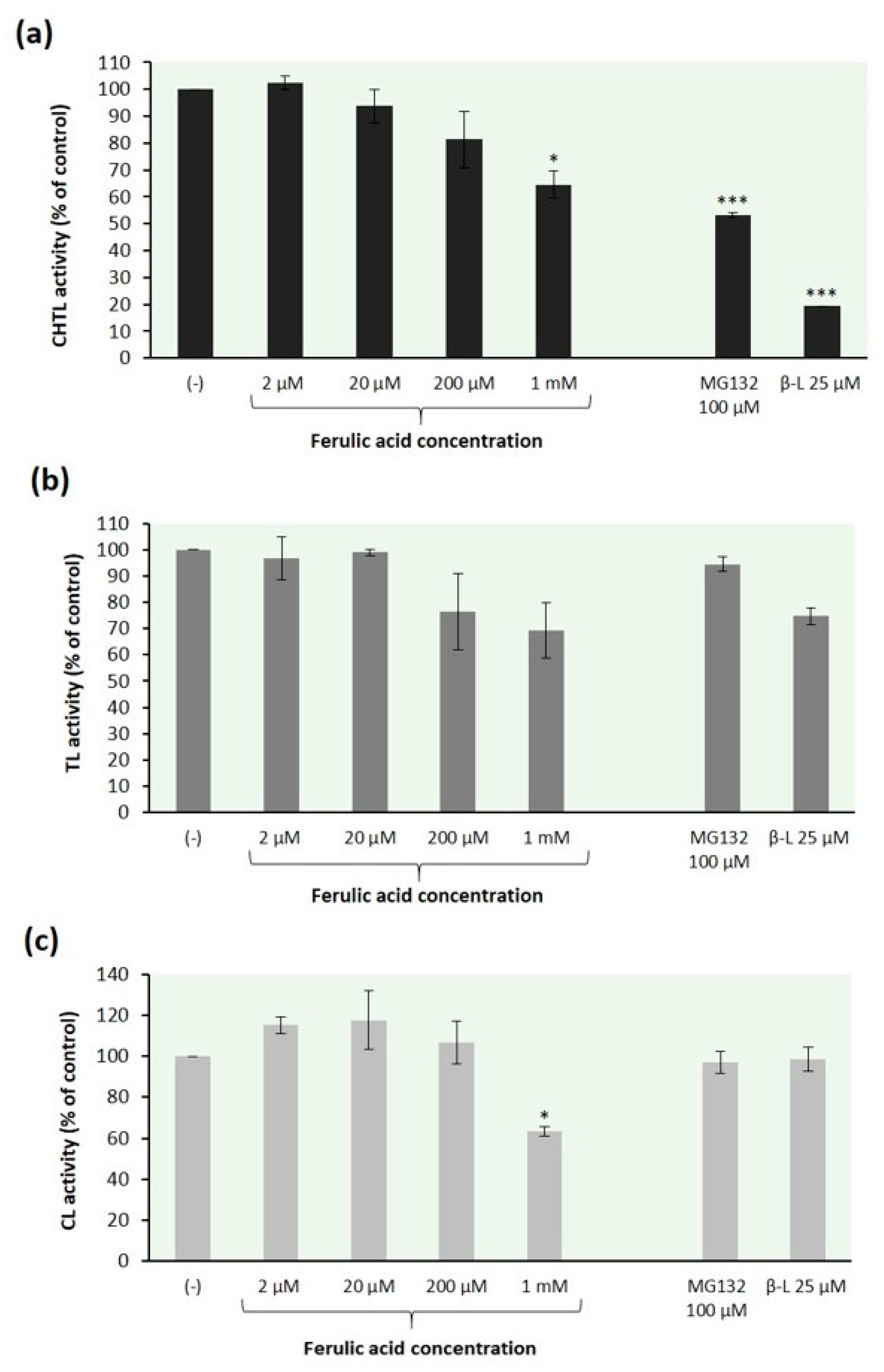
2.2. Effect of Ferulic Acid on 26S Proteasome Peptidase Activities
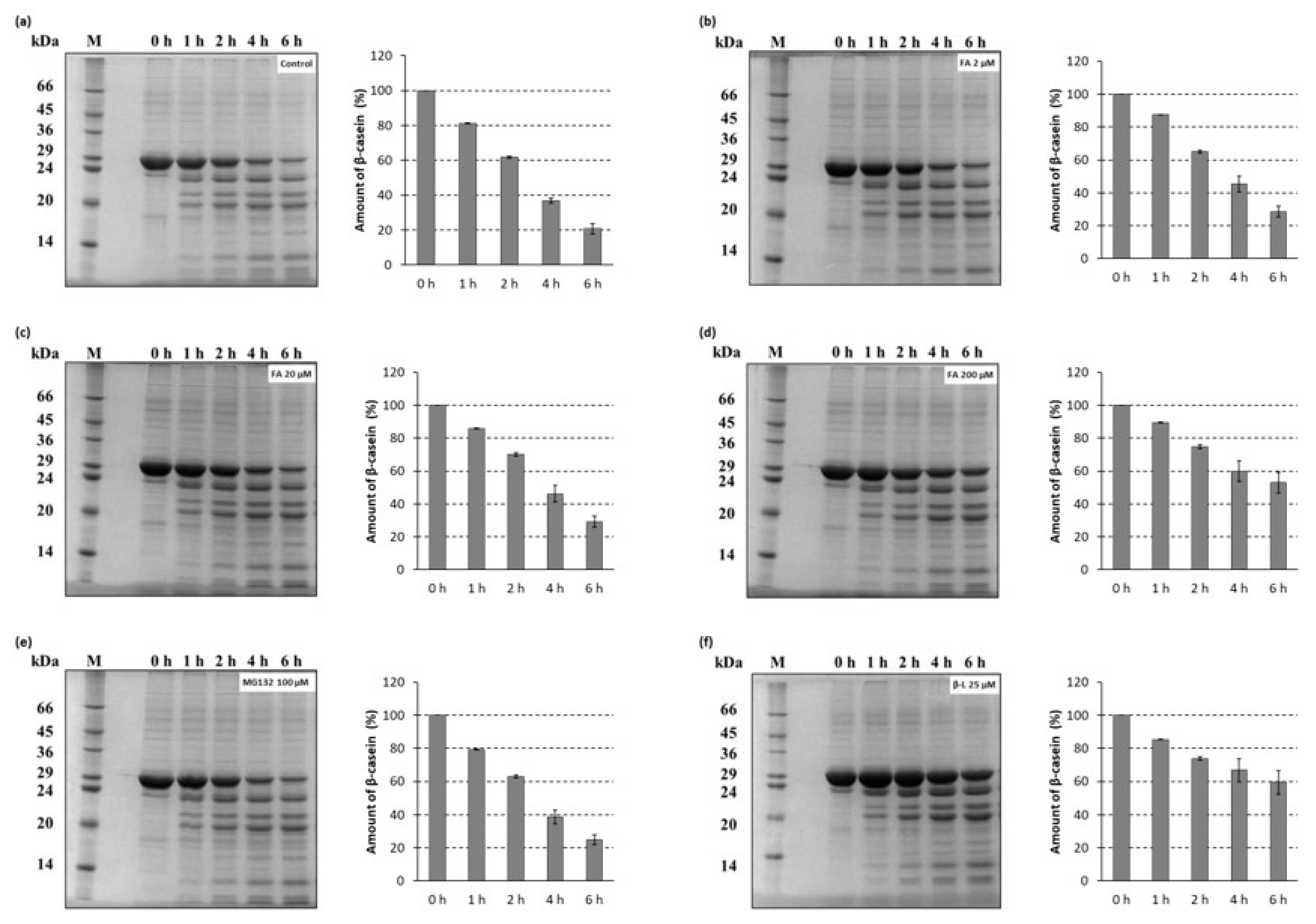
2.3. Degradation of a Full-Length Protein Substrate by 26S Fungal Proteasomes
3. Materials and Methods
3.1. Materials
3.2. Organism and Culture Conditions
3.3. Preparation of Crude Mycelium Extract
3.4. Isolation of 26S Proteasomes
3.5. Assays of 26S Proteasome Peptidase Activities
3.6. Degradation of a Full-Length Protein Substrate by 26S Proteasomes
3.6.1. Fluorescamine Assay
3.6.2. SDS-PAGE
3.7. Protein Determination
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMC | 7-amino-4-methylcoumarin |
| CHTL | chymotrypsin-like |
| CL | caspase-like |
| DMSO | dimethyl sulfoxide |
| MG132 | Z-Leu-Leu-Leu-al |
| Suc-LLVY-AMC | Suc-Leu-Leu-Val-Tyr-7-amido-4-methylcoumarin |
| TL | trypsin-like |
| UPP | ubiquitin-proteasome pathway |
| Z-GGR-AMC | Z-Gly-Gly-Arg-7-amido-4-methylcoumarin |
| Z-LLE-AMC | Z-Leu-Leu-Glu-amido-4-methylcoumarin |
References
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M.; Cohen, R.E. Proteasomes and their kin: Proteases in the machine age. Nat. Rev. Mol. Cell Biol. 2004, 5, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Gallastegui, N.; Groll, M. The 26S proteasome: Assembly and function of a destructive machine. Trends Biochem. Sci. 2010, 35, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Bedford, L.; Paine, S.; Sheppard, P.W.; Mayer, R.J.; Roelofs, J. Assembly, structure, and function of the 26S proteasome. Trends Cell Biol. 2010, 20, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wlodawer, A. Proteasome: A complex protease with a new fold and a distinct mechanism. Structure 1995, 3, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Kunjappu, M.J.; Hochstrasser, M. Assembly of the 20S proteasome. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Lander, G.C.; Estrin, E.; Matyskiela, M.E.; Bashore, C.; Nogales, E.; Martin, A. Complete subunit architecture of the proteasome regulatory particle. Nature 2012, 482, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Orlowski, M.; Wilk, S. Catalytic activities of the 20 S proteasome, a multicatalytic proteinase complex. Arch. Biochem. Biophys. 2000, 383, 1–16. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Callard, A.; Goldberg, A.L. Importance of the different proteolytic sites of the proteasome and the efficacy of inhibitors varies with the protein substrate. J. Biol. Chem. 2006, 281, 8582–8590. [Google Scholar] [CrossRef] [Green Version]
