Functional Analysis of IRF1 Reveals its Role in the Activation of the Type I IFN Pathway in Golden Pompano, Trachinotus ovatus (Linnaeus 1758)



Abstract
:1. Introduction
2. Results
2.1. Sequence Characterization of ToIRF1
2.2. ToIRF1 Structural and Phylogenetic Analysis
2.3. Tissue Expression of ToIRF1
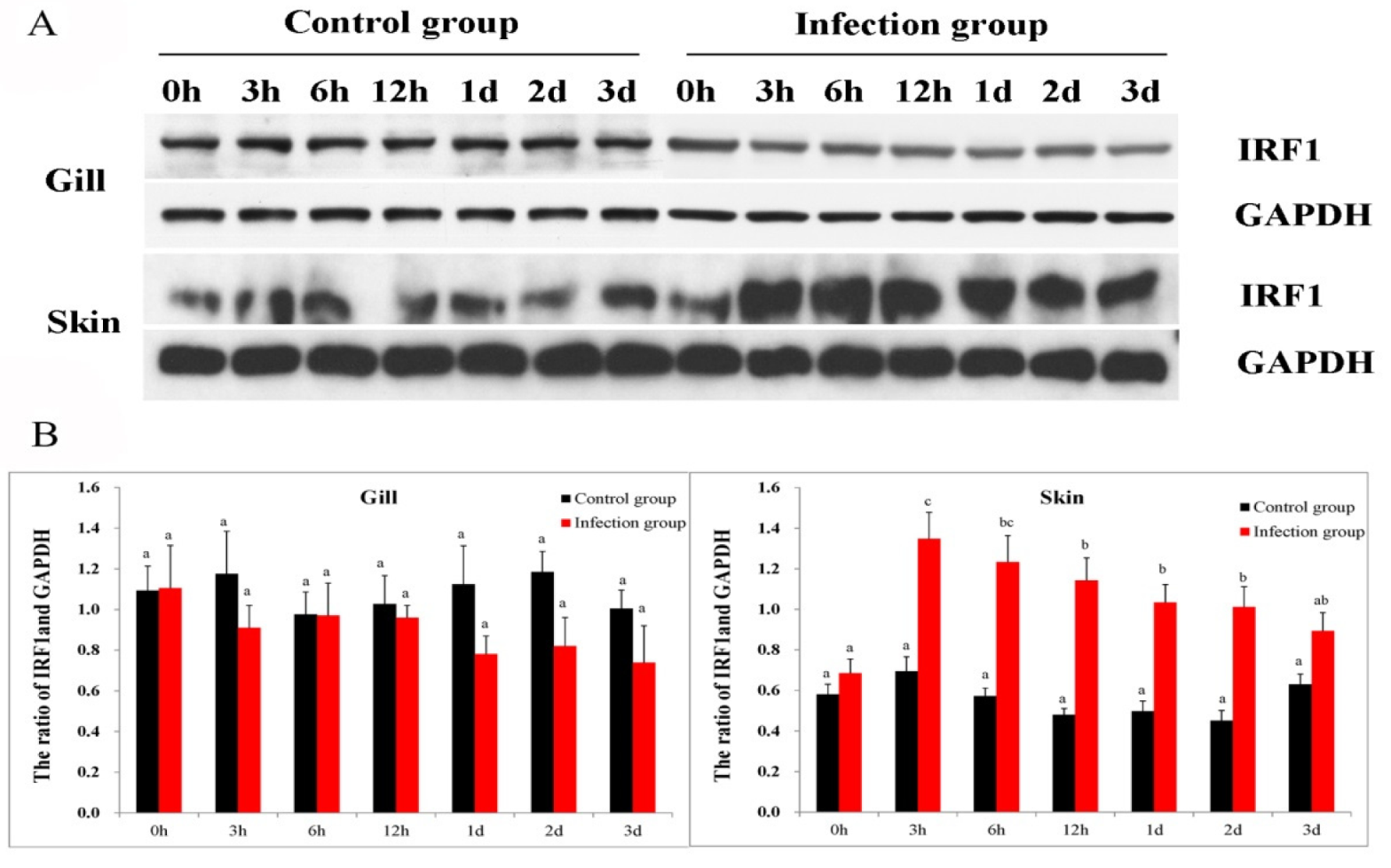
2.4. Protein Expression Pattern after C. irritans Infection
2.5. Cytosol and Nucleus Distribution of ToIRF1
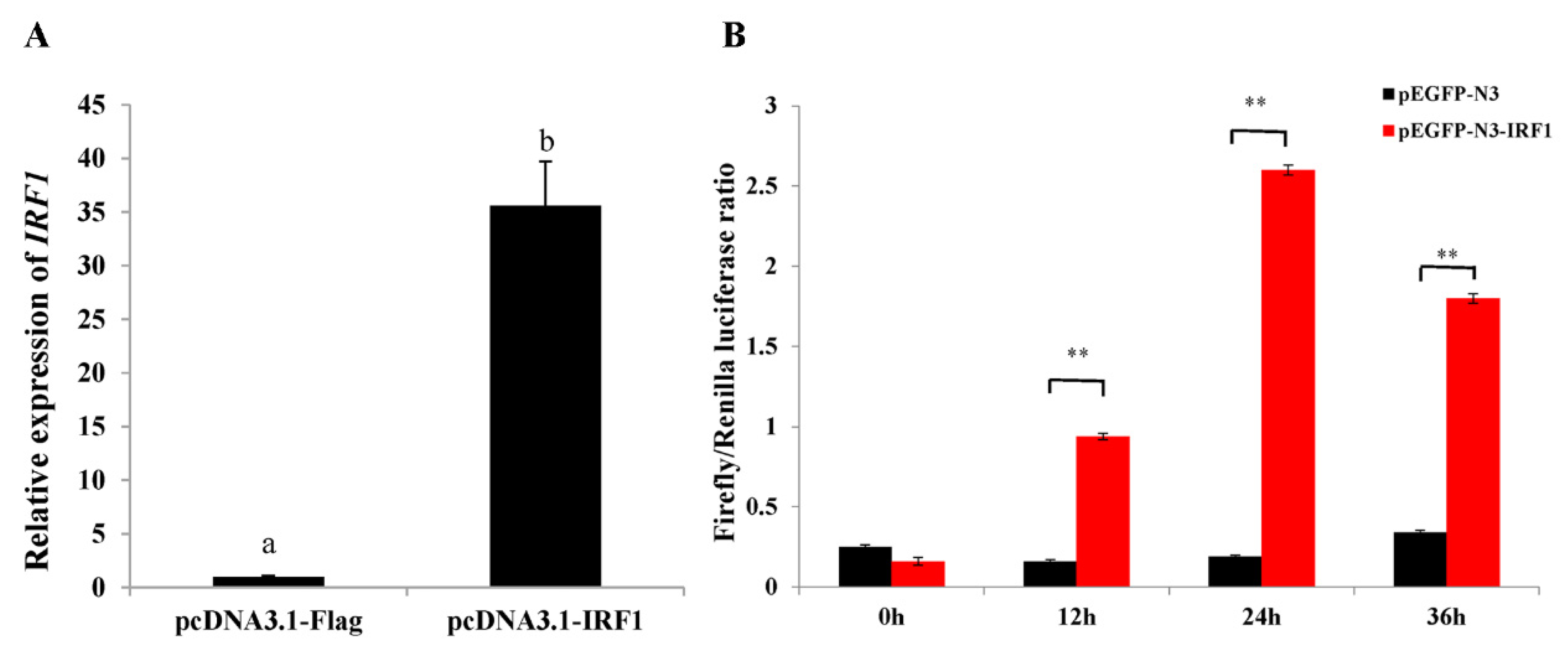
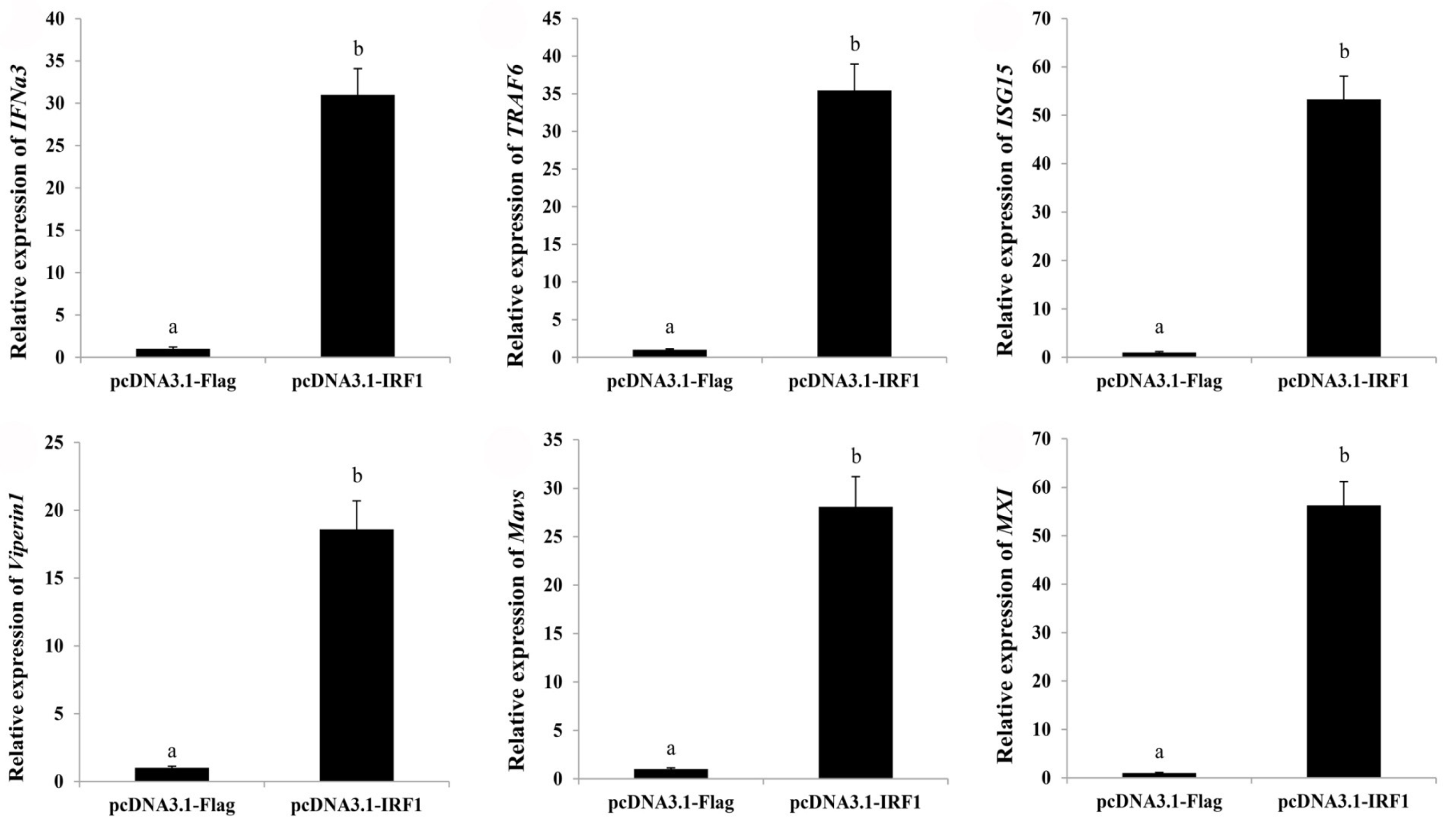
2.6. Ectopic Expression of ToIRF1 Positively Promotes ToIFNa3 Expression and Interferon Immune Response
2.7. Activation of the Type I IFN Response by T. ovatus rIFN
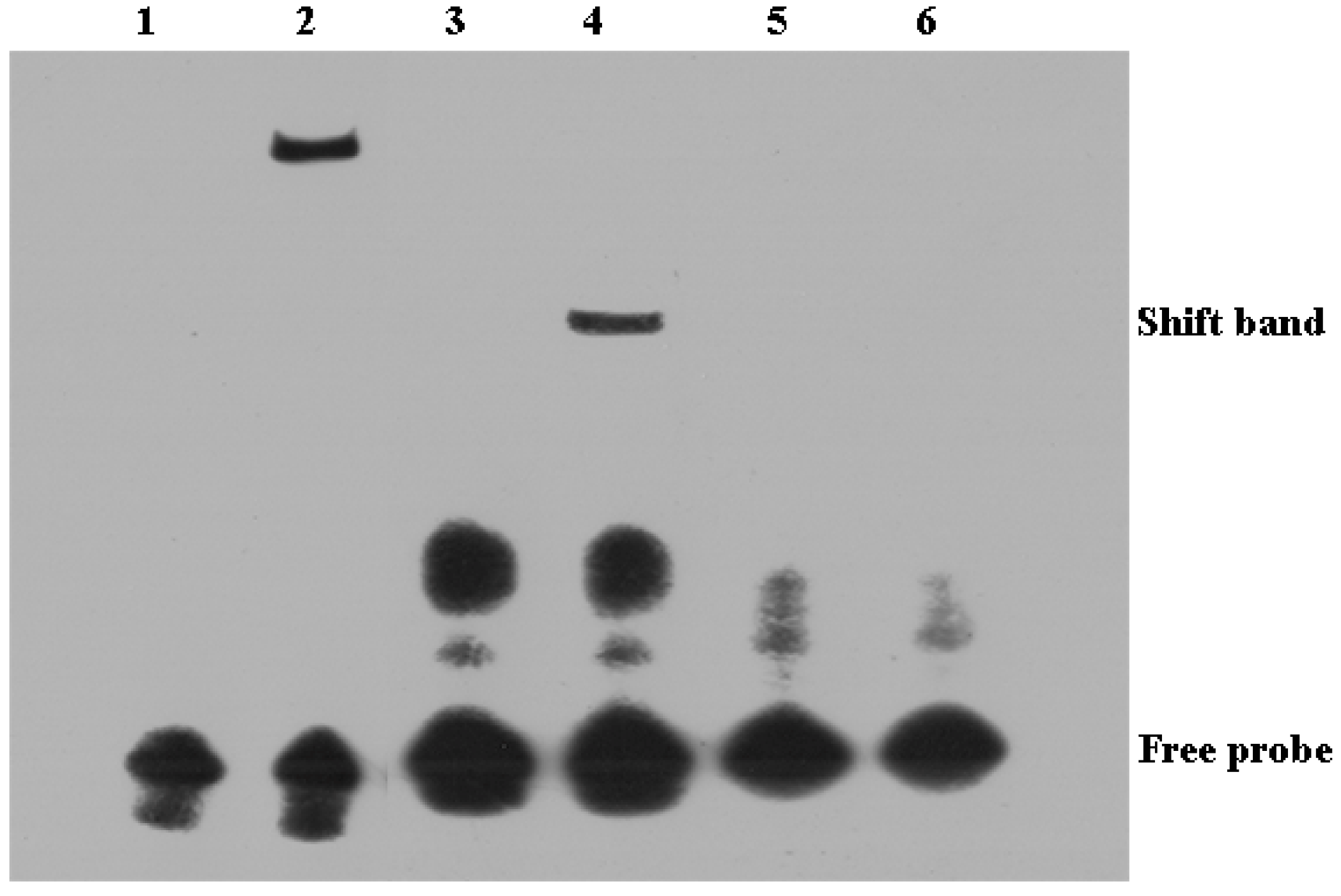
2.8. Binding of ToIRF1 to the ToIFNa3 Sequence
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. C. irritans Challenge and Sampling
4.3. RNA Isolation, cDNA Synthesis, and Protein Extraction
4.4. Cloning of cDNA and Genomic Sequences
4.5. Bioinformatics
4.6. Subcellular Localization
4.7. ToIRF1 Overexpression Promotes ToIFNa3 Expression
4.8. Promoter Deletion Mutation and Point Mutation Analysis
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Quantitative Real-Time PCR and Statistical Analysis
4.11. Preparation of the IRF1 Polyclonal Antibody and Western Blotting Analysis
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| IRF1 | Interferon regulatory factor 1 |
| ORF | Open reading frame |
| DBD | DNA-binding domain |
| ISREs | Interferon stimulating response elements |
| EMSA | Electrophoretic mobile shift assays |
| IFN | Interferon |
| IAD | IRF-associated domain |
| ML | Maximum likelihood |
| GPS | Golden pompano T. ovatus snout cell |
| TRAF6 | TNF receptor-associated factor 6 |
| ISG15 | Interferon stimulated gene |
| Mavs | Mitochondrial antiviral signaling protein |
| MXI | MAX interactor 1 |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| TBST | Tris-buffered saline and Tween-20 |
References
- Mamane, Y.; Heylbroeck, C.; Génin, P.; Algarté, M.; Servant, M.J.; LePage, C.; DeLuca, C.; Kwon, H.; Lin, R.; Hiscott, J. Interferon regulatory factors: The next generation. Gene 1999, 237, 1–14. [Google Scholar] [CrossRef]
- Paun, A.; Pitha, P.M. The IRF family, revisited. Biochimie 2007, 89, 744–753. [Google Scholar] [CrossRef]
