Immediate or Delayed Transplantation of a Vein Conduit Filled with Nasal Olfactory Stem Cells Improves Locomotion and Axogenesis in Rats after a Peroneal Nerve Loss of Substance

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
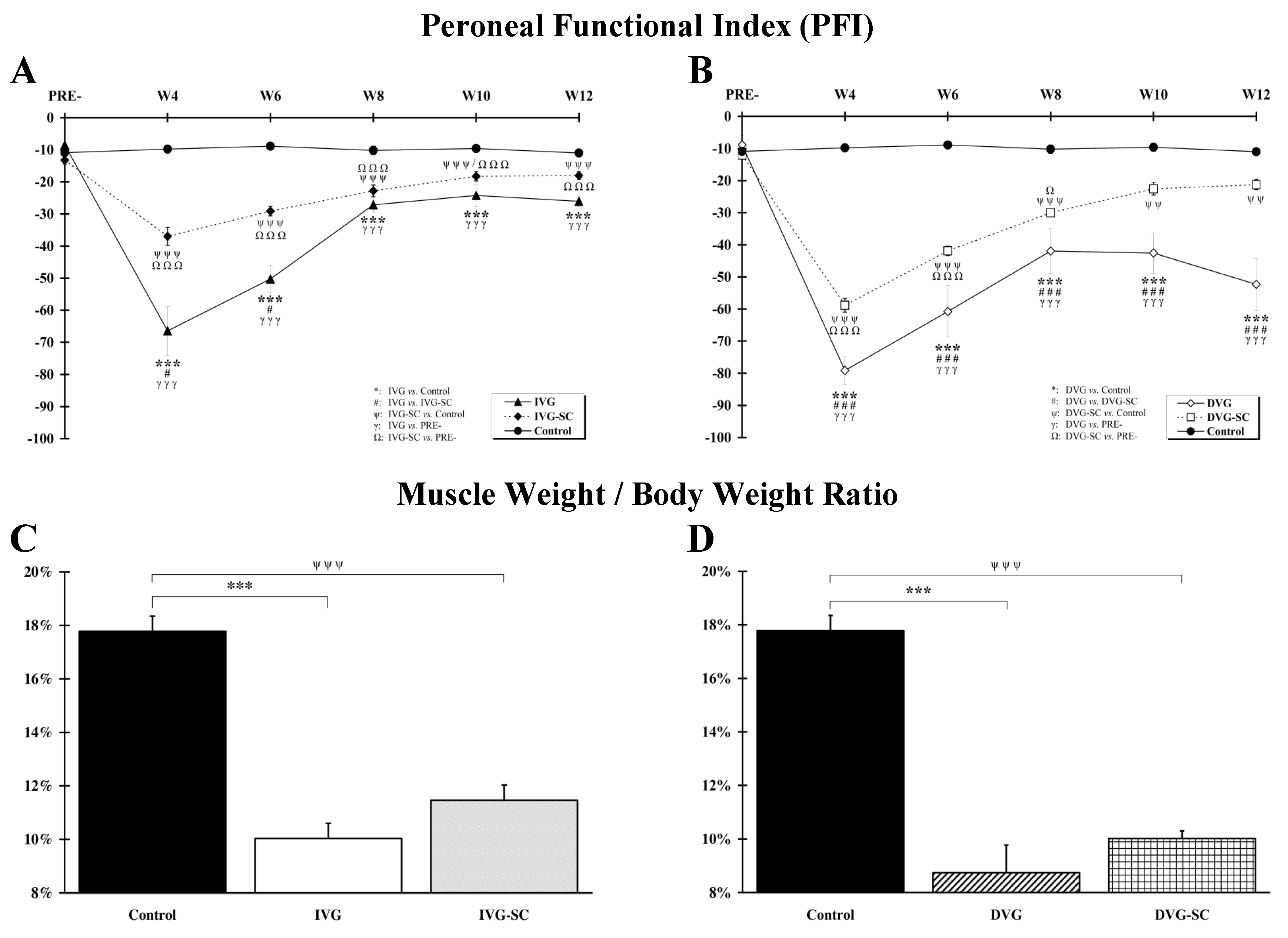
2.1. OEMSCs Improves Locomotion but Not Muscle Weight
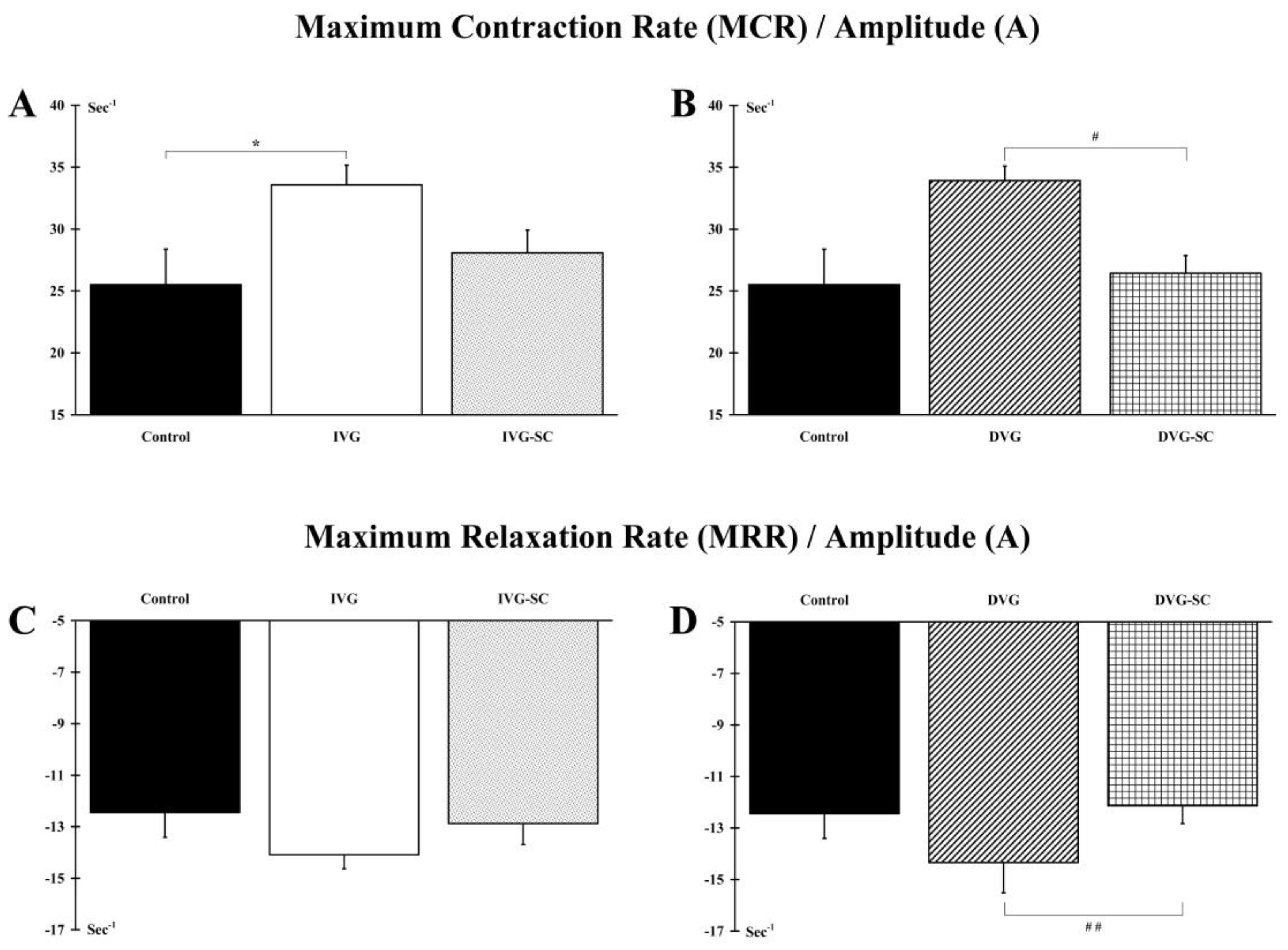
2.2. OEMSCs Partially Maintain the Contractile Phenotype of the Target Muscle
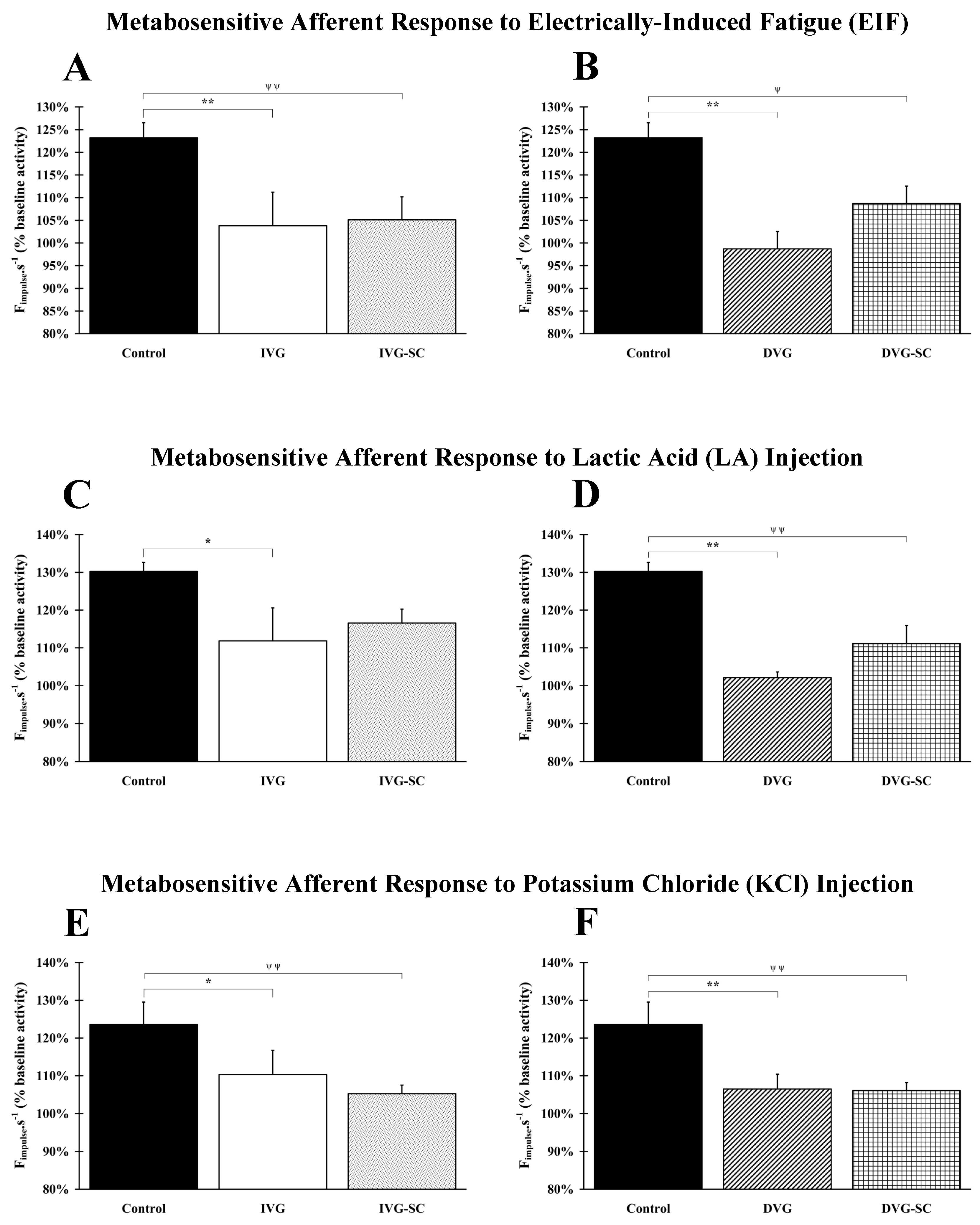
2.3. OEMSCs Do Not Enhance Nerve Afferent Response
2.3.1. Response to Electrically Induced Fatigue
2.3.2. Response to Lactic Acid Injection
2.3.3. Response to Potassium Chloride (KCl) Injection
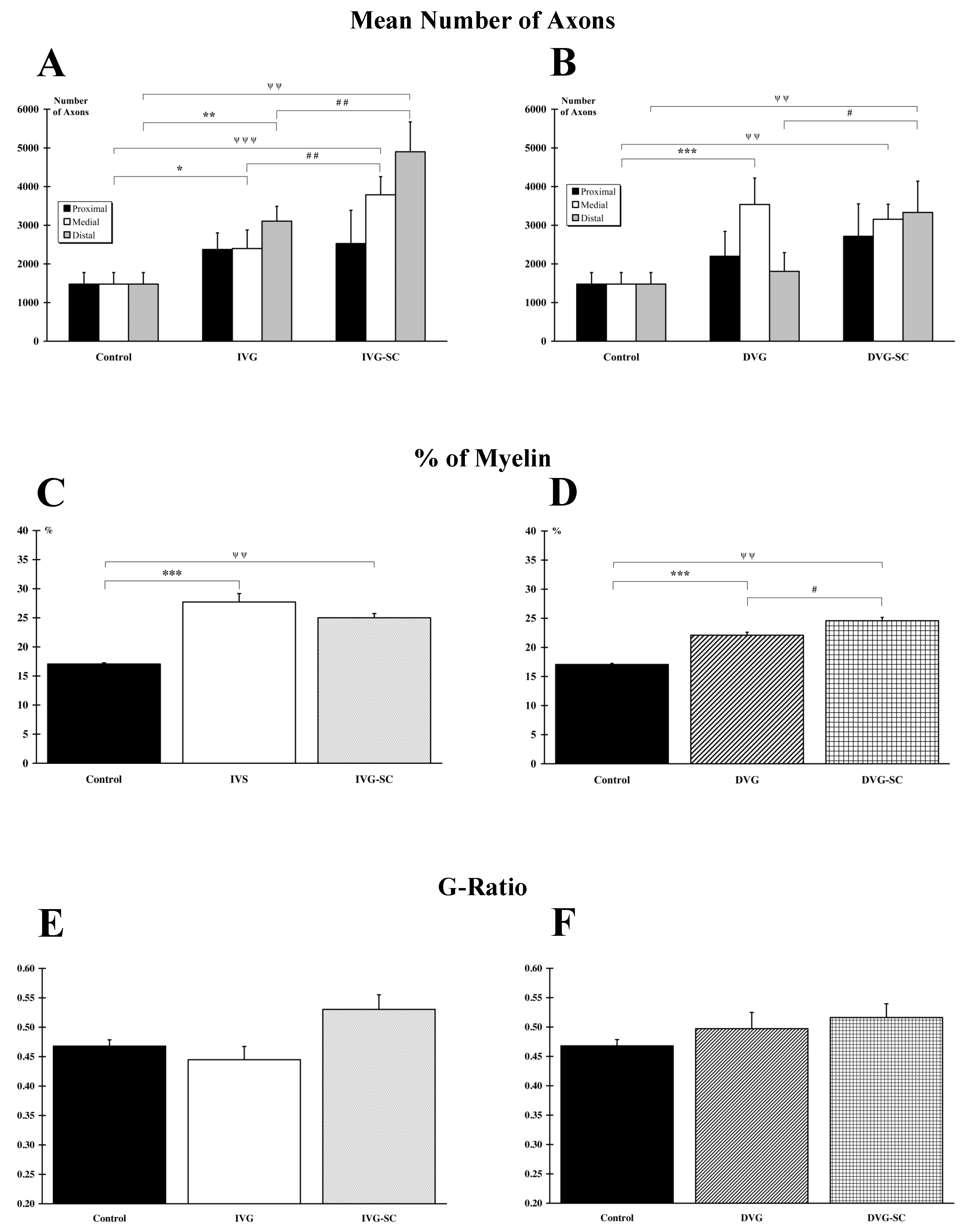
2.4. OEMSCs Increase the Number of Axons in the Delayed Animal Model
2.5. OEMSCs Increase the Total Amount of Myelin but Not the G Ratio
2.6. OEMSCs Do Not Migrate Outside the Implanted Tissue
3. Discussion
3.1. The Vein, a Suitable Biomaterial for Nerve Regeneration
3.2. Muscle Mass Is Not Correlated to Function
3.3. Olfactory Stem Cells, a Positive Add-On for Neglected Wounds
3.4. Clinical Applications
4. Materials and Methods
4.1. Animals
4.2. Experimental Groups and Surgeries
4.3. Surgery Procedure
4.4. Cell Culture
4.5. Functional Assessment of Hind Limb Recovery
4.6. Electrophysiological Recordings
4.6.1. Twitch Measurement
4.6.2. Ventilatory Response
4.6.3. Afferent Activity
4.7. Muscular Atrophy Measurement
4.8. RNA Purification and Quantitative Real-Time PCR
4.9. Histology, Immunocytochemistry and Microscopy
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Noble, J.; Munro, C.A.; Prasad, V.S.S.V.; Midha, R. Analysis of upper and lower extremity peripheral nerve injuries in a population of patients with multiple injuries. J. Trauma Injury Infect. Crit. Care 1998, 45, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Kouyoumdjian, J.A.; Graça, C.R.; Ferreira, V.F.M. Peripheral Nerve Injuries: A Retrospective Survey of 1124 Cases. Neurol. India 2017, 65, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Malahias, M.; Hindocha; Khan, W. Peripheral Nerve Injury: Principles for Repair and Regeneration. Open Orthop. J. 2014, 8, 199–203. [Google Scholar]
