Crocetin Prevents RPE Cells from Oxidative Stress through Protection of Cellular Metabolic Function and Activation of ERK1/2

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
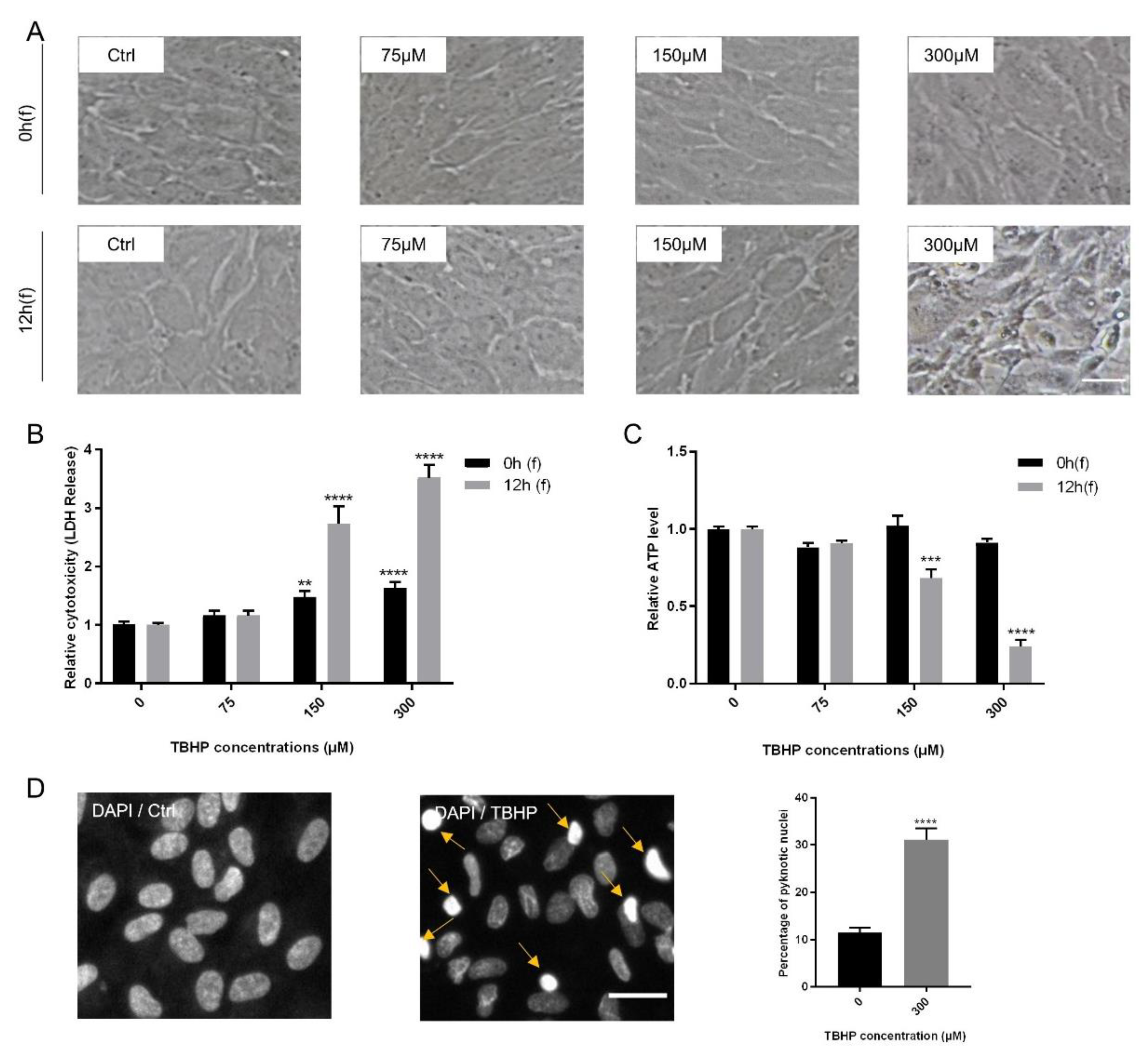
2.1. Establishment of An In Vitro Oxidative Stress Model Using Exposure of ARPE19 Cells to TBHP
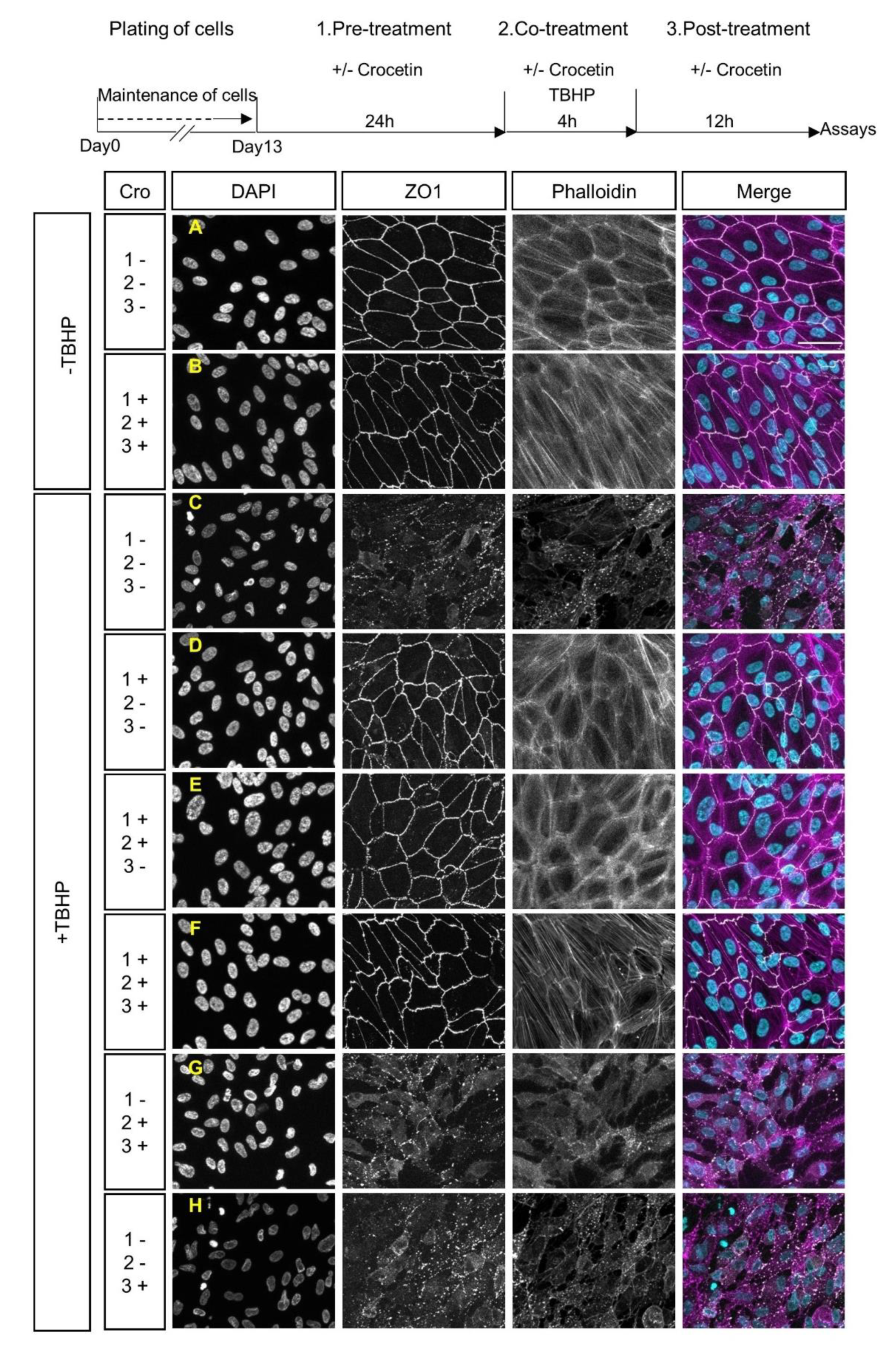
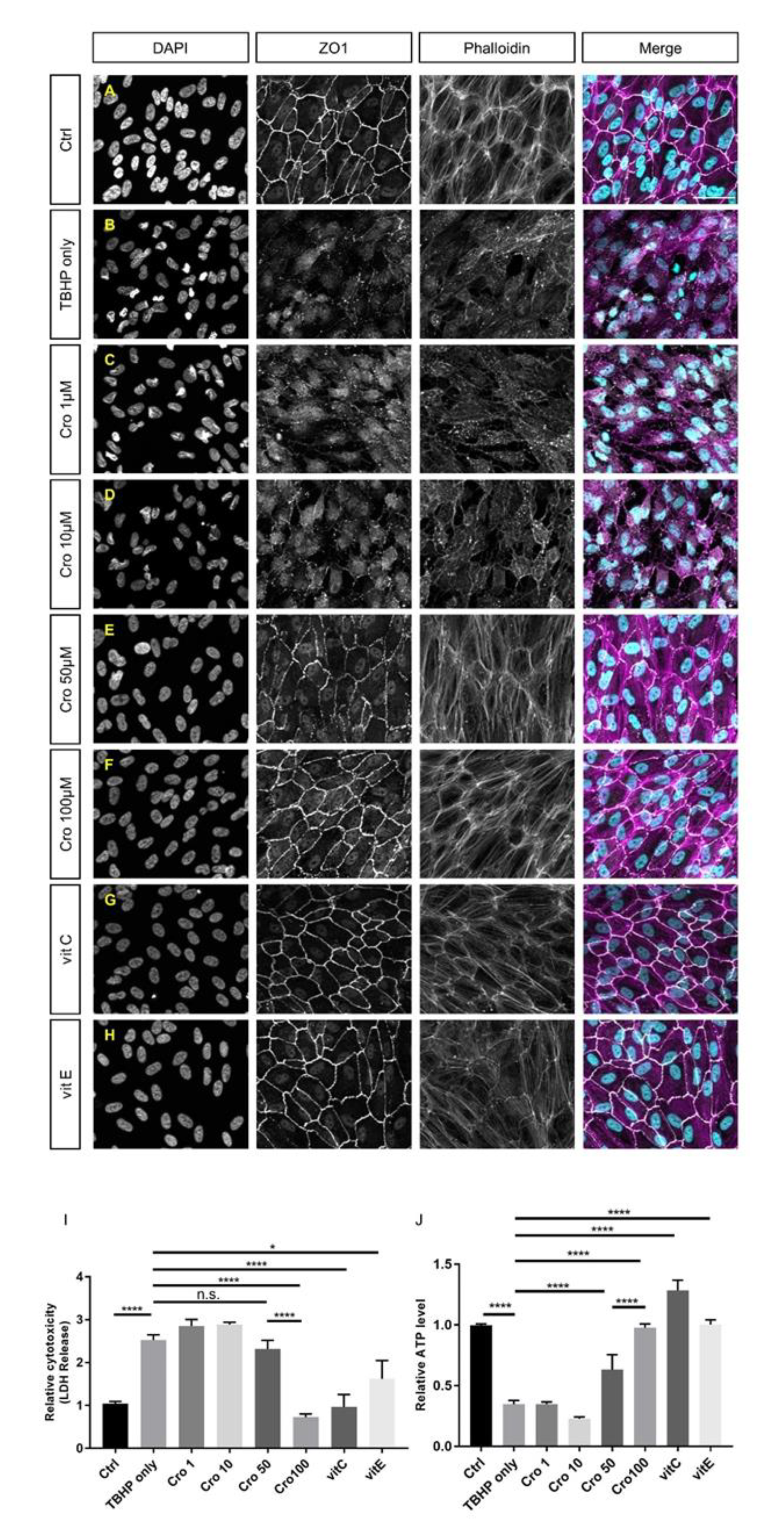
2.2. Protection of Junctional Integrity and Morphology of TBHP-Stressed ARPE19 Cells by Crocetin Treatment
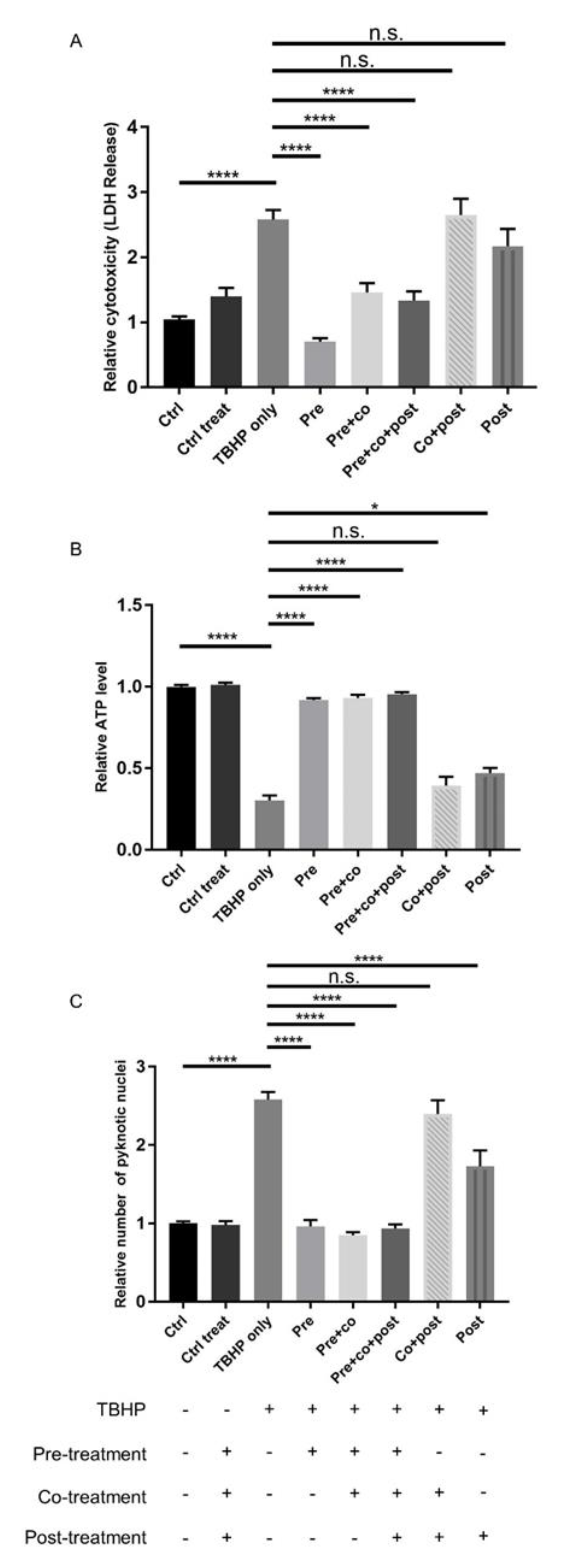
2.3. Protection of ARPE19 Cells from TBHP-induced Oxidative Stress using Crocetin Treatment
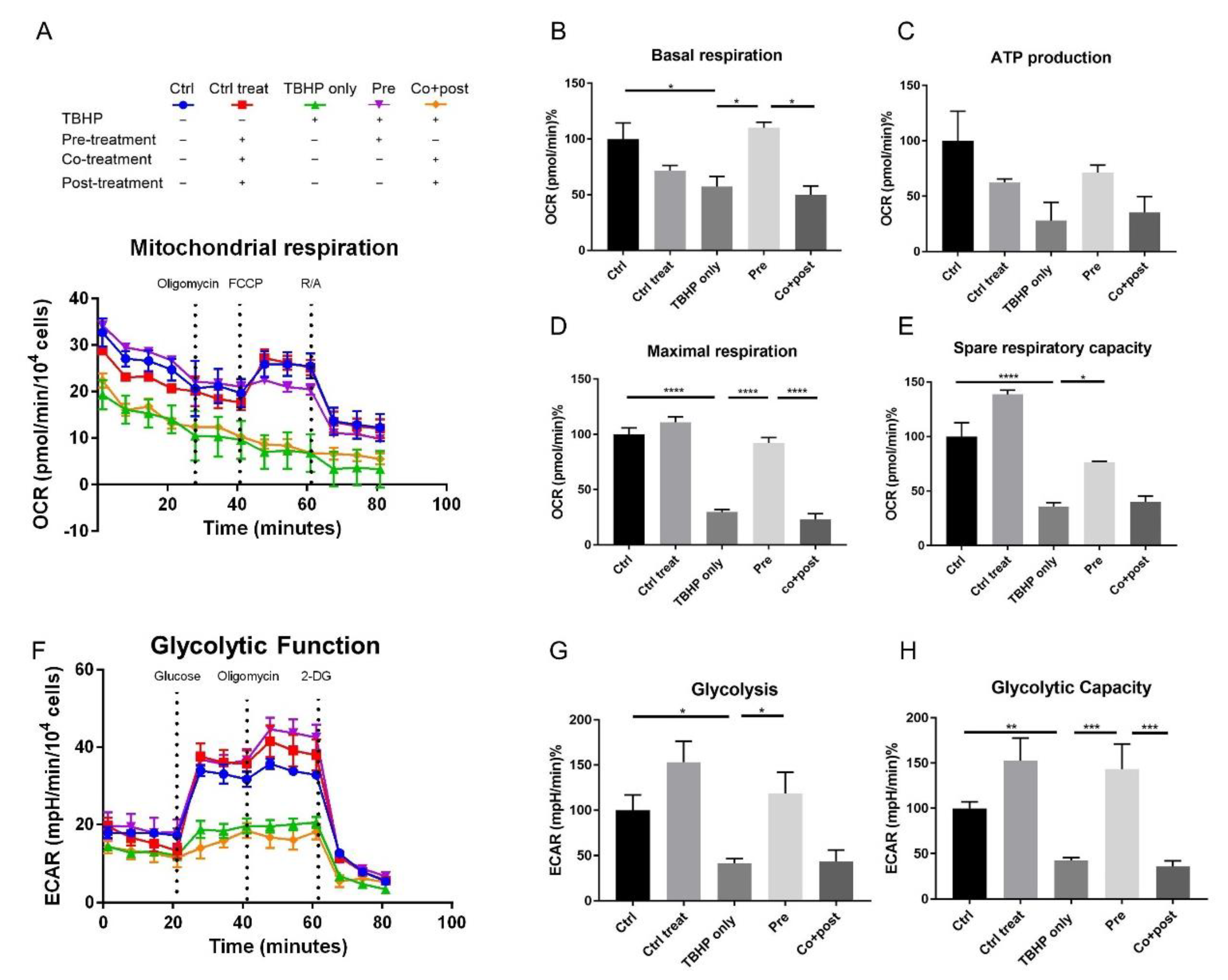
2.4. Protection of Energy Production Pathways in Stressed ARPE19 Cells by Crocetin Pre-Treatment
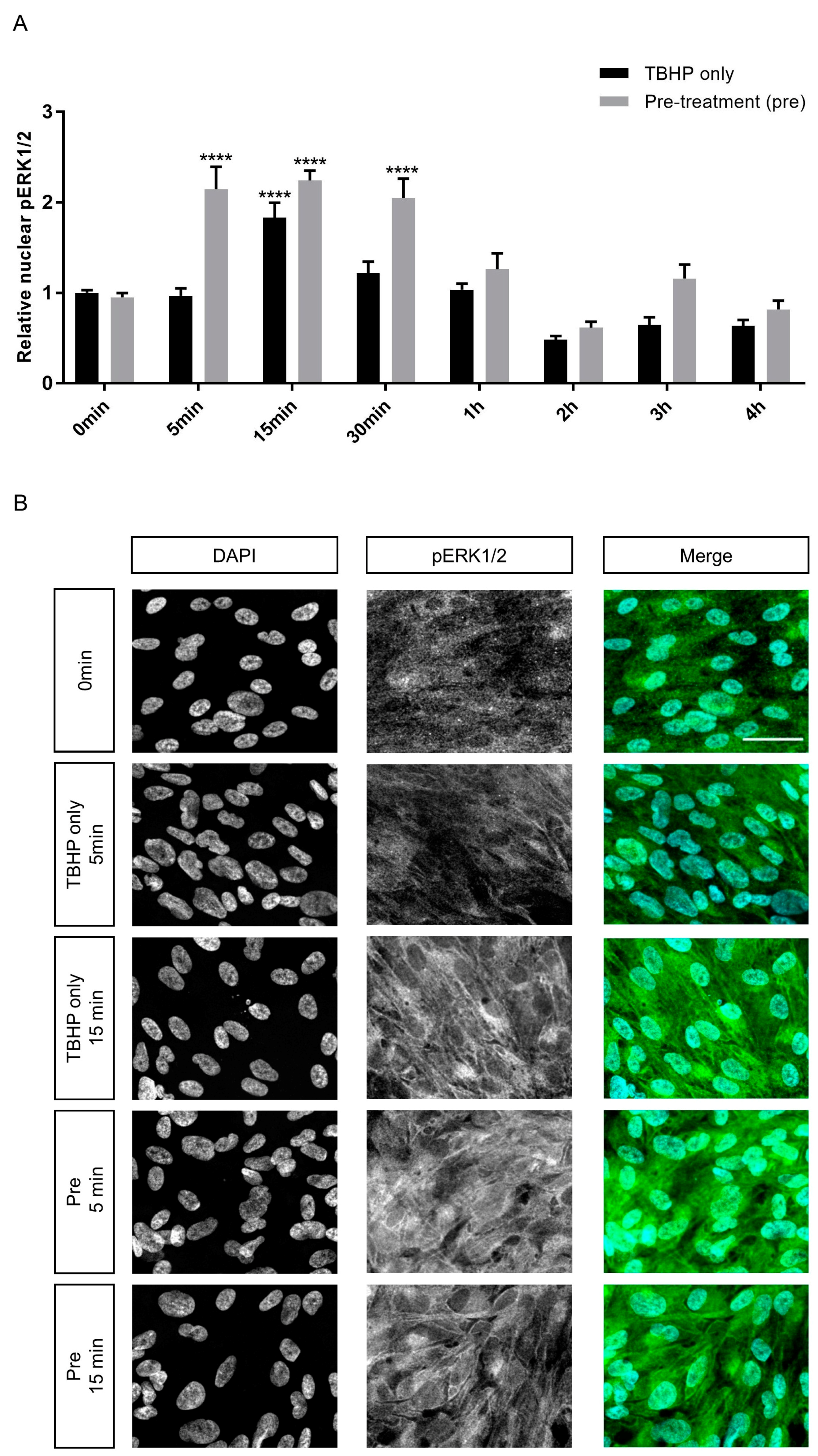
2.5. Crocetin Protects Stressed ARPE19 Cells through ERK1/2 Activation
3. Discussion
4. Materials and Methods
4.1. ARPE19 Cell Culture
4.2. Exposure of ARPE19 Cells to TBHP As An in Vitro Model of Oxidative Stress
4.3. MTS Assay
4.4. Crocetin and Vitamins Solution Preparation and Treatment
4.5. LDH Assay
4.6. ATP Assay
4.7. DAPI Staining
4.8. Immunocytochemistry
4.8.1. Detection of ZO1 and F-actin
4.8.2. MAPK Data Evaluation
4.9. Real-time Metabolic Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMD | Age related macular degeneration |
| RPE | Retinal pigment epithelium |
| ERK | Extracellular signal-regulated kinase |
| pERK | Phospho extracellular signal-regulated kinase |
| MAPK | Mitogen activated protein kinase |
| LDH | Lactate dehydrogenase |
| ATP | Adenosine triphosphate |
| ROS | Reactive oxygen species |
| VEGF | Vascular endothelial growth factor |
| GA | Geographic atrophy |
| TBHP | Tert-butyl hydroperoxide |
| DMSO | Dimethyl-sulfoxide |
| ZO1 | Zonula occludens |
| ETC | Electron transport chain |
| FCCP | Carbonyl cyanide-4 (trifluoromethoxy) phenylhydrazone |
| OCR | Oxygen consumption rate |
| ECAR | Extracellular acidification rate |
| EDTRS | Early Treatment Diabetic Retinopathy Study |
| ERG | Electroretinography |
| MtDNA | Mitochondrial DNA |
| DAPK | Death associated protein kinase |
| PDGF | Platelet derived growth factor |
| H2O2 | Hydrogen peroxide |
| GSH | Glutathione |
| JNK | c-jun N-terminal kinase |
