CD44 Can Compensate for IgSF11 Deficiency by Associating with the Scaffold Protein PSD-95 during Osteoclast Differentiation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Impaired Osteoclast Differentiation in IgSF11-Deficient Culture Is Rescued by Increased Cell Density
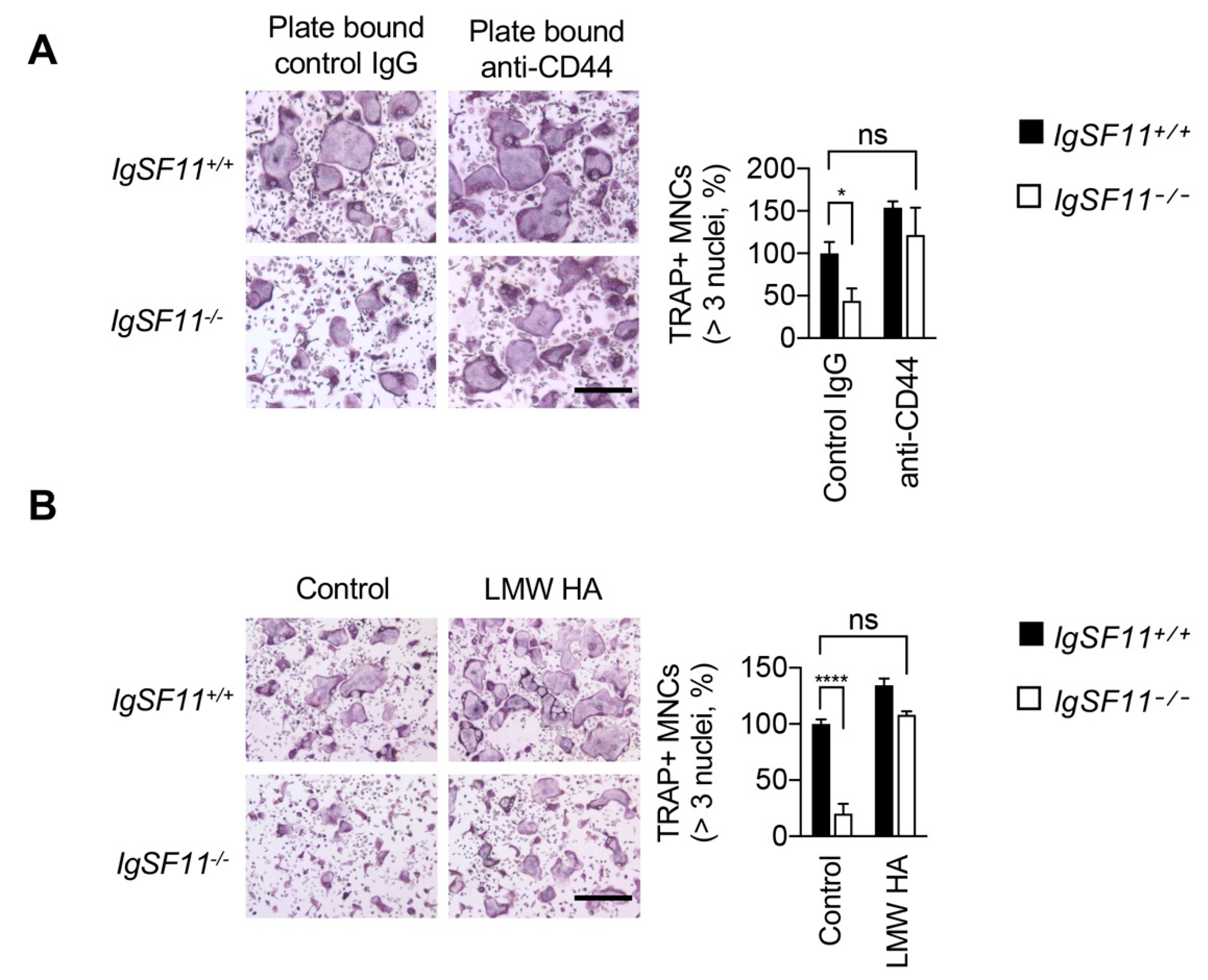
2.2. CD44 Stimulation Rescues Impaired IgSF11-Deficient Osteoclast Differentiation
2.3. PSD-95 Is Required for CD44-Mediated Osteoclast Differentiation
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. In Vitro Osteoclast Differentiation and Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.3. Co-Immunoprecipitation (Co-IP) and Western Blot
4.4. Retrovirus Preparation and Transduction
4.5. Reverse Transcription and Real-Time PCR (Q-PCR)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zaidi, M. Skeletal remodeling in health and disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Zelzer, E.; Olsen, B.R. The genetic basis for skeletal diseases. Nature 2003, 423, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Kim, N.; Kadono, Y.; Rho, J.; Lee, S.Y.; Lorenzo, J.; Choi, Y. Osteoimmunology: Interplay between the immune system and bone metabolism. Ann. Rev. Immunol. 2006, 24, 33–63. [Google Scholar] [CrossRef] [Green Version]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through rankl expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Piemontese, M.; Thostenson, J.D.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Osteocyte-derived rankl is a critical mediator of the increased bone resorption caused by dietary calcium deficiency. Bone 2014, 66, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Takegahara, N.; Kim, H.; Mizuno, H.; Sakaue-Sawano, A.; Miyawaki, A.; Tomura, M.; Kanagawa, O.; Ishii, M.; Choi, Y. Involvement of receptor activator of nuclear factor-kappab ligand (rankl)-induced incomplete cytokinesis in the polyploidization of osteoclasts. J. Biol. Chem. 2016, 291, 3439–3454. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, M.B.; Nakamura, M.C. A comprehensive review of immunoreceptor regulation of osteoclasts. Clin. Rev. Allergy Immunol. 2016, 51, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Takeshita, S. The role of osteoclast differentiation and function in skeletal homeostasis. J. Biochem. 2016, 159, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, H.; Suzu, S.; Hayashi, Y.; Okada, S. Bt-igsf, a novel immunoglobulin superfamily protein, functions as a cell adhesion molecule. J. Cell Physiol. 2005, 204, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Eom, D.S.; Inoue, S.; Patterson, L.B.; Gordon, T.N.; Slingwine, R.; Kondo, S.; Watanabe, M.; Parichy, D.M. Melanophore migration and survival during zebrafish adult pigment stripe development require the immunoglobulin superfamily adhesion molecule igsf11. PLoS Genet. 