- Wrede, L. Uberbrueckungeinesnervendefektesmittelsseidennaht und leben venenstueckes. Dtsch. Med. Wochenschr. 1909, 35, 1125. [Google Scholar]
- Chiu, D.; Janecka, I.; Krizek, T.; Wolff, M.; Lovelace, R. Autogenous vein graft as a conduit for nerve regeneration. Surgery 1982, 91, 226–233. [Google Scholar]
- Sabongi, R.G.; Fernandes, M.; Dos Gomes Santos, J.B. Peripheral nerve regeneration with conduits: Use of vein tubes. Neural Regen. Res. 2015, 10, 529–533. [Google Scholar] [CrossRef]
- Lee, Y.; Shieh, S. Secondary nerve reconstruction using vein conduit grafts for neglected digital nerve injuries. Microsurgery 2009, 2, 504–506. [Google Scholar] [CrossRef]
- Leuzzi, S.; Armenio, A.; Leone, L.; De Santis, V.; Di Turi, A.; Annoscia, P.; Bufano, L.; Pascone, M. Repair of peripheral nerve with vein wrapping. G. Chir. 2014, 35, 101–106. [Google Scholar]
- Ahmad, I.; Akhtar, M.S. Use of Vein Conduit and Isolated Nerve Graft in Peripheral Nerve Repair: A Comparative Study. Plast. Surg. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Benito-Ruiz, J.; Navarro-Monzonis, A.; Piqueras, A.; Baena-Montilla, P. Invaginated vein graft as nerve conduit: An experimental study. Microsurgery 1994, 15, 105–115. [Google Scholar] [CrossRef]
- Bueno, C.R.d.S.; Pereira, M.; Favaretto-Júnior, I.A.; Buchaim, R.L.; Andreo, J.C.; Rodrigues, A.d.C.; Rosa-Júnior, G.M. Comparative study between standard and inside-out vein graft techniques on sciatic nerve repair of rats. Muscular and functional analysis. Acta Cir. Bras. 2017, 32, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çapkın, S.; Akhisaroğlu, M.; Ergür, B.U.; Bacakoğlu, A.A. Periferik sinir defektlerinin onarımında biyolojikt üp kullanımı ‘sinir dolması’. Ulusal Travmave Acil Cerrahi Dergisi 2017, 23, 7–14. [Google Scholar]
