Hydroxychloroquine Mitigates the Production of 8-Isoprostane and Improves Vascular Dysfunction: Implications for Treating Preeclampsia

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
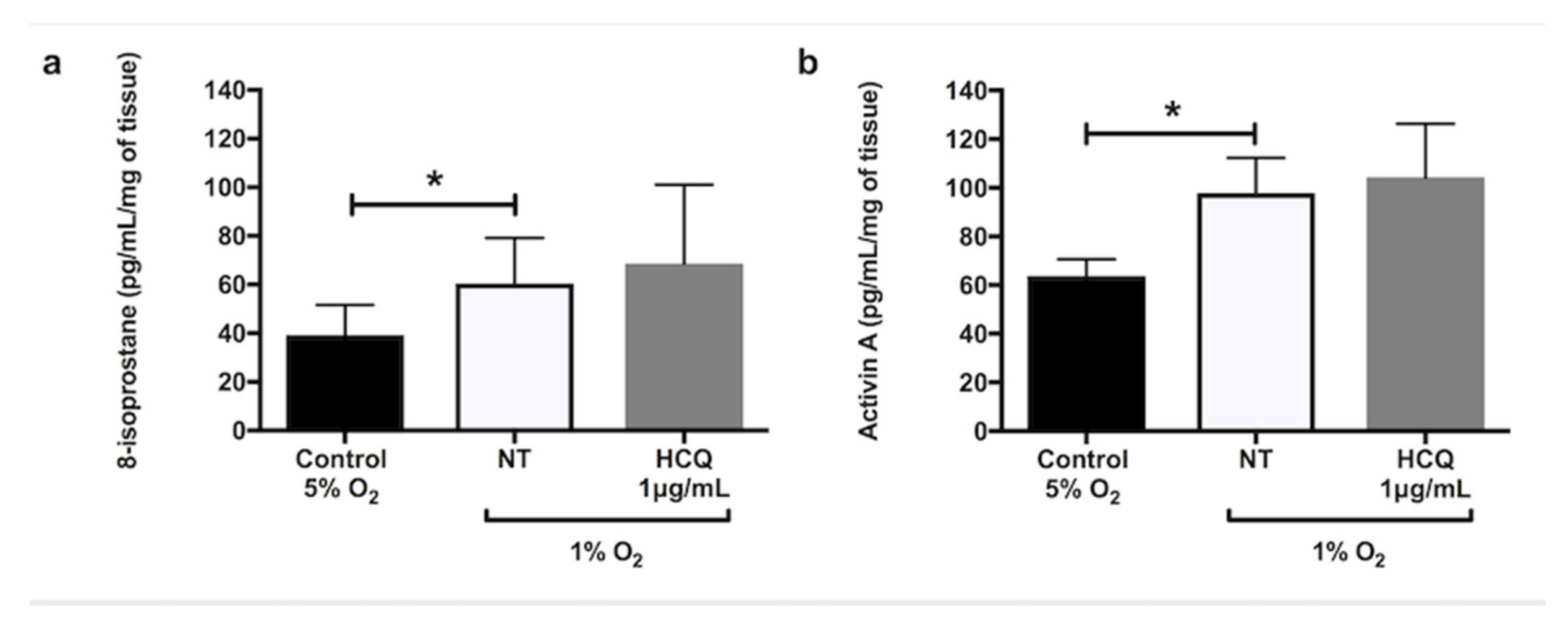
2.1. Effects of Hydroxychloroquine on Placental Secretion
2.2. Effect of Hydroxychloroquine on HUVEC Viability
2.3. Effects of Hydroxychloroquine on Endothelial Function In Vitro
2.4. Effect of Hydroxychloroquine on Vascular Permeability
2.5. Effect of Hydroxychloroquine on Zonula Occludens 1 (ZO-1) Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Blood and Tissue Collection
4.2. Placental Explant Cultures Ex Vivo
4.3. Placental Hypoxia
4.4. Placental Oxidative Stress
4.5. Measurement of sFlt-1, sEng, TNF-α, and Activin A with ELISA
4.6. Human Umbilical vein Endothelial Cell (HUVEC) Isolation
4.7. HUVEC Viability Assay
4.8. Oxidative Stress as Assessed by 8-Isoprostane
4.9. Measurement of NADPH Oxidase (NOX2) mRNA Expression
4.10. Measurement of NOX2 Protein Expression
4.11. Endothelial Permeability Assay
4.12. Zonula Occludens (ZO-1) Immunohistochemistry for the Assessment of Endothelial Integrity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sibai, B.; Dekker, G.; Kupfermine, M. Pre-eclampsia. Lancet 2005, 365, 785–799. [Google Scholar] [CrossRef]
- Mackay, A.; Berg, C.; Atrash, H. Pregnancy-related mortality from preeclampsia and eclampsia. Obstet. Gynecol. 2001, 97, 533–538. [Google Scholar] [PubMed]
- Kucukgoz Gulec, U.; Ozgunen, F.T.; Buyukkurt, S.; Guzel, A.B.; Urunsak, I.F.; Demir, S.C.; Evruke, I.C. Comparison of clinical and laboratory findings in early- and late-onset preeclampsia. J. Matern. Fetal Neonatal Med. 2013, 26, 1228–1233. [Google Scholar] [CrossRef] [PubMed]
- Lisonkova, S.; Sabr, Y.; Mayer, C.; Young, C.; Skoll, A.; Joseph, K.S. Maternal morbidity associated with early-onset and late-onset preeclampsia. Obstet. Gynecol. 2014, 124, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W. Preeclampsia: A multi-stress disorder. Rev. Med. Interne 2011, 32, S41–S44. [Google Scholar] [CrossRef]
- Nagamatsu, T.; Fujii, T.; Kusumi, M.; Zou, L.; Yamashita, T.; Osuga, Y.; Momoeda, M.; Kozuma, S.; Taketani, Y. Cytotrophoblasts up-regulate soluble fms-like tyrosine kinase-1 expression under reduced oxygen: An implication for the placental vascular development and the pathophysiology of preeclampsia. Endocrinology 2004, 145, 4838–4845. [Google Scholar] [CrossRef]
- Gilbert, J.S.; Gilbert, S.A.; Arany, M.; Granger, J.P. Hypertension produced by placental ischemia in pregnant rats is associated with increased soluble endoglin expression. Hypertension 2009, 53, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Schneider, H.; Aliyev, E.; Soydemir, F.; Baumann, M.; Surbek, D.; Hediger, M.; Brownbill, P.; Albrecht, C. Hypoxic treatment of human dual placental perfusion induces a preeclampsia-like inflammatory response. Lab Investig. 2014, 94, 873–880. [Google Scholar] [CrossRef]