2012, 8, e1002899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Takegahara, N.; Walsh, M.C.; Middleton, S.A.; Yu, J.; Shirakawa, J.; Ueda, J.; Fujihara, Y.; Ikawa, M.; Ishii, M.; et al. Igsf11 regulates osteoclast differentiation through association with the scaffold protein psd-95. Bone Res. 2020, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosemeci, A.; Makusky, A.J.; Jankowska-Stephens, E.; Yang, X.; Slotta, D.J.; Markey, S.P. Composition of the synaptic psd-95 complex. Mol. Cell Proteom. 2007, 6, 1749–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orian-Rousseau, V. Cd44 acts as a signaling platform controlling tumor progression and metastasis. Front. Immunol. 2015, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Senbanjo, L.T.; Chellaiah, M.A. Cd44: A multifunctional cell surface adhesion receptor is a regulator of progression and metastasis of cancer cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Goodison, S.; Urquidi, V.; Tarin, D. Cd44 cell adhesion molecules. Mol. Pathol. 1999, 52, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. Cd44 is the principal cell surface receptor for hyaluronate. Cell 1990, 61, 1303–1313. [Google Scholar] [CrossRef]
- Du, Y.; Cao, M.; Liu, Y.; He, Y.; Yang, C.; Wu, M.; Zhang, G.; Gao, F. Low-molecular-weight hyaluronan (lmw-ha) accelerates lymph node metastasis of melanoma cells by inducing disruption of lymphatic intercellular adhesion. Oncoimmunology 2016, 5, e1232235. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, W.; Takahashi, T.; Kanno, T.; Ichimiya, H.; Takano, H.; Koseki, T.; Nishihara, T. Mechanisms involved in enhancement of osteoclast formation and function by low molecular weight hyaluronic acid. J. Biol. Chem. 2005, 280, 18967–18972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan in tissue injury and repair. Ann. Rev. Cell Dev. Biol. 2007, 23, 435–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalkanen, S.; Jalkanen, M. Lymphocyte cd44 binds the cooh-terminal heparin-binding domain of fibronectin. J. Cell Biol. 1992, 116, 817–825. [Google Scholar] [CrossRef]
- Faassen, A.E.; Schrager, J.A.; Klein, D.J.; Oegema, T.R.; Couchman, J.R.; McCarthy, J.B. A cell surface chondroitin sulfate proteoglycan, immunologically related to cd44, is involved in type i collagen-mediated melanoma cell motility and invasion. J. Cell Biol. 1992, 116, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Knutson, J.R.; Iida, J.; Fields, G.B.; McCarthy, J.B. Cd44/chondroitin sulfate proteoglycan and alpha 2 beta 1 integrin mediate human melanoma cell migration on type iv collagen and