- Grinsell, D.; Keating, C.P. Peripheral Nerve Reconstruction after Injury: A Review of Clinical and Experimental Therapies. BioMed Res. Int. 2014, 2014, 698256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battiston, B.; Tos, P.; Conforti, L.G.; Geuna, S. Alternative techniques for peripheral nerve repair: Conduits and end-to-side neurorrhaphy. Acta Neurochir. Suppl. 2007, 100, 43–50. [Google Scholar]
- Arenas, E.; Denham, M.; Villaescusa, J.C. How to make a midbrain dopaminergic neuron. Development (Cambridge) 2015, 142, 1918–1936. [Google Scholar] [CrossRef] [Green Version]
- Monti Graziadei, G.A.; Graziadei, P.P.C. Neurogenesis and neuron regeneration in the olfactory system of mammals. II. Degeneration and reconstitution of the olfactory sensory neurons after axotomy. J. Neurocytol. 1979, 8, 197–213. [Google Scholar] [CrossRef]
- Murrell, W.; Féron, F.; Wetzig, A.; Cameron, N.; Splatt, K.; Bellette, B.; Bianco, J.; Perry, C.; Lee, G.; Mackay-Sim, A. Multipotent Stem Cells From Adult Olfactory Mucosa. Dev. Dyn. 2005, 233, 496–515. [Google Scholar] [CrossRef]
- Roisen, F.J.; Klueber, K.M.; Lu, C.L.; Hatcher, L.M.; Dozier, A.; Shields, C.B.; Maguire, S. Adult human olfactory stem cells. Brain Res. 2001, 890, 11–22. [Google Scholar] [CrossRef]
- Delorme, B.; Nivet, E.; Gaillard, J.; Häupl, T.; Ringe, J.; Devèze, A.; Magnan, J.; Sohier, J.; Khrestchatisky, M.; Roman, F.S.; et al. The human nose harbors a niche of olfactory ectomesenchymal stem cells displaying neurogenic and osteogenic properties. Stem Cells Dev. 2010, 19, 853–866. [Google Scholar] [CrossRef]
- Radtke, C.; Kocsis, J.D. Peripheral nerve injuries and transplantation of olfactory ensheathing cells for axonal regeneration and remyelination: Fact or fiction? Int. J. Mol. Sci. 2012, 13, 12911–12924. [Google Scholar] [CrossRef] [Green Version]
- Di Trapani, M.; Bassi, G.; Ricciardi, M.; Fontana, E.; Bifari, F.; Pacelli, L.; Giacomello, L.; Pozzobon, M.; Féron, F.; De Coppi, P.; et al. Comparative study of immune regulatory properties of stem cells derived from different tissues. Stem Cells Dev. 2013, 22, 2990–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, S.D.; Devéze, A.; Nivet, E.; Gepner, B.; Roman, F.S.; Féron, F. Isolating Nasal Olfactory Stem Cells from Rodents or Humans. J. Vis. Exp. 2011, 22, 27621–27625. [Google Scholar] [CrossRef] [PubMed]