- Mandang, S.; Manuelpillai, U.; Wallace, E.M. Oxidative stress increases placental and endothelial cell activin A secretion. J. Endocrinol. 2007, 192, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Tam Tam, K.B.; Lamarca, B.; Arany, M.; Cockrell, K.; Fournier, L.; Murphy, S.; Martin, J.N., Jr.; Granger, J.P. Role of reactive oxygen species during hypertension in response to chronic antiangiogenic factor (sFlt-1) excess in pregnant rats. Am. J. Hypertens. 2011, 24, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Onda, K.; Tong, S.; Nakahara, A.; Kondo, M.; Monchusho, H.; Hirano, T.; Kaitu’u-Lino, T.; Beard, S.; Binder, N.; Tuohey, L.; et al. Sofalcone upregulates the nuclear factor (erythroid-derived 2)-like 2/heme oxygenase-1 pathway, reduces soluble fms-like tyrosine kinase-1, and quenches endothelial dysfunction: Potential therapeutic for preeclampsia. Hypertension 2015, 65, 855–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownfoot, F.C.; Tong, S.; Hannan, N.J.; Hastie, R.; Cannon, P.; Tuohey, L.; Kaitu’u-Lino, T.J. YC-1 reduces placental sFlt-1 and soluble endoglin production and decreases endothelial dysfunction: A possible therapeutic for preeclampsia. Mol. Cell. Endocrinol. 2015, 413, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.; Acharya, R.; Delpachitra, P.; Hobson, S.; Sobey, C.G.; Drummond, G.R.; Wallace, E.M. Activin and NADPH-oxidase in preeclampsia: Insights from in vitro and murine studies. Am. J. Obstet. Gynecol. 2015, 212, e1–e12. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; Mires, G.; Macleod, M.; Baker, P. In preeclampsia, the circulating factors capable of altering in vitro endothelial function precede clinical disease. Hypertension 2005, 45, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gu, Y.; Zhang, Y.; Lewis, D.F. Evidence of endothelial dysfunction in preeclampsia: Decreased endothelial nitric oxide synthase expression is associated with increased cell permeability in endothelial cells from preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 817–824. [Google Scholar] [CrossRef]
- Wallace, D.J. The history of antimalarials. Lupus 1996, 5, S2–S3. [Google Scholar] [CrossRef]
- Wallace, D.J.; Gudsoorkar, V.S.; Weisman, M.H.; Venuturupalli, S.R. New insights into mechanisms of therapeutic effects of antimalarial agents in SLE. Nat. Rev. Rheumatol. 2012, 8, 522–533. [Google Scholar] [CrossRef]
- US Food and Drug Administration Pregnancy Category. Available online: http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfCFR/CFRSearch.cfm?fr=201.57 (accessed on 6 August 2015).
