Molecular Dynamics Simulation and Kinetic Study of Fluoride Binding to V21C/V66C Myoglobin with a Cytoglobin-like Disulfide Bond


Abstract
:1. Introduction
2. Results and Discussion
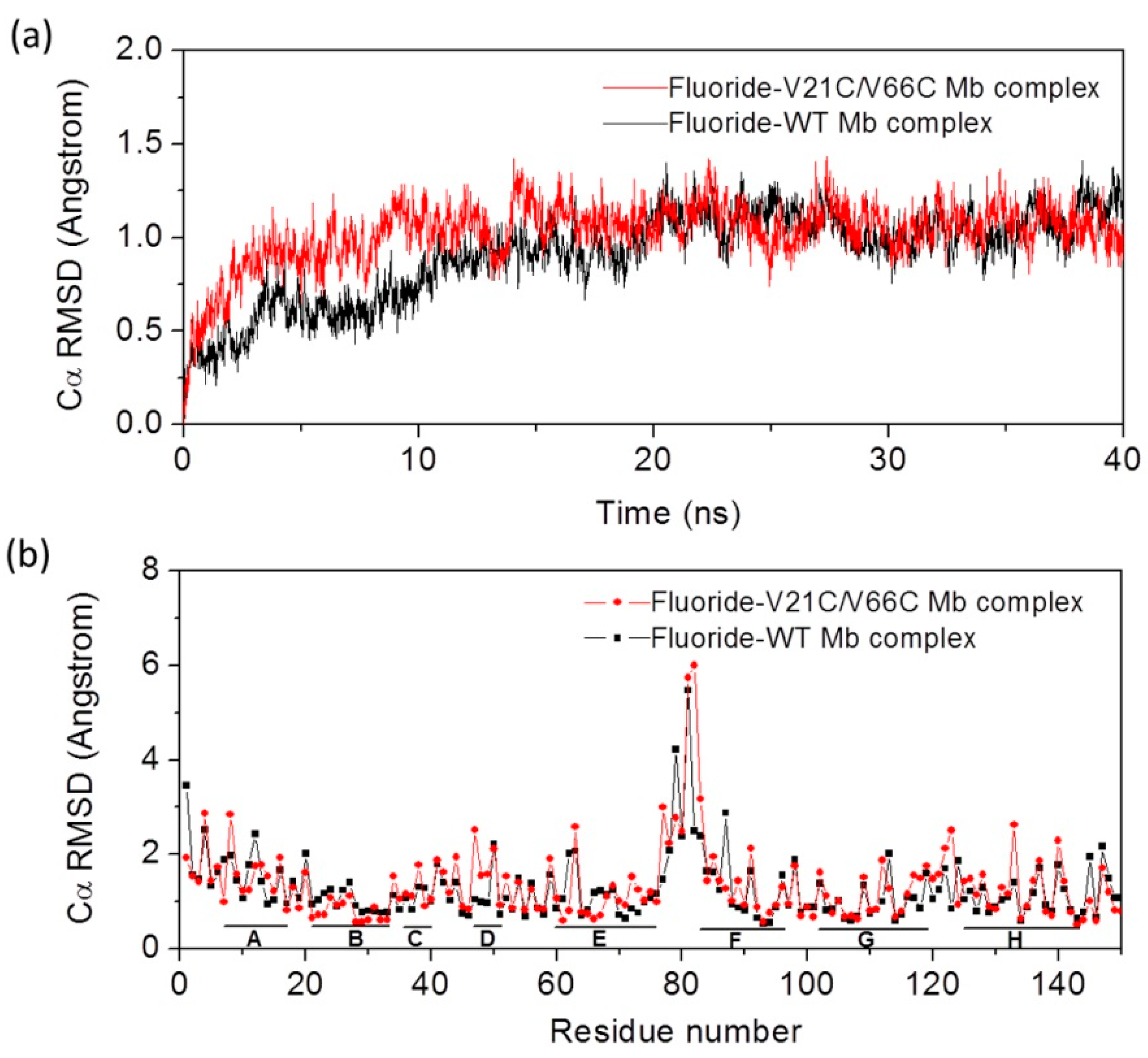
2.1. Protein Motions of Fluoride-Mb Complexes
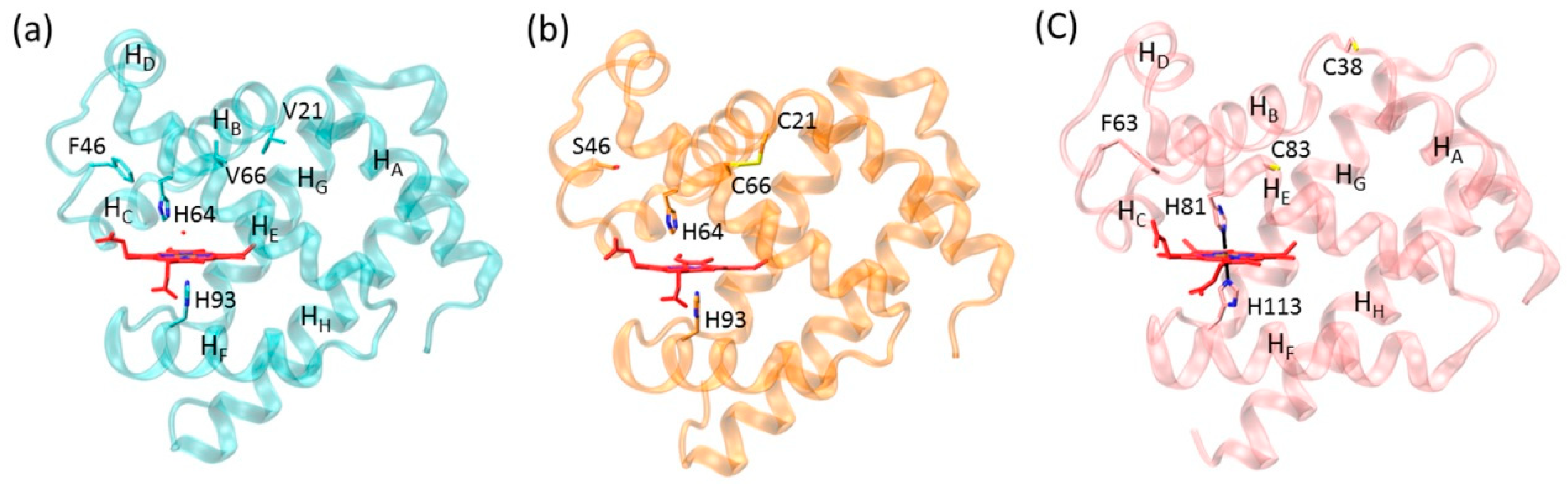
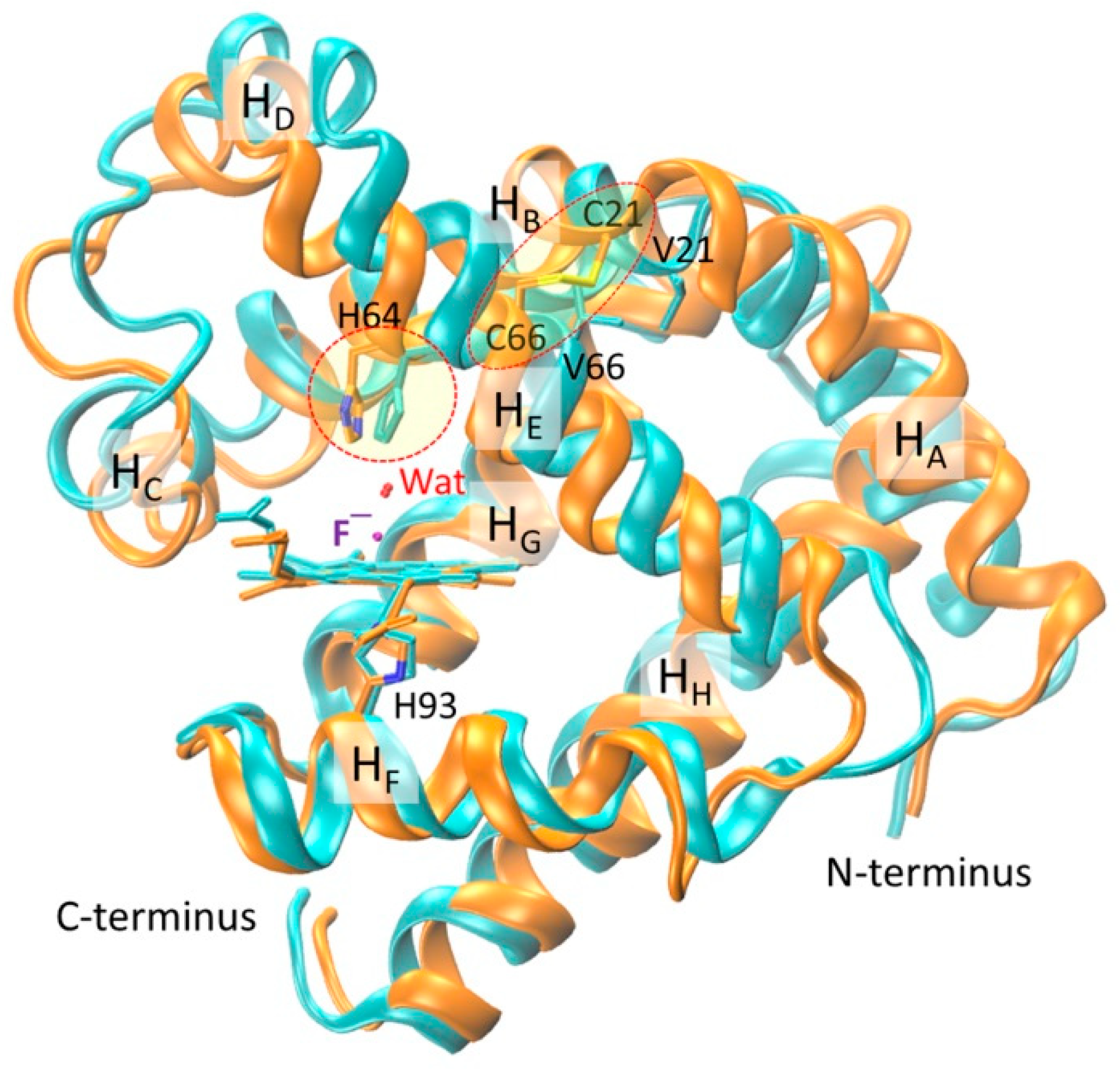
2.2. Structural Comparison of Fluoride–Mb Complexes
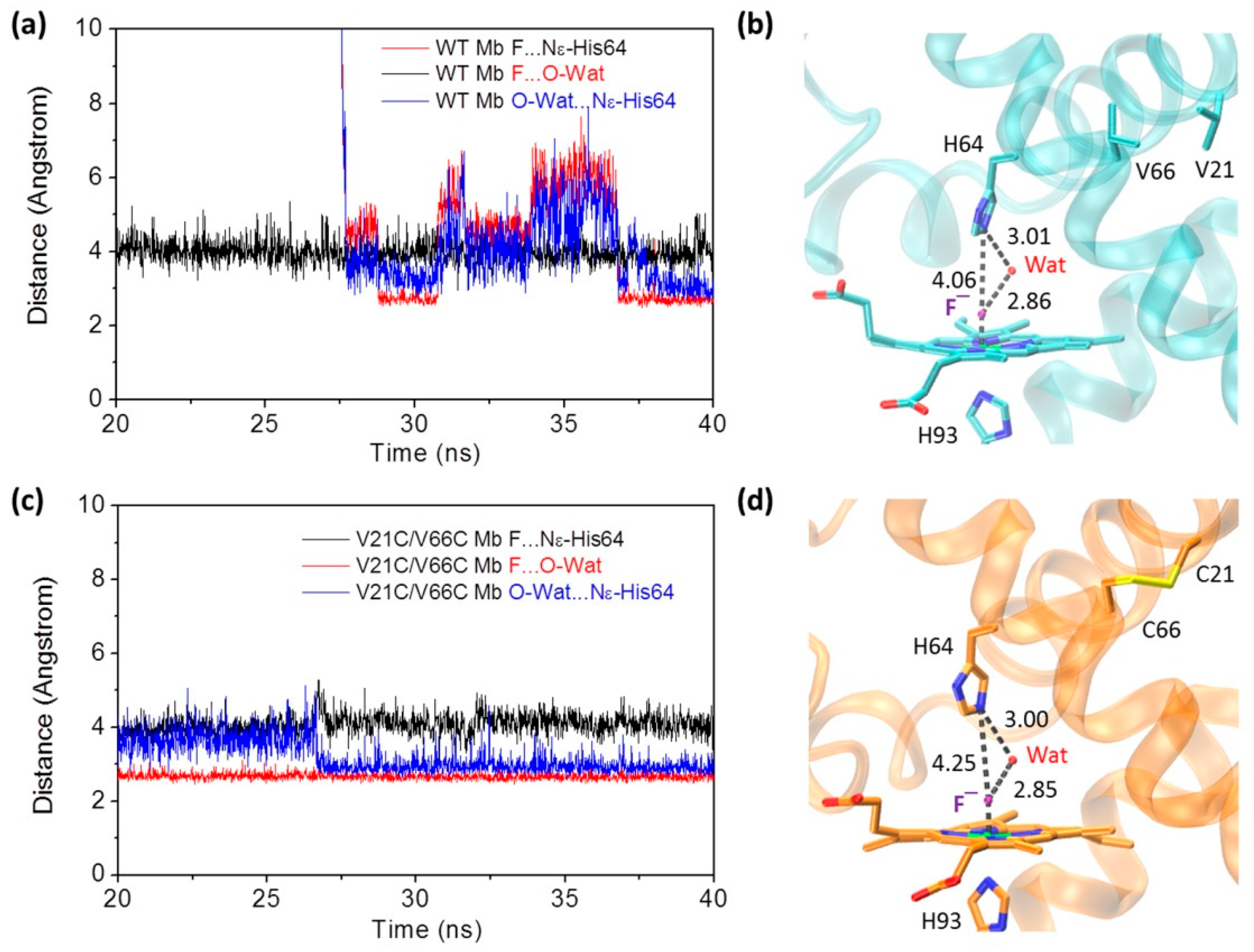
2.3. Interactions of Fluoride ion with Mbs
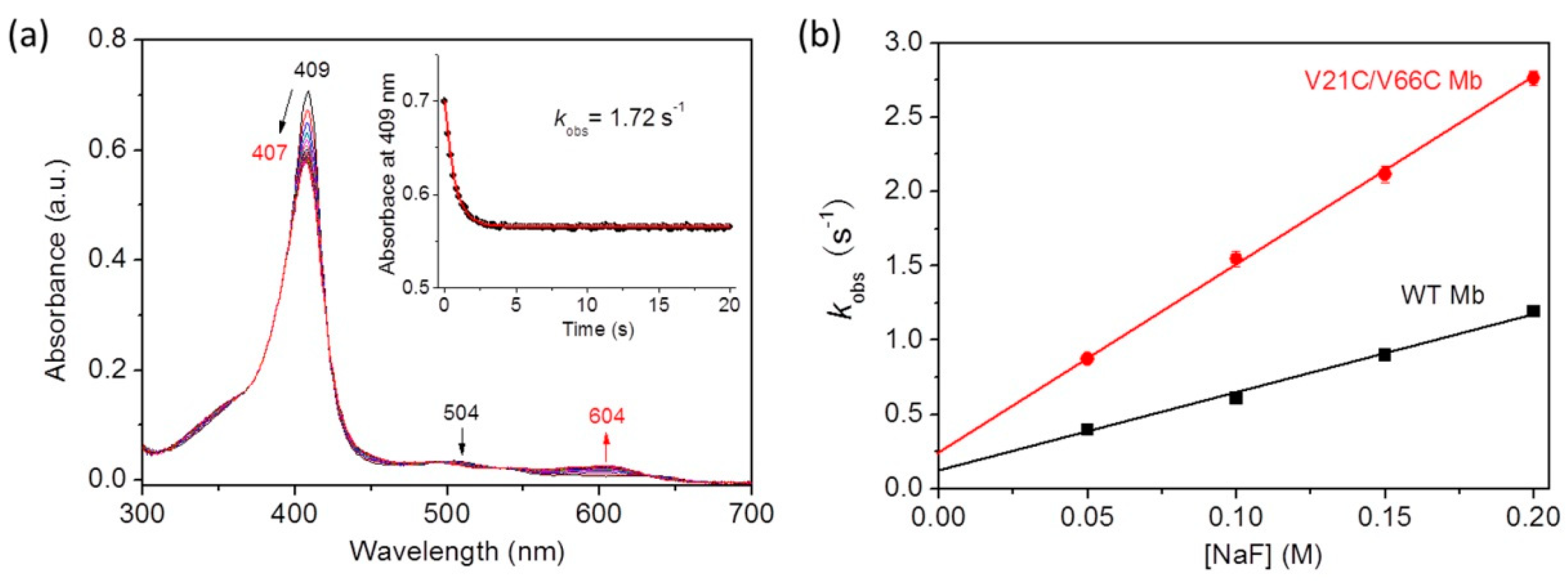
2.4. Kinetic Binding of Fluoride ion to Mbs
3. Conclusions
4. Materials and Methods
4.1. Materials and Reagents
4.2. Molecular Dynamics Studies
4.3. UV–Vis Kinetic Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lu, Y.; Yeung, N.; Sieracki, N.; Marshall, N.M. Design of functional metalloproteins. Nature 2009, 460, 855–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-W. Rational design of metalloenzymes: From single to multiple active sites. Coord. Chem. Rev. 2017, 336, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Hirota, S.; Lin, Y.-W. Design of artificial metalloproteins/metalloenzymes by tuning noncovalent interactions. J. Biol. Inorg. Chem. 2018, 23, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Schwizer, F.; Okamoto, Y.; Heinisch, T.; Gu, Y.; Pellizzoni, M.M.; Lebrun, V.; Reuter, R.; Kohler, V.; Lewis, J.C.; Ward, T.R. Artificial Metalloenzymes: Reaction Scope and Optimization Strategies. Chem. Rev. 2018, 118, 142–231. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hu, C.; Xia, L.; Wang, J. Artificial Metalloenzyme Design with Unnatural Amino Acids and Non-Native Cofactors. ACS Catal. 2018, 8, 1851–1863. [Google Scholar] [CrossRef]