- Girard, S.D.; Virard, I.; Lacassagne, E.; Paumier, J.M.; Lahlou, H.; Jabes, F.; Molino, Y.; Stephan, D.; Baranger, K.; Belghazi, M.; et al. From Blood to Lesioned Brain: An in Vitro Study on Migration Mechanisms of Human Nasal Olfactory Stem Cells. Stem Cells Int. 2017, 2017, 1478606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nivet, E.; Vignes, M.; Girard, S.D.; Pierrisnard, C.; Baril, N.; Devèze, A.; Magnan, J.; Lanté, F.; Khrestchatisky, M.; Féron, F.; et al. Engraftment of human nasal olfactory stem cells restores neuroplasticity in mice with hippocampal lesions. J. Clin. Investig. 2011, 121, 2808–2820. [Google Scholar] [CrossRef]
- Xiao, M.; Klueber, K.M.; Lu, C.; Guo, Z.; Marshall, C.T.; Wang, H.; Roisen, F.J. Human adult olfactory neural progenitors rescue axotomized rodent rubrospinal neurons and promote functional recovery. Exp. Neurol. 2005, 194, 12–30. [Google Scholar] [CrossRef]
- Xiao, M.; Klueber, K.M.; Zhou, J.P.; Guo, Z.; Lu, C.; Wang, H.; Roisen, F.J. Human adult olfactory neural progenitors promote axotomized rubrospinal tract axonal reinnervation and locomotor recovery. Neurobiol. Dis. 2007, 26, 363–374. [Google Scholar] [CrossRef]
- Pandit, S.R.; Sullivan, J.M.; Egger, V.; Borecki, A.A.; Oleskevich, S. Functional effects of adult human olfactory stem cells on early-onset sensorineural hearing loss. Stem Cells 2011, 29, 670–677. [Google Scholar] [CrossRef]
- Chao, X.; Luo, J.; Fan, Z.; Shi, H.; Han, Y.; Wang, R.; Song, Y.; Wang, G.; Wang, H.; Xu, L. Usefulness of radiological findings for predicting cochlear implantation outcomes in children with cochlear nerve deficiency: A pilot study. Acta Oto-Laryngol. 2016, 136, 1051–1057. [Google Scholar] [CrossRef]
- Roche, P.; Alekseeva, T.; Widaa, A.; Ryan, A.; Matsiko, A.; Walsh, M.; Duffy, G.P.; O’Brien, F.J. Olfactory Derived Stem Cells Delivered in a Biphasic Conduit Promote Peripheral Nerve Repair In Vivo. Stem Cells Transl. Med. 2017, 6, 1894–1904. [Google Scholar] [CrossRef]