| iPSC | Induced pluripotent stem cells |
| PBS | Phosphate-buffered saline |
| EDF | Extend Depth-of-Field |
| 2-DG | 2-deoxy glucose |
| Mito | Mitochondrial |
| CS | Contrast sensitivity |
| Pre | Pre-treatment |
| Co | Co-treatment |
| Post | Post-treatment |
| Cro | Crocetin |
References
- Kaur, I.P.; Kakkar, S. Nanotherapy for Posterior Eye Diseases. J. Control. Release 2014, 193, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Chou, R.; Dana, T.; Bougatsos, C.; Grusing, S.; Blazina, I. Screening for Impaired Visual Acuity in Older Adults. JAMA 2016, 315, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.W. Age-Related Macular Degeneration Revisited–Piecing the Puzzle: The LXIX Edward Jackson Memorial Lecture. Am. J. Ophthalmol. 2013, 155, 1–35.e13. [Google Scholar] [CrossRef] [PubMed]
- Biarnés, M.; Monés, J.; Alonso, J.; Arias, L. Update on Geographic Atrophy in Age-Related Macular Degeneration. Optom. Vis. Sci. 2011, 88, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Colijn, J.M.; Buitendijk, G.H.S.; Prokofyeva, E.; Alves, D.; Cachulo, M.L.; Khawaja, A.P.; Cougnard-Gregoire, A.; Merle, B.M.J.; Korb, C.; Erke, M.G.; et al. Prevalence of Age-Related Macular Degeneration in Europe: The Past and the Future. Ophthalmology 2017, 124, 1753–1763. [Google Scholar] [CrossRef] [Green Version]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.-Y.; Wong, T.Y. Global Prevalence of Age-Related Macular Degeneration and Disease Burden Projection for 2020 and 2040: A Systematic Review and Meta-Analysis. Lancet Glob. Heal. 2014, 2, e106–e116. [Google Scholar] [CrossRef] [Green Version]
- Coleman, H.R.; Chan, C.-C.; Ferris, F.L.; Chew, E.Y.; Chew, E.Y. Age-Related Macular Degeneration. Lancet 2008, 372, 1835–1845. [Google Scholar] [CrossRef]
- Miller, J.W. VEGF: From Discovery to Therapy: The Champalimaud Award Lecture. Transl. Vis. Sci. Technol. 2016, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Bisti, S.; Maccarone, R.; Falsini, B. Saffron and Retina: Neuroprotection and Pharmacokinetics. Vis. Neurosci. 2014, 31, 355–361. [Google Scholar] [CrossRef]