- Savitsky, D.; Tamura, T.; Yanai, H.; Taniguchi, T. Regulation of immunity and oncogenesis by the IRF transcription factor family. Cancer Immunol. Immunother. 2010, 59, 489. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.; Blaszczyk, K.; Wesoly, J.; Bluyssen, H.A.R. A positive feedback amplifier circuit that regulates interferon (IFN)-Stimulated gene expression and controls type I and type II IFN responses. Front. Immunol. 2018, 28, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.C.; Liu, B.S.; Zhang, N.; Guo, H.Y.; Guo, L.; Jiang, S.G.; Zhang, D.C. Interferon regulatory factor 2 plays a positive role in interferon gamma expression in golden pompano, Trachinotus ovatus (Linnaeus 1758). Fish Shellfish Immunol. 2020, 96, 107–113. [Google Scholar] [CrossRef]
- Huang, B.; Qi, Z.T.; Xu, Z.; Nie, P. Global characterization of interferon regulatory factor (IRF) genes in vertebrates: Glimpse of the diversification in evolution. BMC Immunol. 2010, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Yanai, H.; Negishi, H.; Taniguchi, T. The IRF family of transcription factors: Inception, impact and implications in oncogenesis. OncoImmunology 2012, 1, 1376–1386. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Cui, J.; Song, W.; Zhao, X.; Xu, T. The evolution and functional characterization of miiuy croaker interferon regulatory factor 9 involved in immune response. Fish Shellfish Immunol. 2017, 66, 524–530. [Google Scholar] [CrossRef]
- Stein, C.; Caccamo, M.; Laird, G.; Leptin, M. Conservation and divergence of gene families encoding components of innate immune response systems in zebrafish. Genome Biol. 2007, 8, 251. [Google Scholar] [CrossRef] [Green Version]
- Shu, C.; Sun, Y.Y.; Xu, T.J. Molecular Characterization of Three IRF1 Subfamily Members Reveals Evolutionary Significance of IRF11 in Miiuy Croaker. Dev. Comp. Immunol. 2015, 53, 385–391. [Google Scholar] [CrossRef]
- Huang, W.S.; Zhu, M.H.; Chen, S.; Wang, Z.X.; Liang, Y.; Huang, B.; Nie, P. Molecular cloning and expression analysis of a fish specific interferon regulatory factor, IRF11, in orange spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2017, 60, 368–379. [Google Scholar] [CrossRef]
- Tanaka, N.; Kawakami, T.; Taniguchi, T. Recognition DNA sequences of interferon regulatory factor 1 (IRF-1) and IRF-2, regulators of cell growth and the interferon system. Mol. Cell. Biol. 1993, 13, 4531–4538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, Y.; Shimizu, T.; Kusumoto, M.; Kyogoku, Y.; Taniguchi, T.; Hakoshima, T. Crystal structure of an IRF-DNA complex reveals novel DNA recognition and cooperative binding to a tandem repeat of core sequences. EMBO J. 1999, 18, 5028–5041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Lamphier, M.S.; Fujita, T.; Taniguchi, T.; Harada, H. The oncogenic transcription factor IRF-2 possesses a transcriptional repression and a latent activation domain. Oncogene 1994, 9, 1423–1428. [Google Scholar] [PubMed]