- Lavasani, M.; Gehrmann, S.; Gharaibeh, B.; Clark, K.A.; Kaufmann, R.A.; Péault, B.; Goitz, R.J.; Huard, J. Venous graft-derived cells participate in peripheral nerve regeneration. PLoS ONE 2011, 6, e24801. [Google Scholar] [CrossRef] [Green Version]
- Demir, A.; Simsek, T.; Acar, M.; Aktaś, A.; Vlamings, R.; Ayyldz, M.; Yldrm, M.; Temel, Y.; Kaplan, S. Comparison between flexible collagen and vein conduits used for size-discrepant nerve repair: An experimental study in rats. J. Reconstr. Microsurg. 2014, 30, 329–334. [Google Scholar] [PubMed]
- Özkan, H.S.; Silistreli, Ö.K.; Ergür, B.; İrkören, S. Yağdokusundan elde edilmiş stromal vasküler fraksiyonla doldurulmuş ven greftleri ile periferik sinir kayıplarının onarımı: Deneysel çalışma. Ulusal Travmave Acil Cerrahi Dergisi 2016, 22, 7–11. [Google Scholar]
- Walton, R.L.; Brown, R.E.; Matory, W.E.; Borah, G.L.; Dolph, J.L. Autogenous vein graft repair of digital nerve defects in the finger: A retrospective clinical study. Plast. Reconstr. Surg. 1989, 84, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.B.; Gu, Y.Q.; Song, Y.S. Repair of digital nerve defect with autogenous vein graft during flexor tendon surgery in zone 2. J. Hand Surg. 1993, 18, 449–453. [Google Scholar] [CrossRef]
- Calcagnotto, G.N.; Braga Silva, J. La réparation de pertes de substance des nerves digitaux avec la technique de la greffeveineuse plus interposition de tissunerveux. Étude prospective et randomisée. Chir. Main 2006, 25, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Tenny, J.R.; Lewis, R.C. Digital Nerve-Grafting for Traumatic defects. J. Bone Joint Surg. 1984, 66, 1375–1379. [Google Scholar] [CrossRef]
- Weinzweig, N.; Gonzale, G.C.; Mead, M.; Stone, A.; Nagle, D.; Mark, G.; Koerber, A. Recovery of Sensibility After Digital Neurorrhaphy: A Clinical Investigation of Prognostic Factors. Ann. Plast. Surg. 2000, 44, 610–617. [Google Scholar] [CrossRef]