- Costedoat-Chalumeau, N.; Amoura, Z.; Duhaut, P.; Huong, D.L.; Sebbough, D.; Wechsler, B.; Vauthier, D.; Denjoy, I.; Lupoglazoff, J.M.; Piette, J.C. Safety of hydroxychloroquine in pregnant patients with connective tissue diseases: A study of one hundred thirty-three cases compared with a control group. Arthritis Rheum. 2003, 48, 3207–3211. [Google Scholar] [CrossRef]
- Miyachi, Y.; Yoshioka, A.; Imamura, S.; Niwa, Y. Antioxidant action of antimalarials. Ann. Rheum. Dis. 1986, 45, 244–248. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ertel, W.; Ayala, A.; Morrison, M.; Perrin, M.; Chaudry, I. Chloroquine inhibits macrophage tumour necrosis factor-α mRNA transcription. Immunology 1993, 80, 122–126. [Google Scholar]
- Karres, I.; Kremer, J.; Dietl, I.; Steckholzer, U.; Jochum, M.; Ertel, W. Chloroquine inhibits proinflammatory cytokine release into human whole blood. Am. J. Physiol. 1998, 43, R1058–R1064. [Google Scholar] [CrossRef] [PubMed]
- Willis, R.; Seif, A.M.; McGwin, G., Jr.; Martinez-Martinez, L.A.; Gonzalez, E.B.; Dang, N.; Papalardo, E.; Liu, J.; Vila, L.M.; Reveille, J.D.; et al. Effect of hydroxychloroquine treatment on pro-inflammatory cytokines and disease activity in SLE patients: Data from LUMINA (LXXV), a multiethnic US cohort. Lupus 2012, 21, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Mariz, H.A.; Rocha Jr, L.F.; Oliveira, P.S.; Dantas, A.T.; Duarte, A.L.; Pitta, I.R.; Galdino, S.L.; Pitta, M.G. Hydroxychloroquine decreases Th17-related cytokines in systemic lupus erythematosus and rheumatoid arthritis patients. Clinics 2013, 68, 766–771. [Google Scholar] [CrossRef]
- Gomez-Guzman, M.; Jimenez, R.; Romero, M.; Sanchez, M.; Zarzuelo, M.J.; Gomez-Morales, M.; O’Valle, F.; Lopez-Farre, A.J.; Algieri, F.; Galvez, J.; et al. Chronic hydroxychloroquine improves endothelial dysfunction and protects kidney in a mouse model of systemic lupus erythematosus. Hypertension 2014, 64, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Rahman, R.; Murthi, P.; Singh, H.; Gurusinghe, S.; Mockler, J.C.; Lim, R.; Wallace, E.M. The effects of hydroxychloroquine on endothelial dysfunction. Pregnancy Hypertens. Int. J. Women’s Cardiovasc. Health 2016, 6, 259–262. [Google Scholar] [CrossRef]
- Albert, C.R.; Schlesinger, W.J.; Viall, C.A.; Mulla, M.J.; Brosens, J.J.; Chamley, L.W.; Abrahams, V.M. Effect of hydroxychloroquine on antiphospholipid antibody-induced changes in first trimester trophoblast function. Am. J. Reprod. Immunol. 2014, 71, 154–164. [Google Scholar] [CrossRef]
- Roberts, J.; Taylor, R.; Musci, T.; Rodgers, G.; Hubel, C.; McLaughlin, M. Preeclampsia-an endothelial disorder. Am. J. Obstet. Gynecol. 1989, 161, 1200–1204. [Google Scholar] [CrossRef]
- LaMarca, B. Endothelial dysfunction; an important mediator in the pathophysiology of hypertension during preeclampsia. Minerva Ginecol. 2012, 64, 309–320. [Google Scholar]
- Bilodeau, J.F.; Qin Wei, S.; Larose, J.; Greffard, K.; Moisan, V.; Audibert, F.; Fraser, W.D.; Julien, P. Plasma F2-isoprostane class VI isomers at 12-18 weeks of pregnancy are associated with later occurrence of preeclampsia. Free Radic. Biol. Med. 2015, 85, 282–287. [Google Scholar] [CrossRef] [Green Version]