- Mirts, E.N.; Bhagi-Damodaran, A.; Lu, Y. Understanding and Modulating Metalloenzymes with Unnatural Amino Acids, Non-Native Metal Ions, and Non-Native Metallocofactors. Acc. Chem. Res. 2019, 52, 935–944. [Google Scholar] [CrossRef]
- Oohora, K.; Onoda, A.; Hayashi, T. Hemoproteins Reconstituted with Artificial Metal Complexes as Biohybrid Catalysts. Acc. Chem. Res. 2019, 52, 945–954. [Google Scholar] [CrossRef]
- Shoji, O.; Aiba, Y.; Watanabe, Y. Hoodwinking Cytochrome P450BM3 into Hydroxylating Non-Native Substrates by Exploiting Its Substrate Misrecognition. Acc. Chem. Res. 2019, 52, 925–934. [Google Scholar] [CrossRef]
- Nastri, F.; D’Alonzo, D.; Leone, L.; Zambrano, G.; Pavone, V.; Lombardi, A. Engineering Metalloprotein Functions in Designed and Native Scaffolds. Trends Biochem. Sci. 2019, 44, 1022–1040. [Google Scholar] [CrossRef]
- Xu, J.; Wang, C.; Cong, Z. Strategies for Substrate-Regulated P450 Catalysis: From Substrate Engineering to Co-Catalysis. Chemistry 2019, 25, 6853–6863. [Google Scholar] [CrossRef]
- Lin, Y.-W. Rational Design of Artificial Metalloproteins and Metalloenzymes with Metal Clusters. Molecules 2019, 24, 2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-W. Rational design of heme enzymes for biodegradation of pollutants toward a green future. Biotechnol. Appl. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nanda, V.; Koder, R.L. Designing artificial enzymes by intuition and computation. Nat. Chem. 2010, 2, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-W. Structural insights into a low-spin myoglobin variant with bis-histidine coordination from molecular modeling. Proteins 2011, 79, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-W.; Nie, C.M.; Liao, L.F. Rational design of a nitrite reductase based on myoglobin: A molecular modeling and dynamics simulation study. J. Mol. Model. 2012, 18, 4409–4415. [Google Scholar] [CrossRef]
- Lin, Y.-W.; Shu, X.-G.; Du, K.-J.; Nie, C.-M.; Wen, G.-B. Computational insight into nitration of human myoglobin. Comput. Biol. Chem. 2014, 52, 60–65. [Google Scholar] [CrossRef]
- Muñoz Robles, V.; Ortega-Carrasco, E.; Alonso-Cotchico, L.; Rodriguez-Guerra, J.; Lledós, A.; Maréchal, J.-D. Toward the Computational Design of Artificial Metalloenzymes: From Protein–Ligand Docking to Multiscale Approaches. ACS Catal. 2015, 5, 2469–2480. [Google Scholar] [CrossRef]
- Moore, E.J.; Zorine, D.; Hansen, W.A.; Khare, S.D.; Fasan, R. Enzyme stabilization via computationally guided protein stapling. Proc. Natl. Acad. Sci. USA 2017, 114, 12472–12477. [Google Scholar] [CrossRef] [Green Version]
- Yeung, N.; Lin, Y.-W.; Gao, Y.-G.; Zhao, X.; Russell, B.S.; Lei, L.; Miner, K.D.; Robinson, H.; Lu, Y. Rational design of a structural and functional nitric oxide reductase. Nature 2009, 462, 1079–1082. [Google Scholar] [CrossRef] [Green Version]
- Mirts, E.N.; Petrik, I.D.; Hosseinzadeh, P.; Nilges, M.J.; Lu, Y. A designed heme-[4Fe-4S] metalloenzyme catalyzes sulfite reduction like the native enzyme. Science 2018, 361, 1098–1101. [Google Scholar] [CrossRef]