- Lohoff, M.; Mak, T.W. Roles of interferon-regulatory factors in T-helper-cell differentiation. Nat. Rev. Immunol. 2005, 5, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.J.; Field, A.E.; Pitha, P.M. Virus-induced heterodimer formation between IRF-5 and IRF-7 modulates assembly of the IFNA enhanceosome in vivo and transcriptional activity of IFNA genes. J. Biol. Chem. 2003, 278, 16630–16641. [Google Scholar] [CrossRef] [Green Version]
- Barnes, B.J.; Richards, J.; Mancl, M.; Hanash, S.; Beretta, L.; Pitha, P.M. Global and distinct targets of IRF-5 and IRF-7 during innate response to viral infection. J. Biol. Chem. 2004, 279, 45194–45207. [Google Scholar] [CrossRef] [Green Version]
- Stellacci, E.; Testa, U.; Petrucci, E.; Benedetti, E.; Orsatti, R.; Feccia, T.; Stafsnes, M.; Coccia, E.M.; Marziali, G.; Battistini, A. Interferon regulatory factor-2 drives megakaryocytic differentiation. Biochem. J. 2004, 377, 367–378. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Tafalla, C.; Truckle, J.; Secombes, C.J. Identification of a second group of type I IFNs in fish sheds light on IFN evolution in vertebrates. J. Immunol. 2007, 179, 3859–3871. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Munoz, A.; Roca, F.J.; Meseguer, J.; Mulero, V. New insights into the evolution of IFNs: Zebrafish group II IFNs induce a rapid and transient expression of IFN-dependent genes and display powerful antiviral activities. J. Immunol. 2009, 182, 3440–3449. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Gorgoglione, B.; Taylor, N.G.; Summathed, T.; Lee, P.T.; Panigrahi, A. Salmonids have an extraordinary complex type I IFN system: Characterization of the IFN locus in rainbow trout Oncorhynchus mykiss reveals two novel IFN subgroups. J. Immunol. 2014, 193, 2273–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Z.; Chen, S.N.; Huang, B.; Zou, J.; Nie, P. Fish type I and type II interferons: Composition, receptor usage, production and function. Rev. Aquac. 2019, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.M.; Yang, C.Y.; Li, M.J.; Yi, Q.L.; Lu, G.X.; Wu, Y.C.; Qu, C.; Wang, L.L.; Song, L.S. A conserved interferon regulation factor 1 (IRF-1) from Pacific oyster Crassostrea gigas functioned as an activator of IFN pathway. Fish Shellfish Immunol. 2018, 76, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Hiscott, J.; Pitha, P.M. The growing family of interferon regulatory factors. Cytokine Growth Factor Rev. 1997, 8, 293–312. [Google Scholar] [CrossRef]