- Miller, J.W.; Bagheri, S.; Vavvas, D.G. Advances in Age-Related Macular Degeneration Understanding and Therapy. US Ophthalmic Rev. 2017, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Beatty, S.; Koh, H.-H.; Phil, M.; Henson, D.; Boulton, M. The Role of Oxidative Stress in the Pathogenesis of Age-Related Macular Degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Buschini, E.; Piras, A.; Nuzzi, R.; Vercelli, A. Age Related Macular Degeneration and Drusen: Neuroinflammation in the Retina. Prog. Neurobiol. 2011, 95, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carocho, M.; Ferreira, I.C.F.R.; Morales, P.; Soković, M. Antioxidants and Prooxidants: Effects on Health and Aging 2018. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubotav, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccà, S.C.; Cutolo, C.A.; Ferrari, D.; Corazza, P.; Traverso, C.E. The Eye, Oxidative Damage and Polyunsaturated Fatty Acids. Nutrients 2018, 10, 668. [Google Scholar] [CrossRef] [Green Version]
- Felszeghy, S.; Viiri, J.; Paterno, J.J.; Hyttinen, J.M.T.; Koskela, A.; Chen, M.; Leinonen, H.; Tanila, H.; Kivinen, N.; Koistinen, A.; et al. Loss of NRF-2 and PGC-1α Genes Leads to Retinal Pigment Epithelium Damage Resembling Dry Age-Related Macular Degeneration. Redox Biol. 2019, 20, 1–12. [Google Scholar] [CrossRef]
- Telegina, D.V.; Kozhevnikova, O.S.; Bayborodin, S.I.; Kolosova, N.G. Contributions of Age-Related Alterations of the Retinal Pigment Epithelium and of Glia to the AMD-like Pathology in OXYS Rats. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Bellezza, I. Oxidative Stress in Age-Related Macular Degeneration: NRF2 as Therapeutic Target. Front. Pharmacol. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Rohowetz, L.J.; Kraus, J.G.; Koulen, P. Reactive Oxygen Species-Mediated Damage of Retinal Neurons: Drug Development Targets for Therapies of Chronic Neurodegeneration of the Retina. Int. J. Mol. Sci. 2018, 19, 3362. [Google Scholar] [CrossRef] [Green Version]
- Rivera, J.C.; Dabouz, R.; Noueihed, B.; Omri, S.; Tahiri, H.; Chemtob, S. Ischemic Retinopathies: Oxidative Stress and Inflammation. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Resnikoff, S.; Pascolini, D.; Etya’ale, D.; Kocur, I.; Pararajasegaram, R.; Pokharel, G.P.; Mariotti, S.P. Global Data on Visual Impairment in the Year 2002. Bull. World Health Organ. 