- Ronchi, G.; Morano, M.; Fregnan, F.; Pugliese, P.; Crosio, A.; Tos, P.; Geuna, S.; Haastert-Talini, K.; Gambarotta, G. The median nerve injury model in pre-clinical research—A critical review on benefits and limitations. Front. Cell. Neurosci. 2019, 13, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, J.; Mackinnon, S.E.; Watanabe, O.; Ball, D.J.; Gu, X.M.; Hunter, D.A.; Kuzon, W.M. The effect of duration of muscle denervation on functional recovery in the rat model. Muscle Nerve 1997, 20, 858–866. [Google Scholar] [CrossRef]
- Aydin, M.A.; Mackinnon, S.E.; Gu, X.M.; Kobayashi, J.; Kuzon, W.M. Force deficits in skeletal muscle after delayed reinnervation. Plast. Reconstr. Surg. 2004, 113, 1712–1718. [Google Scholar] [CrossRef]
- Marqueste, T.; Decherchi, P.; Dousset, E.; Berthelin, F.; Jammes, Y. Effect of muscle electrostimulation on afferent activities from tibialis anterior muscle after nerve repair by self-anastomosis. Neuroscience 2002, 113, 257–271. [Google Scholar] [CrossRef]
- Gordon, T.; Zepetnek, J.E. TMotor unit and muscle fiber type grouping after peripheral nerve injury in the rat. Exp. Neurol. 2016, 285, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Bense, F.; Montava, M.; Duclos, C.; Bisson, A.; Delarue, Q.; Bon Mardion, N.; Boyer, O.; Feron, F.; Guérout, N.; Marie, J.-P. Syngeneic transplantation of rat olfactory stem cells improves facial movements and reduces synkinesis after nerve injury. Plast. Reconst. Surg. 2020, in press. [Google Scholar]
- Gu, J.; Xu, H.; Xu, Y.P.; Liu, H.H.; Lang, J.T.; Chen, X.P.; Xu, W.H.; Deng, Y.; Fan, J.P. Olfactory ensheathing cells promote nerve regeneration and functional recovery after facial nerve defects. Neural Regen. Res. 2019, 14, 124–131. [Google Scholar] [PubMed]
- Ge, X.; Tu, S.; Mao, G.; Wang, C.X.; Han, T. 5G Ultra-Dense Cellular Networks. IEEE Wirel. Commun. 2016, 23, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Rui, K.; Zhang, Z.; Tian, J.; Lin, X.; Wang, X.; Ma, J.; Tang, X.; Xu, H.; Lu, L.; Wang, S. Olfactory ecto-mesenchymal stem cells possess immunoregulatory function and suppress autoimmune arthritis. Cell. Mol. Immunol. 2016, 13, 401–408. [Google Scholar] [CrossRef] [Green Version]