- Ding, Y.; Ao, J.; Huang, X.; Chen, X.H. Identification of two subgroups of type I IFNs in perciforme fish large yellow croaker Larimichthys crocea provides novel insights into function and regulation of fish type I IFNs. Front. Immunol. 2016, 7, 343. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T.; Sakakibara, J.; Sudo, Y.; Miyamoto, M.; Kimura, Y.; Taniguchi, T. Evidence for a nuclear factor(s) IRF-1, mediating induction and silencing properties to human IFN-beta gene regulatory elements. EMBO J. 1988, 7, 3397–3405. [Google Scholar] [CrossRef] [PubMed]
- Hochhaus, A.; Yan, X.H.; Willer, A.; Hehlmann, R.; Gordon, M.Y.; Goldman, J.M.; Melo, J.V. Expression of interferon regulatory factor (IRF) genes and response to interferon-a in chronic myeloid leukaemia. Leukemia 1997, 11, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.C.; Guo, H.Y.; Zhang, N.; Guo, L.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Functional characterization of interferon regulatory factor 2 and its role in the transcription of interferon a3 in golden pompano Trachinotus ovatus (Linnaeus 1758). Fish Shellfish Immunol. 2019, 93, 90–98. [Google Scholar] [CrossRef]
- Zhu, K.C.; Guo, H.Y.; Zhang, N.; Guo, L.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Structural and expression analysis of golden pompano Trachinotus ovatus IRF5 and its role in regulation of type I IFN. Fish Shellfish Immunol. 2020, 97, 313–321. [Google Scholar] [CrossRef]
- Yuan, S.; Zheng, T.; Li, P.; Yang, R.; Ruan, J.; Huang, S.; Wu, Z.; Xu, A. Characterization of amphioxus IFN regulatory factor family reveals an archaic signaling framework for innate immune response. J. Immunol. 2015, 195, 5657–5666. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.Y.; Guo, H.Y.; Zhu, C.Y.; Ma, Z.H.; Jiang, S.G.; Zhang, D.C. Genetic polymorphism of breeding populations of golden pompano (Trachinotus ovatus). South China Fish. Sci. 2014, 10, 67–71. [Google Scholar]
- Zhen, P.L.; Ma, Z.H.; Guo, H.Y.; Jiang, S.G.; Zhang, D.C. Ontogenetic development of caudal skeletons in Trachinotus ovatus larvae. South China Fish. Sci. 2014, 10, 45–50. [Google Scholar]