2004, 82, 844–851. [Google Scholar] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The Impact of Oxidative Stress and Inflammation on RPE Degeneration in Non-Neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Garg, T.K.; Chang, J.Y. Oxidative Stress Causes ERK Phosphorylation and Cell Death in Cultured Retinal Pigment Epithelium: Prevention of Cell Death by AG126 and 15-Deoxy-Delta 12, 14-PGJ2. BMC Ophthalmol. 2003, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Maidana, D.E.; Dib, B.; Miller, J.B.; Bouzika, P.; Miller, J.W.; Vavvas, D.G.; Lin, H. MiR-17-3p Exacerbates Oxidative Damage in Human Retinal Pigment Epithelial Cells. PLoS ONE 2016, 11, e0160887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.-Q.; Godley, B.F. Oxidative Stress-Induced Mitochondrial DNA Damage in Human Retinal Pigment Epithelial Cells: A Possible Mechanism for RPE Aging and Age-Related Macular Degeneration. Exp. Eye Res. 2003, 76, 397–403. [Google Scholar] [CrossRef]
- Chong, C.-M.; Zheng, W. Artemisinin Protects Human Retinal Pigment Epithelial Cells from Hydrogen Peroxide-Induced Oxidative Damage through Activation of ERK/CREB Signaling. Redox Biol. 2016, 9, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanneken, A.; Lin, F.F.; Johnson, J.; Maher, P. Flavonoids Protect Human Retinal Pigment Epithelial Cells from Oxidative-Stress-Induced Death. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3164–3177. [Google Scholar] [CrossRef]
- Cai, H.; Gong, J.; Abriola, L.; Hoyer, D.; Noggle, S.; Paull, D.; Del Priore, L.V.; Fields, M.A.; NYSCF Global Stem Cell Array Team. High-Throughput Screening Identifies Compounds That Protect RPE Cells from Physiological Stressors Present in AMD. Exp. Eye Res. 2019, 185. [Google Scholar] [CrossRef]
- The Age-Related Eye Disease Study System for Classifying Age-Related Macular Degeneration from Stereoscopic Color Fundus Photographs: The Age-Related Eye Disease Study Report Number 6. Am. J. Ophthalmol. 2001, 132, 668–681. [CrossRef]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The Effects of Plant Flavonoids on Mammalian Cells: Implications for Inflammation, Heart Disease, and Cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Johnsen-Soriano, S.; Garcia-Pous, M.; Arnal, E.; Sancho-Tello, M.; Garcia-Delpech, S.; Miranda, M.; Bosch-Morell, F.; Diaz-Llopis, M.; Navea, A.; Javier