- Chalker, J.M.; Bernardes, G.J.; Lin, Y.A.; Davis, B.G. Chemical modification of proteins at cysteine: Opportunities in chemistry and biology. Chem. Asian J. 2009, 4, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-W. Structure and function of heme proteins regulated by diverse post-translational modifications. Arch. Biochem. Biophys. 2018, 641, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Reading, N.S.; Aust, S.D. Engineering a disulfide bond in recombinant manganese peroxidase results in increased thermostability. Biotechnol. Prog. 2000, 16, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Reading, N.S.; Aust, S.D. Role of disulfide bonds in the stability of recombinant manganese peroxidase. Biochemistry 2001, 40, 8161–8168. [Google Scholar] [CrossRef]
- Tiso, M.; Tejero, J.; Basu, S.; Azarov, I.; Wang, X.; Simplaceanu, V.; Frizzell, S.; Jayaraman, T.; Geary, L.; Shapiro, C.; et al. Human neuroglobin functions as a redox-regulated nitrite reductase. J. Biol. Chem. 2011, 286, 18277–18289. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, B.G.; Hamdane, D.; Lechauve, C.; Marden, M.C.; Golinelli-Pimpaneau, B. The crystal structure of wild-type human brain neuroglobin reveals flexibility of the disulfide bond that regulates oxygen affinity. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1005–1014. [Google Scholar] [CrossRef]
- Sugimoto, H.; Makino, M.; Sawai, H.; Kawada, N.; Yoshizato, K.; Shiro, Y. Structural basis of human cytoglobin for ligand binding. J. Mol. Biol. 2004, 339, 873–885. [Google Scholar] [CrossRef]