- Colorni, A.; Burgess, P. Cryptocaryon irritans Brown 1951, the cause of ‘white spot disease’ in marine fish: Anupdate. Aquar. Sci. Conserv. 1997, 1, 217–238. [Google Scholar] [CrossRef]
- Dan, X.M.; Li, A.X.; Lin, X.T.; Teng, N.; Zhu, X.Q. A standardized method to propagate Cryptocaryon irritans, on a susceptible host pompano Trachinotus ovatus. Aquaculture 2006, 258, 127–133. [Google Scholar] [CrossRef]
- Nigrelli, R.D.; Ruggieri, G.D. Enzootics in the New York aquarium caused by Cryptocaryon irritans Brown, 1951 (=Ichthyophthirius marinuss Sikama, 1961), a histophagous ciliate in the skin, eyes and gills of marine fishes. Zoologica 1966, 51, 97–102. [Google Scholar]
- Yoshinaga, T. Controlling of Cryptocaryon irritans. Kaiyo. Mon. 1998, 14, 73–76. [Google Scholar]
- Ordás, M.C.; Abollo, E.; Costa, M.M.; Figueras, A.; Novoa, B. Molecular Cloning and Expression Analysis of Interferon Regulatory factor-1 (IRF-1) of Turbot and Sea Bream. Mol. Immunol. 2006, 43, 882–890. [Google Scholar] [CrossRef]
- Bergan, V.; Kileng, Ø.; Sun, B.J.; Robertsen, B. Regulation and function of interferon regulatory factors of Atlantic salmon. Mol. Immunol. 2010, 47, 2005–2014. [Google Scholar] [CrossRef]
- Yao, C.L.; Kong, P.; Huang, X.N.; Wang, Z.Y. Molecular cloning and expression of IRF1 in large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol. 2010, 28, 654–660. [Google Scholar] [CrossRef]
- Gan, X.N.; Chen, Z.; Wang, X.Z.; Wang, D.Q.; Chen, X.W. Molecular cloning and characterization of interferon regulatory factor 1 (IRF-1), IRF-2 and IRF-5 in the chondrostean paddlefish Polyodon spathula and their phylogenetic importance in the Osteichthyes. Dev. Comp. Immunol. 2012, 36, 74–84. [Google Scholar]
- Lu, Y.; Wang, Q.L.; Liu, Y.; Shao, C.W.; Chen, S.L.; Sha, Z.X. Gene cloning and expression analysis of IRF1 in half-smooth tongue sole (Cynoglossus semilaevis). Mol. Biol. Rep. 2014, 41, 4093–4101. [Google Scholar] [CrossRef] [PubMed]
- Breathnach, R.; Chambon, P. Organization and expression of eukaryotic split genes coding for proteins. Annu. Rev. Biochem. 1981, 50, 349–383. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Nakayama, K.; Penninger, J.; Kitagawa, M.; Harada, H.; Matsuyama, T.; Tanaka, N.; Kamijo, R.; Vilcek, J.; Mak, T.W. Involvement of the IRF-1 transcription factor in antiviral responses to interferons. Science 1994, 264, 1921–1924. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Yanai, H.; Savitsky, D.; Taniguchi, T. The IRF family transcription factors in immunity and oncogenesis. Annu. Rev. Immunol. 2008, 26, 535–584. [Google Scholar] [CrossRef]