- Seemüller, E.; Lupas, A.; Stock, D.; Löwe, J.; Huber, R.; Baumeister, W. Proteasome from Thermoplasma acidophilum: A threonine protease. Science 1995, 268, 579–582. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Songyang, Z.; Goldberg, A.L. Why does threonine, and not serine, function as the active site nucleophile in proteasomes? J. Biol. Chem. 2000, 275, 14831–14837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groll, M.; Huber, R. Substrate access and processing by the 20S proteasome core particle. Int. J. Biochem. Cell Biol. 2003, 35, 606–616. [Google Scholar] [CrossRef]
- Thrower, J.S.; Hoffman, L.; Rechsteiner, M.; Pickart, C.M. Recognition of the polyubiquitin proteolytic signal. EMBO J. 2000, 19, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciechanover, A. Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell Biol. 2005, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.L. Functions of the proteasome: From protein degradation and immune surveillance to cancer therapy. Biochem. Soc. Trans. 2007, 35, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahlmann, B. Role of proteasomes in disease. BMC Biochem. 2007, 8, S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroski, M.D. The ubiquitin system, disease, and drug discovery. BMC Biochem. 2008, 9, S7. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Finley, D. Regulation of proteasome activity in health and disease. Biochim. Biophys. Acta 2014, 1843, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Bibo-Verdugo, B.; Jiang, Z.; Caffrey, C.R.; O’Donoghue, A.J. Targeting proteasomes in infectious organisms to combat disease. FEBS J. 2017, 284, 1503–1517. [Google Scholar] [CrossRef] [Green Version]
- Adams, J. Proteasome inhibition: A novel approach to cancer therapy. Trends Mol. Med. 2002, 8, S49–S54. [Google Scholar] [CrossRef]
- de Bettignies, G.; Coux, O. Proteasome inhibitors: Dozens of molecules and still counting. Biochimie 2010, 92, 1530–1545. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; van der Linden, W.A.; Overkleeft, H.S. Proteasome inhibitors: An expanding army attacking a unique target. Chem. Biol. 2012, 19, 99–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, E.M.; Groll, M. Inhibitors for the immuno- and constitutive proteasome: Current and future trends in drug development. Angew. Chem. Int. Ed. 2012, 51, 8708–8720. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Groettrup, M. Subunit specific inhibitors of proteasomes and their potential for immunomodulation. Curr. Opin. Chem. Biol. 2014, 23, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, D.L.; Crews, C.M. Small-molecule control of intracellular protein levels through modulation of the ubiquitin proteasome system. Angew. Chem. Int. Ed. 2014, 53, 2312–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staszczak, M. Ubiquitin-proteasome pathway as a target for therapeutic strategies. Postepy Biochem. 2017, 63, 287–303. [Google Scholar]