- Astudillo, L.; Bernad, S.; Derrien, V.; Sebban, P.; Miksovska, J. Reduction of the internal disulfide bond between Cys 38 and 83 switches the ligand migration pathway in cytoglobin. J. Inorg. Biochem. 2013, 129, 23–29. [Google Scholar] [CrossRef]
- Beckerson, P.; Reeder, B.J.; Wilson, M.T. Coupling of disulfide bond and distal histidine dissociation in human ferrous cytoglobin regulates ligand binding. FEBS Lett. 2015, 589, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Reeder, B.J.; Ukeri, J. Strong modulation of nitrite reductase activity of cytoglobin by disulfide bond oxidation: Implications for nitric oxide homeostasis. Nitric Oxide 2018, 72, 16–23. [Google Scholar] [CrossRef]
- Urayama, P.; Phillips Jr, G.N.; Gruner, S.M. Probing substates in sperm whale myoglobin using high-pressure crystallography. Structure 2002, 10, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.-B.; Yuan, H.; Zhou, H.; Gao, S.-Q.; Nie, C.-M.; Tan, X.; Wen, G.-B.; Lin, Y.-W. An intramolecular disulfide bond designed in myoglobin fine-tunes both protein structure and peroxidase activity. Arch. Biochem. Biophys. 2016, 600, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.-L.; Yuan, H.; Du, K.-J.; He, B.; Gao, S.-Q.; Wen, G.-B.; Tan, X.; Lin, Y.-W. Regulation of both the structure and function by a de novo designed disulfide bond: A case study of heme proteins in myoglobin. Chem. Commun. (Camb.) 2018, 54, 4356–4359. [Google Scholar] [CrossRef] [PubMed]