- Kung, H.C.; Evensen, Ø.; Hong, J.R.; Kuo, C.Y.; Tso, C.H.; Ngou, F.H.; Lu, M.W.; Wu, J.L. Interferon Regulatory factor-1 (IRF-1) Is Involved in the Induction of Phosphatidylserine Receptor (PSR) in Response to dsRNA Virus Infection and Contributes to Apoptotic Cell Clearance in CHSE-214 Cell. Int. J. Mol. Sci. 2014, 15, 19281–19306. [Google Scholar] [CrossRef]
- Shi, Y.; Zhu, X.P.; Yin, J.K.; Zhang, Q.Y.; Gui, J.F. Identification and characterization of interferon regulatory factor-1 from orange-spotted grouper (Epinephelus coioides). Mol. Biol. Rep. 2010, 37, 1483–1493. [Google Scholar] [CrossRef]
- Sun, B.J.; Chang, M.X.; Song, Y.; Yao, W.J.; Nie, P. Gene structure and transcription of IRF-1 and IRF-7 in the mandarin fish Siniperca chuatsi. Vet. Immunol. Immunopathol. 2007, 116, 26–36. [Google Scholar] [CrossRef]
- Escalante, C.R.; Yie, J.; Thanos, D.; Aggarwal, A.K. Structure of IRF-1 with bound DNA reveals determinants of interferon regulation. Nature 1998, 391, 103–106. [Google Scholar] [CrossRef]
- Gupta, M.; Rath, P.C. Interferon regulatory factor-1 (IRF-1) interacts with regulated in development and DNA damage response 2 (REDD2) in the cytoplasm of mouse bone marrow cells. Int. J. Biol. Macromol. 2014, 65, 41–50. [Google Scholar] [CrossRef]
- Khoo, C.K.; Abdul-Murad, A.M.; Kua, B.C.; Mohd-Adnan, A. Cryptocaryon irritans, infection induces the acute phase response in Lates calcarifer: A transcriptomic perspective. Fish Shellfish Immunol. 2012, 33, 788–794. [Google Scholar] [CrossRef]
- Negishi, H.; Fujita, Y.; Yanai, H.; Sakaguchi, S.; Ouyang, X.S.; Shinohara, M.; Takayanagi, H.; Ohba, Y.; Taniguchi, T.; Honda, K. Evidence for licensing of IFN-γ-induced IFN regulatory factor 1 transcription factor by MyD88 in Toll-like receptor-dependent gene induction program. Proc. Natl. Acad. Sci. USA 2006, 103, 15136–15141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamane, D.; Feng, H.; Rivera-Serrano, E.E.; Selitsky, S.R.; Hirai-Yuki, A.; Das, A.; McKnight, K.L.; Misumi, I.; Hensley, L.; Lovell, W.; et al. Basal Expression of Interferon Regulatory Factor 1 Drives Intrinsic Hepatocyte Resistance to Multiple RNA Viruses. Nat. Microbiol. 2019, 4, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Lai, Q.; Lin, G.; Ma, M.; Huang, S.; Li, W.; Li, D.; Gu, M.; Mao, H.; Hu, C. IRF-1 acts as a positive regulator in the transcription of grass carp (Ctenopharyngodon idella) IFN gene. Fish Shellfish Immunol. 