- MacKinnon, S.E.; Dellon, A.L.; O’Brien, J.P. Changes in nerve fiber numbers distal to a nerve repair in the rat sciatic nerve model. Muscle Nerve 1991, 14, 1116–1122. [Google Scholar] [CrossRef]
- Kushnerev, E.; Yates, J.M. Evidence-based outcomes following inferior alveolar and lingual nerve injury and repair: A systematic review. J. Oral Rehabil. 2015, 42, 786–802. [Google Scholar] [CrossRef]
- Yawn, R.J.; Wright, H.V.; Francis, D.O.; Stephan, S.; Bennett, M.L. Facial nerve repair after operative injury: Impact of timing on hypoglossal-facial nerve graft outcomes. Am. J. Otolaryngol. Head Neck Med. Surg. 2016, 37, 493–496. [Google Scholar] [CrossRef]
- Harrell, C.R.; Fellabaum, C.; Arsenijevic, A.; Markovic, B.S.; Djonov, V.; Volarevic, V. Therapeutic potential of mesenchymal stem cells and their secretome in the treatment of glaucoma. Stem Cells Int. 2019, 27, 7869130. [Google Scholar] [CrossRef]
- Xia, B.; Gao, J.; Li, S.; Huang, L.; Ma, T.; Zhao, L.; Yang, Y.; Huang, J.; Luo, Z. Extracellular Vesicles Derived From Olfactory Ensheathing Cells Promote Peripheral Nerve Regeneration in Rats. Front. Cell. Neurosci. 2019, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Féron, F.; Perry, C.; Girard, S.D.; Mackay-Sim, A. Isolation of Adult Stem Cells from the Human Olfactory Mucosa. Neural Progenit. Cells 2013, 107–114. [Google Scholar] [CrossRef]
- Chabas, J.; Alluin, O.; Rao, G.; Garcia, S.; Lavaut, M.; Risso, J.J.; Legre, R.; Magalon, G.; Khrestchatisky, M.; Marqueste, T.; et al. Vitamin D 2 Potentiates Axon Regeneration. J. Neurotrauma 2008, 1256, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.R.; Mackinnon, S.E.; Hunter, D.A. Functional Evaluation of Complete Sciatic, Peroneal, and Posterior Tibial Nerve Lesions in the Rat. Plast. Reconstr. Surg. 1989, 83, 129–136. [Google Scholar] [CrossRef]
- Decherchi, P.; Dousset, E.; Jammes, Y. Respiratory and cardiovascular responses evoked by tibialis anterior muscle a V erent W bers in rats. Exp. Brain Res. 2007, 183, 299–312. [Google Scholar] [CrossRef]