- Allegra, A.; Alonci, A.; Gerace, D.; Russo, S.; Innao, V.; Calabrò, L.; Musolino, C. New orally active proteasome inhibitors in multiple myeloma. Leuk. Res. 2014, 38, 1–9. [Google Scholar] [CrossRef]
- Buac, D.; Shen, M.; Schmitt, S.; Kona, F.R.; Deshmukh, R.; Zhang, Z.; Neslund-Dudas, C.; Mitra, B.; Dou, Q.P. From bortezomib to other inhibitors of the proteasome and beyond. Curr. Pharm. Des. 2013, 19, 4025–4038. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.L.; Groll, M. Applied techniques for mining natural proteasome inhibitors. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Momose, I.; Kawada, M. The therapeutic potential of microbial proteasome inhibitors. Int. Immunopharmacol. 2016, 37, 23–30. [Google Scholar] [CrossRef]
- Vierstra, R.D. The ubiquitin/26S proteasome pathway, the complex last chapter in the life of many plant proteins. Trends Plant Sci. 2003, 8, 135–142. [Google Scholar] [CrossRef]
- Fischer, M.; Hilt, W.; Richter-Ruoff, B.; Gonen, H.; Ciechanover, A.; Wolf, D.H. The 26S proteasome of the yeast Saccharomyces cerevisiae. FEBS Lett. 1994, 355, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Staszczak, M.; Jarosz-Wilkołazka, A. Inhibition of the proteasome strongly affects cadmium stimulated laccase activity in Trametes versicolor. Biochimie 2005, 87, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Staszczak, M. The 26S proteasome of the lignin-degrading Basidiomycete Phlebia radiata. Enzyme Microb. Technol. 2007, 40, 347–353. [Google Scholar] [CrossRef]
- Staszczak, M. Proteasomal degradation pathways in Trametes versicolor and Phlebia radiata. Enzyme Microb. Technol. 2002, 30, 537–541. [Google Scholar] [CrossRef]
- Staszczak, M. The role of the ubiquitin–proteasome system in the response of the ligninolytic fungus Trametes versicolor to nitrogen deprivation. Fungal Genet. Biol. 2008, 45, 328–337. [Google Scholar] [CrossRef]
- Staszczak, M.; Sajewicz, J.; Ohga, S. Effect of endoplasmic reticulum stress on laccase production and the 26S proteasome activity in the white rot fungus Trametes versicolor. J. Fac. Agric. Kyushu Univ. 2011, 56, 205–211. [Google Scholar]
- Hatakka, A. Lignin-modifying enzymes from selected white-rot fungi: Production and role from in lignin degradation. FEMS Microbiol. Rev. 1994, 13, 125–135. [Google Scholar] [CrossRef]
- Janusz, G.; Kucharzyk, K.H.; Pawlik, A.; Staszczak, M.; Paszczynski, A.J. Fungal laccase, manganese peroxidase and lignin peroxidase: Gene expression and regulation. Enzym. Microb. Technol. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell. Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Kenealy, W.R.; Jeffries, T.W. Enzyme processes for pulp and paper: A review of recent developments. ACS Symp. Ser. 2003, 845, 210–239. [Google Scholar] [CrossRef]