Romero, F. Early Lipoic Acid Intake Protects Retina of Diabetic Mice. Free Radic. Res. 2008, 42, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Hanus, J.; Kolkin, A.; Chimienti, J.; Botsay, S.; Wang, S. 4-Acetoxyphenol Prevents RPE Oxidative Stress–Induced Necrosis by Functioning as an NRF2 Stabilizer. Investig. Opthalmology Vis. Sci. 2015, 56, 5048. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.M.; Shin, D.-Y.; Lee, S.J.; Joe, Y.; Zheng, M.; Yim, J.H.; Callaway, Z.; Chung, H.T. Curcumin Protects Retinal Pigment Epithelial Cells against Oxidative Stress via Induction of Heme Oxygenase-1 Expression and Reduction of Reactive Oxygen. Mol. Vis. 2012, 18, 901–908. [Google Scholar] [PubMed]
- Silván, J.M.; Reguero, M.; de Pascual-Teresa, S. A Protective Effect of Anthocyanins and Xanthophylls on UVB-Induced Damage in Retinal Pigment Epithelial Cells. Food Funct. 2016, 7, 1067–1076. [Google Scholar] [CrossRef]
- Bucolo, C.; Drago, F.; Lin, L.-R.; Reddy, V.N. Sigma Receptor Ligands Protect Human Retinal Cells against Oxidative Stress. Neuroreport 2006, 17, 287–291. [Google Scholar] [CrossRef]
- Ochiai, T.; Shimeno, H.; Mishima, K.; Iwasaki, K.; Fujiwara, M.; Tanaka, H.; Shoyama, Y.; Toda, A.; Eyanagi, R.; Soeda, S. Protective Effects of Carotenoids from Saffron on Neuronal Injury in Vitro and in Vivo. Biochim. Biophys. Acta-Gen. Subj. 2007, 1770, 578–584. [Google Scholar] [CrossRef]
- Corso, L.; Cavallero, A.; Baroni, D.; Garbati, P.; Prestipino, G.; Bisti, S.; Nobile, M.; Picco, C. Saffron Reduces ATP-Induced Retinal Cytotoxicity by Targeting P2X7 Receptors. Purinergic Signal. 2016, 12, 161–174. [Google Scholar] [CrossRef] [Green Version]
- Maccarone, R.; Di Marco, S.; Bisti, S. Saffron Supplement Maintains Morphology and Function after Exposure to Damaging Light in Mammalian Retina. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1254–1261. [Google Scholar] [CrossRef] [Green Version]
- Jabbarpoor Bonyadi, M.H.; Yazdani, S.; Saadat, S. The Ocular Hypotensive Effect of Saffron Extract in Primary Open Angle Glaucoma: A Pilot Study. BMC Complement. Altern. Med. 2014, 14, 399. [Google Scholar] [CrossRef] [Green Version]