- Darques, J.L.; Jammes, Y. Fatigue-induced changes in group IV muscle afferent activity: Differences between high- and low-frequency electrically induced fatigues. Brain Res. 1997, 750, 147–154. [Google Scholar] [CrossRef]
- Chabas, J.F.; Stephan, D.; Marqueste, T.; Garcia, S.; Lavaut, M.N.; Nguyen, C.; Legre, R.; Khrestchatisky, M.; Decherchi, P.; Feron, F. Cholecalciferol (Vitamin D3) Improves Myelination and Recovery after Nerve Injury. PLoS ONE 2013, 8, e65034. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnet, M.; Guiraudie-Capraz, G.; Marqueste, T.; Garcia, S.; Jaloux, C.; Decherchi, P.; Féron, F. Immediate or Delayed Transplantation of a Vein Conduit Filled with Nasal Olfactory Stem Cells Improves Locomotion and Axogenesis in Rats after a Peroneal Nerve Loss of Substance. Int. J. Mol. Sci. 2020, 21, 2670. https://doi.org/10.3390/ijms21082670
Bonnet M, Guiraudie-Capraz G, Marqueste T, Garcia S, Jaloux C, Decherchi P, Féron F. Immediate or Delayed Transplantation of a Vein Conduit Filled with Nasal Olfactory Stem Cells Improves Locomotion and Axogenesis in Rats after a Peroneal Nerve Loss of Substance. International Journal of Molecular Sciences. 2020; 21(8):2670. https://doi.org/10.3390/ijms21082670
Chicago/Turabian StyleBonnet, Maxime, Gaëlle Guiraudie-Capraz, Tanguy Marqueste, Stéphane Garcia, Charlotte Jaloux, Patrick Decherchi, and François Féron. 2020. "Immediate or Delayed Transplantation of a Vein Conduit Filled with Nasal Olfactory Stem Cells Improves Locomotion and Axogenesis in Rats after a Peroneal Nerve Loss of Substance" International Journal of Molecular Sciences 21, no. 8: 2670. https://doi.org/10.3390/ijms21082670
APA StyleBonnet, M., Guiraudie-Capraz, G., Marqueste, T., Garcia, S., Jaloux, C., Decherchi, P., & Féron, F. (2020). Immediate or Delayed Transplantation of a Vein Conduit Filled with Nasal Olfactory Stem Cells Improves Locomotion and Axogenesis in Rats after a Peroneal Nerve Loss of Substance. International Journal of Molecular Sciences, 21(8), 2670. https://doi.org/10.3390/ijms21082670