- Aime, S.; Fasano, M.; Paoletti, S.; Cutruzzola, F.; Desideri, A.; Bolognesi, M.; Rizzi, M.; Ascenzi, P. Structural determinants of fluoride and formate binding to hemoglobin and myoglobin: Crystallographic and 1H-NMR relaxometric study. Biophys. J. 1996, 70, 482–488. [Google Scholar] [CrossRef] [Green Version]
- Neri, F.; Kok, D.; Miller, M.A.; Smulevich, G. Fluoride binding in hemoproteins: The importance of the distal cavity structure. Biochemistry 1997, 36, 8947–8953. [Google Scholar] [CrossRef]
- Merryweather, J.; Summers, F.; Vitello, L.B.; Erman, J.E. Metmyoglobin/fluoride: Effect of distal histidine protonation on the association and dissociation rate constants. Arch. Biochem. Biophys. 1998, 358, 359–368. [Google Scholar] [CrossRef]
- Nicoletti, F.P.; Droghetti, E.; Boechi, L.; Bonamore, A.; Sciamanna, N.; Estrin, D.A.; Feis, A.; Boffi, A.; Smulevich, G. Fluoride as a probe for H-bonding interactions in the active site of heme proteins: The case of Thermobifida fusca hemoglobin. J. Am. Chem. Soc. 2011, 133, 20970–20980. [Google Scholar] [CrossRef]
- Droghetti, E.; Nicoletti, F.P.; Bonamore, A.; Sciamanna, N.; Boffi, A.; Feis, A.; Smulevich, G. The optical spectra of fluoride complexes can effectively probe H-bonding interactions in the distal cavity of heme proteins. J. Inorg. Biochem. 2011, 105, 1338–1343. [Google Scholar] [CrossRef]
- Kosowicz, J.G.; Boon, E.M. Insights into the distal heme pocket of H-NOX using fluoride as a probe for H-bonding interactions. J. Inorg. Biochem. 2013, 126, 91–95. [Google Scholar] [CrossRef]