- Sepahi, S.; Mohajeri, S.A.; Hosseini, S.M.; Khodaverdi, E.; Shoeibi, N.; Namdari, M.; Tabassi, S.A.S. Effects of Crocin on Diabetic Maculopathy: A Placebo-Controlled Randomized Clinical Trial. Am. J. Ophthalmol. 2018, 190, 89–98. [Google Scholar] [CrossRef]
- Falsini, B.; Piccardi, M.; Minnella, A.; Savastano, C.; Capoluongo, E.; Fadda, A.; Balestrazzi, E.; Maccarone, R.; Bisti, S. Influence of Saffron Supplementation on Retinal Flicker Sensitivity in Early Age-Related Macular Degeneration. Investig. Opthalmology Vis. Sci. 2010, 51, 6118. [Google Scholar] [CrossRef]
- Piccardi, M.; Marangoni, D.; Minnella, A.M.; Savastano, M.C.; Valentini, P.; Ambrosio, L.; Capoluongo, E.; MacCarone, R.; Bisti, S.; Falsini, B. A Longitudinal Follow-up Study of Saffron Supplementation in Early Age-Related Macular Degeneration: Sustained Benefits to Central Retinal Function. Evid.-Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef]
- Marangoni, D.; Falsini, B.; Piccardi, M.; Ambrosio, L.; Minnella, A.; Savastano, M.; Bisti, S.; Maccarone, R.; Fadda, A.; Mello, E.; et al. Functional Effect of Saffron Supplementation and Risk Genotypes in Early Age-Related Macular Degeneration: A Preliminary Report. J. Transl. Med. 2013, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Lashay, A.; Sadough, G.; Ashrafi, E.; Lashay, M.; Movassat, M.; Akhondzadeh, S. Short-Term Outcomes of Saffron Supplementation in Patients with Age-Related Macular Degeneration: A Double-Blind, Placebo-Controlled, Randomized Trial. Med. Hypothesis Discov. Innov. Ophthalmol. J. 2016, 5, 32–38. [Google Scholar]
- Riazi, A.; Panahi, Y.; Alishiri, A.A.; Hosseini, M.A.; Karimi Zarchi, A.A.; Sahebkar, A. The Impact of Saffron (Crocus Sativus) Supplementation on Visual Function in Patients with Dry Age-Related Macular Degeneration. Ital. J. Med. 2016, 10, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Broadhead, G.K.; Grigg, J.R.; McCluskey, P.; Hong, T.; Schlub, T.E.; Chang, A.A. Saffron Therapy for the Treatment of Mild/Moderate Age-Related Macular Degeneration: A Randomised Clinical Trial. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Li, K.; Liu, X.; Zhong, J.; Xiang, Y. Crocin Protects Retinal Pigment Epithelial Cells from Oxidative Stress through Suppression of the MAPK Signaling Pathway. Int. J. Clin. Exp. Med. 2016, 9, 11016–11022. [Google Scholar]