2013, 34, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.C.; Guo, H.Y.; Zhang, N.; Liu, B.S.; Guo, L.; Jiang, S.G.; Zhang, D.C. Functional characterization of IRF8 regulation of type II IFN in golden pompano (Trachinotus ovatus). Fish Shellfish Immunol. 2019, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Li, J.; Lu, N.; Shen, X.Z. Interferon regulatory factors: A key to tumour immunity. Int. Immunopharmacol. 2017, 49, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Zhang, Y.B.; Zhang, Q.M.; Li, Z.; Zhang, Q.Y.; Gui, J.F. Zebrafish IRF1 regulates IFN antiviral response through binding to IFNϕ1 and IFNϕ3 promoters downstream of MyD88 signaling. J. Immunol. 2015, 194, 1225–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C.; Guo, L.; Guo, H.Y.; Zhu, K.C.; Li, S.Q.; Zhang, Y.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Li, J.T. Chromosome-level genome assembly of golden pompano (Trachinotus ovatus) in the family Carangidae. Sci. Data 2019, 6, 216. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D. MEGA 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 3, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wei, S.; Wang, Z.; Huang, X.; Huang, Y.; Cai, J.; Li, C.; Qin, Q. Establishment of a new cell line from the snout tissue of golden pompano Trachinotus ovatus, and its application in virus susceptibility. J. Fish Biol. 2016, 88, 2251–2262. [Google Scholar] [CrossRef]
- Zhu, K.C.; Song, L.; Guo, H.Y.; Guo, L.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Elovl4a participates in LC-PUFA biosynthesis and is regulated by PPARαβ in golden pompano Trachinotus ovatus (Linnaeus 1758). Sci. Rep. 2019, 9, 4684. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wan, X.L.; Huang, C.X.; Wang, W.M.; Liu, H.; Wang, H.L. Study on the immune response to recombinant Hsp70 protein from Megalobrama amblycephala. Immunobiology 2014, 219, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; Volume 2. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject and Primers | Nucleotide Sequence |
|---|---|
| Primers for Sequence Cloning | |
| IRF1-ORF-F | CGCGGATCCATGCCTGTGTCTCGGATGA |
| IRF1-ORF-R | CCGCTCGAGTCAGTGGAGGTATGGTTGGCA |
| IRF1-genome-F | GCTCTCATCGTATCGTGT |
| IRF1-genome-R | CCAAACTGGTACAGAGTC |
| IRF1-3′RACE-outer | TGATACTCAGTCTCCCTCGG |
| IRF1-3′RACE-inner | GTTACACAACACTGGGCT |
| IRF1-5′RACE-outer | GCACACAGAGCAGCGAGA |
| IRF1-5′RACE-inner | GAACTGTGACCCAAAGAC |
| Deletion mutant construction | |