- Paszczynski, A.; Crawford, R.L. Recent advances in the use of fungi in environmental remediation and biotechnology. In Soil Biochemistry; Bollag, J.-M., Stotzky, G., Eds.; Marcel Dekker: New York, NY, USA, 2000; Volume 10, pp. 379–422. [Google Scholar]
- Jarosz-Wilkołazka, A.; Ruzgas, T.; Gorton, L. Use of laccase-modified electrode for amperometric detection of plant flavonoids. Enzym. Microb. Technol. 2004, 35, 238–241. [Google Scholar] [CrossRef]
- Stolarczyk, K.; Sepelowska, M.; Lyp, D.; Żelechowska, K.; Biernat, J.F.; Rogalski, J.; Farmer, K.D.; Roberts, K.N.; Bilewicz, R. Hybrid biobattery based on arylated carbon nanotubes and laccase. Bioelectrochemistry 2012, 87, 154–163. [Google Scholar] [CrossRef]
- Matuszewska, A.; Karp, M.; Jaszek, M.; Janusz, G.; Osińska-Jaroszuk, M.; Sulej, J.; Stefaniuk, D.; Tomczak, W.; Giannopoulos, K. Laccase purified from Cerrena unicolor exerts antitumor activity against leukemic cells. Oncol. Lett. 2016, 11, 2009–2018. [Google Scholar] [CrossRef] [Green Version]
- Jasim, A. Medicinal properties of laccase from Basidiomycetes mushroom: A review. Adv. Life Sci. Technol. 2017, 54, 99–109. [Google Scholar]
- Leonowicz, A.; Trojanowski, J. Induction of laccase by ferulic acid in Basidiomycetes. Acta Biochim. Pol. 1975, 22, 291–295. [Google Scholar]
- Gianfreda, L.; Xu, F.; Bollag, J.M. Laccases: A useful group of oxidoreductive enzymes. Bioremediat. J. 1999, 3, 1–25. [Google Scholar] [CrossRef]
- Leonowicz, A.; Trojanowski, J.; Orlicz, B. Induction of laccase in Basidiomycetes: Apparent activity of the inducible and constitutive forms of the enzyme with phenolic substrates. Acta Biochim. Pol. 1978, 25, 369–378. [Google Scholar]
- Cho, N.S.; Kim, D.H.; Cho, H.Y.; Ohga, S.; Leonowicz, A. Effect of various compounds on the activity of laccases from Basidiomycetes and their oxidative and demethoxylating activities. J. Fac. Agric. Kyushu Univ. 2006, 51, 211–218. [Google Scholar]
- De Souza, C.G.M.; Tychanowicz, G.K.; De Souza, D.F.; Peralta, R.M. Production of laccase isoforms by Pleurotus pulmonarius in response to presence of phenolic and aromatic compounds. J. Basic Microbiol. 2004, 44, 129–136. [Google Scholar] [CrossRef]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and transcriptional regulation of laccases in fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Kollmann, A.; Boyer, F.D.; Ducrot, P.H.; Kerhoas, L.; Jolivalt, C.; Touton, I.; Einhorn, J.; Mougin, C. Oligomeric compounds formed from 2,5-xylidine (2,5-dimethylaniline) are potent enhancers of laccase production in Trametes versicolor ATCC 32745. Appl. Microbiol. Biotechnol. 2005, 68, 251–258. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, dietary sources and bioavailability. Ann. Ist. Super. Sanita 2007, 43, 348–361. [Google Scholar]