- Asai, A.; Nakano, T.; Takahashi, M.; Nagao, A. Orally Administered Crocetin and Crocins Are Absorbed into Blood Plasma as Crocetin and Its Glucuronide Conjugates in Mice. J. Agric. Food Chem. 2005, 53, 7302–7306. [Google Scholar] [CrossRef]
- Yamauchi, M.; Tsuruma, K.; Imai, S.; Nakanishi, T.; Umigai, N.; Shimazawa, M.; Hara, H. Crocetin Prevents Retinal Degeneration Induced by Oxidative and Endoplasmic Reticulum Stresses via Inhibition of Caspase Activity. Eur. J. Pharmacol. 2011, 650, 110–119. [Google Scholar] [CrossRef]
- GIACCIO, M. Crocetin from Saffron: An Active Component of an Ancient Spice. Crit. Rev. Food Sci. Nutr. 2004, 44, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, F.; Yoshida, A.; Umigai, N.; Kubo, K.; Lee, M.-C. Crocetin Reduces the Oxidative Stress Induced Reactive Oxygen Species in the Stroke-Prone Spontaneously Hypertensive Rats (SHRSPs) Brain. J. Clin. Biochem. Nutr. 2011, 49, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.S.; Ansari, M.A.; Ahmad, M.; Saleem, S.; Yousuf, S.; Hoda, M.N.; Islam, F. Neuroprotection by Crocetin in a Hemi-Parkinsonian Rat Model. Pharmacol. Biochem. Behav. 2005, 81, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kakkar, R.; Wang, J. In Vivo and in Vitro Approach to Anti-Arthritic and Anti-Inflammatory Effect of Crocetin by Alteration of Nuclear Factor-E2-Related Factor 2/Hem Oxygenase (HO)-1 and NF-ΚB Expression. Front. Pharmacol. 2018, 9, 1341. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.; Guha, D.; Chakraborty, J.; Banerjee, S.; Adhikary, A.; Chakraborty, S.; Das, T.; Sa, G. Crocetin Exploits P53-Induced Death Domain (PIDD) and FAS-Associated Death Domain (FADD) Proteins to Induce Apoptosis in Colorectal Cancer. Sci. Rep. 2016, 6, 32979. [Google Scholar] [CrossRef]
- Cai, J.; Yi, F.F.; Bian, Z.Y.; Shen, D.F.; Yang, L.; Yan, L.; Tang, Q.Z.; Yang, X.C.; Li, H. Crocetin Protects against Cardiac Hypertrophy by Blocking MEK-ERK1-2 Signalling Pathway. J. Cell. Mol. Med. 2009, 13, 909–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.F.; Ma, J.X.; Shang, Q.L.; An, J.B.; Chen, H.T.; Wang, C.X. Safety, Pharmacokinetics, and Prevention Effect of Intraocular Crocetin in Proliferative Vitreoretinopathy. Biomed. Pharmacother. 2019, 109, 1211–1220. [Google Scholar] [CrossRef]