invasion of basement membranes. Mol. Biol. Cell 1996, 7, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-ligand interaction between cd44 and osteopontin (eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Kajita, M.; Itoh, Y.; Chiba, T.; Mori, H.; Okada, A.; Kinoh, H.; Seiki, M. Membrane-type 1 matrix metalloproteinase cleaves cd44 and promotes cell migration. J. Cell Biol. 2001, 153, 893–904. [Google Scholar] [CrossRef]
- Miletti-Gonzalez, K.E.; Murphy, K.; Kumaran, M.N.; Ravindranath, A.K.; Wernyj, R.P.; Kaur, S.; Miles, G.D.; Lim, E.; Chan, R.; Chekmareva, M.; et al. Identification of function for cd44 intracytoplasmic domain (cd44-icd): Modulation of matrix metalloproteinase 9 (mmp-9) transcription via novel promoter response element. J. Biol. Chem. 2012, 287, 18995–19007. [Google Scholar] [CrossRef] [Green Version]
- Sterling, H.; Saginario, C.; Vignery, A. Cd44 occupancy prevents macrophage multinucleation. J. Cell Biol. 1998, 143, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Vignery, A. Osteoclasts and giant cells: Macrophage-macrophage fusion mechanism. Int. J. Exp. Pathol. 2000, 81, 291–304. [Google Scholar] [CrossRef]
- Cui, W.; Ke, J.Z.; Zhang, Q.; Ke, H.Z.; Chalouni, C.; Vignery, A. The intracellular domain of cd44 promotes the fusion of macrophages. Blood 2006, 107, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Kania, J.R.; Kehat-Stadler, T.; Kupfer, S.R. Cd44 antibodies inhibit osteoclast formation. J. Bone Miner. Res. 1997, 12, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Fiorino, C.; Harrison, R.E. E-cadherin is important for cell differentiation during osteoclastogenesis. Bone 2016, 86, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Ishii, M.; Iwai, K.; Koike, M.; Ohshima, S.; Kudo-Tanaka, E.; Ishii, T.; Mima, T.; Katada, Y.; Miyatake, K.; Uchiyama, Y.; et al. Rankl-induced expression of tetraspanin cd9 in lipid raft membrane microdomain is essential for cell fusion during osteoclastogenesis. J. Bone Miner. Res. 2006, 21, 965–976. [Google Scholar] [CrossRef]
- Lundberg, P.; Koskinen, C.; Baldock, P.A.; Lothgren, H.; Stenberg, A.; Lerner, U.H.; Oldenborg, P.A. Osteoclast formation is strongly reduced both in vivo and in vitro in the absence of cd47/sirpalpha-interaction. Biochem. Biophys. Res. Commun. 2007, 352, 444–448. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, G.; Sun, W.; Zhao, C.; Zhang, P.; Song, J.; Zhao, D.; Jin, X.; Li, Q.; Ling, S.; et al. Cd44 deficiency inhibits unloading-induced cortical bone loss through downregulation of osteoclast activity. Sci. Rep. 2015, 5, 16124. [Google Scholar] [CrossRef]
- de Vries, T.J.; Schoenmaker, T.; Beertsen, W.; van der Neut, R.; Everts, V. Effect of cd44 deficiency on in vitro and in vivo osteoclast formation. J. Cell Biochem. 2005, 94, 954–966. [Google Scholar] [CrossRef]