- Itagaki, S.; Kurokawa, T.; Nakata, C.; Saito, Y.; Oikawa, S.; Kobayashi, M.; Hirano, T.; Iseki, K. In vitro and in vivo antioxidant properties of ferulic acid: A comparative study with other natural oxidation inhibitors. Food Chem. 2009, 114, 466–471. [Google Scholar] [CrossRef]
- Barone, E.; Calabrese, V.; Mancuso, C. Ferulic acid and its therapeutic potential as a hormetin for age-related diseases. Biogerontology 2009, 10, 97–108. [Google Scholar] [CrossRef]
- Maurya, D.K.; Devasagayam, T.P.A. Antioxidant and prooxidant nature of hydroxycinnamic acid derivatives ferulic and caffeic acids. Food Chem. Toxicol. 2010, 48, 3369–3373. [Google Scholar] [CrossRef]
- Mancuso, C.; Santangelo, R. Ferulic acid: Pharmacological and toxicological aspects. Food Chem. Toxicol. 2014, 65, 185–195. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Amici, M.; Cuccioloni, M.; Angeletti, M.; Keller, J.N.; Eleuteri, A.M. Natural polyphenols as proteasome modulators and their role as anti-cancer compounds. FEBS J. 2008, 275, 5512–5526. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Atanasov, A.G.; Khan, H.; Barreca, D.; Trombetta, D.; Testai, L.; Sureda, A.; Tejada, S.; Vacca, R.A.; Pittalà, V.; et al. Targeting ubiquitin-proteasome pathway by natural, in particular polyphenols, anticancer agents: Lessons learned from clinical trials. Cancer Lett. 2018, 434, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, K.J.; Dean, R.T. Assessment of proteasome activity in cell lysates and tissue homogenates using peptide substrates. Int. J. Biochem. Cell Biol. 2003, 35, 716–727. [Google Scholar] [CrossRef]
- Kim, H.T.; Collins, G.A.; Goldberg, A.L. Measurement of the multiple activities of 26S proteasomes. Methods Mol. Biol. 2018, 1844, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. Tracing the history of the ubiquitin proteolytic system: The pioneering article. Biochem. Biophys. Res. Commun. 2009, 387, 1–10. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Goldberg, A.L. Proteasome inhibitors: From research tools to drug candidates. Chem. Biol. 2001, 8, 739–758. [Google Scholar] [CrossRef] [Green Version]
- Milacic, V.; Banerjee, S.; Landis-Piwowar, K.R.; Sarkar, F.H.; Majumdar, A.P.N.; Dou, Q.P. Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res. 2008, 68, 7283–7292. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.E.; Rattan, S.I.S. Curcumin’s biphasic hormetic response on proteasome activity and heat-shock protein synthesis in human keratinocytes. Ann. N. Y. Acad. Sci. 2006, 1067, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Cascio, P.; Goldberg, A.L. Preparation of hybrid (19S-20S-PA28) proteasome complexes and analysis of peptides generated during protein degradation. Methods Enzymol. 2005, 398, 336–352. [Google Scholar] [CrossRef]