- George, E.M.; Hosick, P.A.; Stec, D.E.; Granger, J.P. Heme oxygenase inhibition increases blood pressure in pregnant rats. Am. J. Hypertens. 2013, 26, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Miesel, R.; Hartung, R.; Kroeger, H. Priming Of NADPH oxidase by tumor necrosis factor-α in patients with inflammatory and autoimmune rheumatic diseases. Inflammation 1996, 20, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Virdis, A.; Tani, C.; Duranti, E.; Vagnani, S.; Carli, L.; Kuhl, A.A.; Solini, A.; Baldini, C.; Talarico, R.; Bombardieri, S.; et al. Early treatment with hydroxychloroquine prevents the development of endothelial dysfunction in a murine model of systemic lupus erythematosus. Arthritis Res. Ther. 2015, 17, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller-Calleja, N.; Manukyan, D.; Canisius, A.; Strand, D.; Lackner, K.J. Hydroxychloroquine inhibits proinflammatory signalling pathways by targeting endosomal NADPH oxidase. Ann. Rheum. Dis. 2017, 76, 891–897. [Google Scholar] [CrossRef]
- Dan, I.; Watanabe, M.N.; Kusumi, A. The Ste20 group kinases as regulators of MAP kinase cascades. Trends Cell Biol. 2001, 11, 220–230. [Google Scholar] [CrossRef]
- Hobson, S.R.; Acharya, R.; Lim, R.; Chan, S.T.; Mockler, J.; Wallace, E.M. Role of activin A in the pathogenesis of endothelial cell dysfunction in preeclampsia. Pregnancy Hypertens. 2016, 6, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Muttukrishna, S.; Knight, P.G.; Groome, N.P.; Redman, C.W.G.; Ledger, W.L. Activin A and inhibin A as possible endocrine markers for pre-eclampsia. Lancet 1997, 349, 1285–1288. [Google Scholar] [CrossRef]
- Kakei, Y.; Akashi, M.; Shigeta, T.; Hasegawa, T.; Komori, T. Alteration of cell-cell junctions in cultured human lymphatic endothelial cells with inflammatory cytokine stimulation. Lymphat. Res. Biol. 2014, 12, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Aveleira, C.; Lin, C.; Abcouwer, S.; Ambrosio, A.; Antonetti, D. TNF-α signals through PKC/NFkB to alter the tight junction complex and increase retinal endothelial cell permeability. Diabetes 2010, 59, 2872–2882. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, Z.; Bayraktutan, U. NADPH oxidase mediates TNF-α-evoked in vitro brain barrier dysfunction: Roles of apoptosis and time. Mol. Cell. Neurosci. 2014, 61, 72–84. [Google Scholar] [CrossRef]
- Vince, S.G.; Starkey, M.P.; Austgulen, R.; Kowiatkowski, D.; Redman, C.W.G. Interleukin-6, turnour necrosis factor and soluble tumour necrosis factor receptors in women with pre-eclampsia. Br. J. Obstet. Gynaecol. 1995, 102, 20–25. [Google Scholar] [CrossRef]
- Conrad, K.P.; Miles, T.M.; Benyo, D.F. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am. J. Reprod. Immunol. 1998, 40, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Teran, E.; Escudero, C.; Moya, W.; Flores, M.; Vallance, P.; Lopez-Jaramillo, P. Elevated C-reactive protein and pro-inflammatory cytokines in Andean women with pre-eclampsia. Int. J. Gynaecol. Obstet. 2001, 75, 243–249. [Google Scholar] [CrossRef]
- Tannetta, D.S.; Muttukrishna, S.; Groome, N.P.; Redman, C.W.; Sargent, I.L. Endothelial cells and peripheral blood mononuclear cells are a potential source of extraplacental activin a in preeclampsia. J. Clin. Endocrinol. Metab. 2003, 88, 5995–6001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznik, A.; Bencina, M.; Svajger, U.; Jeras, M.; Rozman, B.; Jerala, R. Mechanism of endosomal TLR inhibition by antimalarial drugs and imidazoquinolines. J. Immunol. 2011, 186, 4794–4804. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, P.; Weaver, L.E.; Doersch, K.M.; Kopriva, S.E.; Chiasson, V.L.; Allen, S.J.; Narayanan, A.M.; Young, K.J.; Jones, K.A.; Kuehl, T.J.; et al. Placental Toll-like receptor 3 and Toll-like receptor 7/8 activation contributes to preeclampsia in humans and mice. PLoS ONE 2012, 7, e41884. [Google Scholar] [CrossRef]
- Tinsley, J.H.; Chiasson, V.L.; Mahajan, A.; Young, K.J.; Mitchell, B.M. Toll-like receptor 3 activation during pregnancy elicits preeclampsia-like symptoms in rats. Am. J. Hypertens. 2009, 22, 1314–1319. [Google Scholar] [CrossRef] [Green Version]