| IFNa3-pF1 | CGGGGTACCAAAAGACAACTGATTGTTGA |
| IFNa3-pF2 | CGGGGTACCCTGCTACATATAAAAATGT |
| IFNa3-pF3 | CGGGGTACCCAATGTGAAGAGGGTTCAG |
| IFNa3-pF4 | CGGGGTACCTTTATTTTGTAAAGGTGAGTG |
| IFNa3-pF5 | CGGGGTACCTACTGCACTGGTATCAGTACT |
| IFNa3-pR | CCGCTCGAGCATTGACATGATGCCTAACTCT |
| Primers for qRT-PCR | |
| qRT-IRF1-F | TGATACTCAGTCTCCCTCGG |
| qRT-IRF1-R | TCTCGCTGCTCTGTGTGC |
| qRT-IFNa3-F | ACACTATGGTCACTACAGCAAC |
| qRT-IFNa3-R | ACCTCAGTGTTTCGTATGTG |
| qRT-TRAF6-F | CCCTAAAGCACCCATCGC |
| qRT-TRAF6-R | AAGGTCACGCAGGAACTCAG |
| qRT-MXI-F | CATACCCTTGGGACCTGA |
| qRT-MXI-R | TGCTTTGGCTTTGTTGAGT |
| qRT-ISG15-F | TACGCTGAGTGAGACCCG |
| qRT-ISG15-R | GGAGGAACACCTGGATGG |
| qRT-Viperin1-F | GACCCGTCCAAGTCCATC |
| qRT-Viperin1-R | CAAAGCCACTGAAGCAAAT |
| qRT-Viperin2-F | CCCGAGTCCAATGAGAAGA |
| qRT-Viperin2-R | CGAAGCCACTAAAGCAGATG |
| qRT-Mavs-F | GTTTGGAGGTGCGGATGA |
| qRT-Mavs-R | CCTTTTCGGCTTTGCTGTA |
| EF1α-F | AAGCCAGGTATGGTTGTCAACTTT |
| EF1α-R | CGTGGTGCATCTCCACAGACT |
| EMSA assays | |
| IFNa3-P2-MUT5 | CAGCAAGAATCTGCTGAATGGGAGGAATAT |
| IFNa3-P2-WT5 | CAGCAGAAATCCACTGAGCGGGAAAAATAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.-C.; Zhang, N.; Liu, B.-S.; Guo, L.; Guo, H.-Y.; Jiang, S.-G.; Zhang, D.-C. Functional Analysis of IRF1 Reveals its Role in the Activation of the Type I IFN Pathway in Golden Pompano, Trachinotus ovatus (Linnaeus 1758). Int. J. Mol. Sci. 2020, 21, 2652. https://doi.org/10.3390/ijms21072652
Zhu K-C, Zhang N, Liu B-S, Guo L, Guo H-Y, Jiang S-G, Zhang D-C. Functional Analysis of IRF1 Reveals its Role in the Activation of the Type I IFN Pathway in Golden Pompano, Trachinotus ovatus (Linnaeus 1758). International Journal of Molecular Sciences. 2020; 21(7):2652. https://doi.org/10.3390/ijms21072652
Chicago/Turabian StyleZhu, Ke-Cheng, Nan Zhang, Bao-Suo Liu, Liang Guo, Hua-Yang Guo, Shi-Gui Jiang, and Dian-Chang Zhang. 2020. "Functional Analysis of IRF1 Reveals its Role in the Activation of the Type I IFN Pathway in Golden Pompano, Trachinotus ovatus (Linnaeus 1758)" International Journal of Molecular Sciences 21, no. 7: 2652. https://doi.org/10.3390/ijms21072652
APA StyleZhu, K. -C., Zhang, N., Liu, B. -S., Guo, L., Guo, H. -Y., Jiang, S. -G., & Zhang, D. -C. (2020). Functional Analysis of IRF1 Reveals its Role in the Activation of the Type I IFN Pathway in Golden Pompano, Trachinotus ovatus (Linnaeus 1758). International Journal of Molecular Sciences, 21(7), 2652. https://doi.org/10.3390/ijms21072652