- Bossa, C.; Amadei, A.; Daidone, I.; Anselmi, M.; Vallone, B.; Brunori, M.; Di Nola, A. Molecular dynamics simulation of sperm whale myoglobin: Effects of mutations and trapped CO on the structure and dynamics of cavities. Biophys. J. 2005, 89, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Du, J.-F.; Li, W.; Li, L.; Wen, G.-B.; Lin, Y.-W.; Tan, X. Regulating the coordination state of a heme protein by a designed distal hydrogen-bonding network. ChemistryOpen 2015, 4, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, F.P.; Thompson, M.K.; Howes, B.D.; Franzen, S.; Smulevich, G. New Insights into the Role of Distal Histidine Flexibility in Ligand Stabilization of Dehaloperoxidase−Hemoglobin from Amphitrite ornata. Biochemistry 2010, 49, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Hamdane, D.; Kiger, L.; Dewilde, S.; Green, B.N.; Pesce, A.; Uzan, J.; Burmester, T.; Hankeln, T.; Bolognesi, M.; Moens, L.; et al. Coupling of the heme and an internal disulfide bond in human neuroglobin. Micron 2004, 35, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Sturms, R.; DiSpirito, A.A.; Fulton, D.B.; Hargrove, M.S. Hydroxylamine Reduction to Ammonium by Plant and Cyanobacterial Hemoglobins. Biochemistry 2011, 50, 10829–10835. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Kalé, L.; Skeel, R.; Bhandarkar, M.; Brunner, R.; Gursoy, A.; Krawetz, N.; Phillips, J.; Shinozaki, A.; Varadarajan, K.; Schulten, K. NAMD2: Greater Scalability for Parallel Molecular Dynamics. J. Comput. Phys. 1999, 151, 283–312. [Google Scholar] [CrossRef]