- Pulkkinen, M.; Kivikoski, A.; Nevalainen, T. Group 1 and group II phospholipase A2 in serum during normal and pathological pregnancy. Gynecol. Obstet. Investig. 1993, 36, 96–101. [Google Scholar] [CrossRef]
- Staff, A.; Ranheim, T.; Halvorsen, B. Augmented PLA2 activity in pre-eclamptic decidual tissue—A key player in the pathophysiology of ‘acute atherosis’ in pre-eclampsia? Placenta 2003, 24, 965–973. [Google Scholar] [CrossRef]
- Pruzanski, W.; Goulding, N.; Flower, R.; Gladman, D.; Urowitz, M.; Goodman, P.; Scott, K.; Vadas, P. Circulating group II phospholipase A2 activity and antilipocortin antibodies in systemic lupus erythematosus. Correlative study with disease activity. J. Rheumatol. 1994, 21, 252–257. [Google Scholar]
- Au, A.; Chan, P.; Fishman, R. Stimulation of phospholipase A2 activity by oxygen-derived free radicals in isolated brain capillaries. J. Cell. Biochem. 1985, 27, 449–453. [Google Scholar] [CrossRef]
- Zabul, P.; Wozniak, M.; Slominski, A.T.; Preis, K.; Gorska, M.; Korozan, M.; Wieruszewski, J.; Zmijewski, M.A.; Zabul, E.; Tuckey, R.; et al. A proposed molecular mechanism of high-dose vitamin D3 supplementation in prevention and treatment of preeclampsia. Int. J. Mol. Sci. 2015, 16, 13043–13064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maridonneau-Parini, I.; Tauber, A. Activation of NADPH-oxidase by arachidonic acid involves phospholipase A2 in intact human neutrophils but not in the cell-free system. Biochem. Biophys. Res. Commun. 1986, 138, 1099–1105. [Google Scholar] [CrossRef]
- Henderson, L.; Chappell, J.; Jones, O. Superoxide generation is inhibited by phospholipase A2 inhibitors. Biochem. J. 1989, 264, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, S.; Bowyer, L.; Lust, K.; McMahon, L.; Morton, M.; North, R.; Paech, M.; Said, J. Guidelines for the management of hypertensive disorders of pregnancy 2014. Aust. N. Z. J. Obstet. Gynaecol. 2014, 49, 242–246. [Google Scholar] [CrossRef]
- Murata, M.; Fukushima, K.; Seki, H.; Takeda, S.; Wake, N. Oxidative stress produced by xanthine oxidase induces apoptosis in human extravillous trophoblast cells. J. Reprod. Dev. 2013, 59, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Montuschi, P.; Barnes, P.J.; Roberts, L.J., 2nd. Isoprostanes: Markers and mediators of oxidative stress. FASEB J. 2004, 18, 1791–1800. [Google Scholar] [CrossRef]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, R.A.; Murthi, P.; Singh, H.; Gurungsinghe, S.; Leaw, B.; Mockler, J.C.; Lim, R.; Wallace, E.M. Hydroxychloroquine Mitigates the Production of 8-Isoprostane and Improves Vascular Dysfunction: Implications for Treating Preeclampsia. Int. J. Mol. Sci. 2020, 21, 2504. https://doi.org/10.3390/ijms21072504
Rahman RA, Murthi P, Singh H, Gurungsinghe S, Leaw B, Mockler JC, Lim R, Wallace EM. Hydroxychloroquine Mitigates the Production of 8-Isoprostane and Improves Vascular Dysfunction: Implications for Treating Preeclampsia. International Journal of Molecular Sciences. 2020; 21(7):2504. https://doi.org/10.3390/ijms21072504
Chicago/Turabian StyleRahman, Rahana Abd, Padma Murthi, Harmeet Singh, Seshini Gurungsinghe, Bryan Leaw, Joanne C. Mockler, Rebecca Lim, and Euan M. Wallace. 2020. "Hydroxychloroquine Mitigates the Production of 8-Isoprostane and Improves Vascular Dysfunction: Implications for Treating Preeclampsia" International Journal of Molecular Sciences 21, no. 7: 2504. https://doi.org/10.3390/ijms21072504
APA StyleRahman, R. A., Murthi, P., Singh, H., Gurungsinghe, S., Leaw, B., Mockler, J. C., Lim, R., & Wallace, E. M. (2020). Hydroxychloroquine Mitigates the Production of 8-Isoprostane and Improves Vascular Dysfunction: Implications for Treating Preeclampsia. International Journal of Molecular Sciences, 21(7), 2504. https://doi.org/10.3390/ijms21072504