- Smith, D.M.; Kafri, G.; Cheng, Y.; Ng, D.; Walz, T.; Goldberg, A.L. ATP binding to PAN or the 26S ATPases causes association with the 20S proteasome, gate opening, and translocation of unfolded proteins. Mol. Cell 2005, 20, 687–698. [Google Scholar] [CrossRef]
- Staszczak, M.; Zdunek, E.; Leonowicz, A. Studies on the role of proteases in the white-rot fungus Trametes versicolor: Effect of PMSF and chloroquine on ligninolytic enzymes activity. J. Basic Microbiol. 2000, 40, 51–63. [Google Scholar] [CrossRef]
- Fåhraeus, G.; Reinhammar, B. Large scale production and purification of laccase from cultures of the fungus Polyporus versicolor and some properties of laccase A. Acta Chem. Scand. 1967, 21, 2367–2378. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Yoshimura, T.; Ichihara, A. Role of substrate in reversible activation of proteasomes (multi-protease complexes) by sodium dodecyl sulfate. J. Biochem. 1989, 106, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Skoda, B.; Malek, L. Dry pea seed proteasome: Purification and enzymic activities. Plant Physiol. 1992, 99, 1515–1519. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.; Stober, F.; Lichtenthaler, H.K. Fluorescence emission spectra of plant leaves and plant constituents. Radiat. Environ. Biophys. 1991, 30, 333–347. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Wong, C.; Sridhara, S.; Bardwell, J.C.A.; Jakob, U. Heating greatly speeds Coomassie blue staining and destaining. Biotechniques 2000, 28, 426–432. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Final Concentration (mM) | Remaining CHTL Activity 1 (% of Control) |
|---|---|---|
| None | - | 100 |
| Ferulic acid | 0.002 | 101 |
| 0.02 | 99 | |
| 0.2 | 85 | |
| 1 | 68 | |
| p-Hydroxybenzoic acid | 0.002 | 101 |
| 0.02 | 98 | |
| 0.2 | 94 | |
| 1 | 100 | |
| Protocatechuic acid | 0.002 | 105 |
| 0.02 | 98 | |
| 0.2 | 92 | |
| 1 | 103 | |
| Syringic acid | 0.002 | 102 |
| 0.02 | 104 | |
| 0.2 | 105 | |
| 1 | 102 | |
| Vanillic acid | 0.002 | 107 |
| 0.02 | 107 | |
| 0.2 | 96 | |
| 1 | 100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swatek, A.; Staszczak, M. Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro. Int. J. Mol. Sci. 2020, 21, 2463. https://doi.org/10.3390/ijms21072463
Swatek A, Staszczak M. Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro. International Journal of Molecular Sciences. 2020; 21(7):2463. https://doi.org/10.3390/ijms21072463
Chicago/Turabian StyleSwatek, Anita, and Magdalena Staszczak. 2020. "Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro" International Journal of Molecular Sciences 21, no. 7: 2463. https://doi.org/10.3390/ijms21072463
APA StyleSwatek, A., & Staszczak, M. (2020). Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro. International Journal of Molecular Sciences, 21(7), 2463. https://doi.org/10.3390/ijms21072463