- Hayer, S.; Steiner, G.; Gortz, B.; Reiter, E.; Tohidast-Akrad, M.; Amling, M.; Hoffmann, O.; Redlich, K.; Zwerina, J.; Skriner, K.; et al. Cd44 is a determinant of inflammatory bone loss. J. Exp. Med. 2005, 201, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Schmits, R.; Filmus, J.; Gerwin, N.; Senaldi, G.; Kiefer, F.; Kundig, T.; Wakeham, A.; Shahinian, A.; Catzavelos, C.; Rak, J.; et al. Cd44 regulates hematopoietic progenitor distribution, granuloma formation, and tumorigenicity. Blood 1997, 90, 2217–2233. [Google Scholar] [CrossRef] [Green Version]
- Protin, U.; Schweighoffer, T.; Jochum, W.; Hilberg, F. Cd44-deficient mice develop normally with changes in subpopulations and recirculation of lymphocyte subsets. J. Immunol. 1999, 163, 4917–4923. [Google Scholar]
- Jang, S.; Oh, D.; Lee, Y.; Hosy, E.; Shin, H.; van Riesen, C.; Whitcomb, D.; Warburton, J.M.; Jo, J.; Kim, D.; et al. Synaptic adhesion molecule igsf11 regulates synaptic transmission and plasticity. Nat. Neurosci. 2016, 19, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier, C.; Laursen, L.; Jemth, P.; Gianni, S. Seeking allosteric networks in pdz domains. Protein Eng. Des. Sel. 2018, 31, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, T.; Jeong, B.C.; Cho, I.T.; Han, D.; Takegahara, N.; Negishi-Koga, T.; Takayanagi, H.; Lee, J.H.; Sul, J.Y.; et al. Tmem64 modulates calcium signaling during rankl-mediated osteoclast differentiation. Cell Metab. 2013, 17, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Takegahara, N.; Walsh, M.C.; Choi, Y. CD44 Can Compensate for IgSF11 Deficiency by Associating with the Scaffold Protein PSD-95 during Osteoclast Differentiation. Int. J. Mol. Sci. 2020, 21, 2646. https://doi.org/10.3390/ijms21072646
Kim H, Takegahara N, Walsh MC, Choi Y. CD44 Can Compensate for IgSF11 Deficiency by Associating with the Scaffold Protein PSD-95 during Osteoclast Differentiation. International Journal of Molecular Sciences. 2020; 21(7):2646. https://doi.org/10.3390/ijms21072646
Chicago/Turabian StyleKim, Hyunsoo, Noriko Takegahara, Matthew C. Walsh, and Yongwon Choi. 2020. "CD44 Can Compensate for IgSF11 Deficiency by Associating with the Scaffold Protein PSD-95 during Osteoclast Differentiation" International Journal of Molecular Sciences 21, no. 7: 2646. https://doi.org/10.3390/ijms21072646
APA StyleKim, H., Takegahara, N., Walsh, M. C., & Choi, Y. (2020). CD44 Can Compensate for IgSF11 Deficiency by Associating with the Scaffold Protein PSD-95 during Osteoclast Differentiation. International Journal of Molecular Sciences, 21(7), 2646. https://doi.org/10.3390/ijms21072646