- Springer, B.A.; Sligar, S.G. High-level expression of sperm whale myoglobin in Escherichia coli. Proc. Natl. Acad. Sci. USA 1987, 84, 8961–8965. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | kon (M−1 s −1) | koff (s −1) | kon/koff (M−1 ) |
|---|---|---|---|
| WT Mb | 5.14 ± 0.15 | 0.13 ± 0.01 | 39.5 |
| V21C/V66C Mb | 12.48 ± 0.26 | 0.27 ± 0.04 | 46.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, L.-L.; Xu, J.-K.; Wang, X.-J.; Gao, S.-Q.; Lin, Y.-W. Molecular Dynamics Simulation and Kinetic Study of Fluoride Binding to V21C/V66C Myoglobin with a Cytoglobin-like Disulfide Bond. Int. J. Mol. Sci. 2020, 21, 2512. https://doi.org/10.3390/ijms21072512
Yin L-L, Xu J-K, Wang X-J, Gao S-Q, Lin Y-W. Molecular Dynamics Simulation and Kinetic Study of Fluoride Binding to V21C/V66C Myoglobin with a Cytoglobin-like Disulfide Bond. International Journal of Molecular Sciences. 2020; 21(7):2512. https://doi.org/10.3390/ijms21072512
Chicago/Turabian StyleYin, Lu-Lu, Jia-Kun Xu, Xiao-Juan Wang, Shu-Qin Gao, and Ying-Wu Lin. 2020. "Molecular Dynamics Simulation and Kinetic Study of Fluoride Binding to V21C/V66C Myoglobin with a Cytoglobin-like Disulfide Bond" International Journal of Molecular Sciences 21, no. 7: 2512. https://doi.org/10.3390/ijms21072512
APA StyleYin, L. -L., Xu, J. -K., Wang, X. -J., Gao, S. -Q., & Lin, Y. -W. (2020). Molecular Dynamics Simulation and Kinetic Study of Fluoride Binding to V21C/V66C Myoglobin with a Cytoglobin-like Disulfide Bond. International Journal of Molecular Sciences, 21(7), 2512. https://doi.org/10.3390/ijms21072512