- Ishizuka, F.; Shimazawa, M.; Umigai, N.; Ogishima, H.; Nakamura, S.; Tsuruma, K.; Hara, H. Crocetin, a Carotenoid Derivative, Inhibits Retinal Ischemic Damage in Mice. Eur. J. Pharmacol. 2013, 703, 1–10. [Google Scholar] [CrossRef]
- Rabin, D.M.; Rabin, R.L.; Blenkinsop, T.A.; Temple, S.; Stern, J.H. Chronic Oxidative Stress Upregulates Drusen-Related Protein Expression in Adult Human RPE Stem Cell-Derived RPE Cells: A Novel Culture Model for Dry AMD. Aging 2013, 5, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Bend, J.R.; Xia, X.Y.; Chen, D.; Awaysheh, A.; Lo, A.; Rieder, M.J.; Jane Rylett, R. Attenuation of Oxidative Stress in HEK 293 Cells by the TCM Constituents Schisanhenol, Baicalein, Resveratrol or Crocetin and Two Defined Mixtures. J. Pharm. Pharm. Sci. 2015, 18, 661–682. [Google Scholar] [CrossRef] [Green Version]
- Mirabelli, F.; Salis, A.; Marinoni, V.; Finardi, G.; Bellomo, G.; Thor, H.; Orrenius, S. Menadione-Induced Bleb Formation in Hepatocytes Is Associated with the Oxidation of Thiol Groups in Actin. Arch. Biochem. Biophys. 1988, 264, 261–269. [Google Scholar] [CrossRef]
- DUNN, K.C.; AOTAKI-KEEN, A.E.; PUTKEY, F.R.; HJELMELAND, L.M. ARPE-19, A Human Retinal Pigment Epithelial Cell Line with Differentiated Properties. Exp. Eye Res. 1996, 62, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.A.; Kanuga, N.; Romero, I.A.; Greenwood, J.; Luthert, P.J.; Cheetham, M.E. Oxidative Stress Affects the Junctional Integrity of Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 675–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanus, J.; Anderson, C.; Wang, S. RPE Necroptosis in Response to Oxidative Stress and in AMD. Ageing Res. Rev. 2015, 24, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totsuka, K.; Ueta, T.; Uchida, T.; Roggia, M.F.; Nakagawa, S.; Vavvas, D.G.; Honjo, M.; Aihara, M. Oxidative Stress Induces Ferroptotic Cell Death in Retinal Pigment Epithelial Cells. Exp. Eye Res. 2019, 181, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; Gaidano, G. Messengers of Cell Death: Apoptotic Signaling in Health and Disease. Haematologica 2003, 88, 212–218. [Google Scholar] [PubMed]
- Ziegler, U.; Groscurth, P. Morphological Features of Cell Death. Physiology 2019, 19, 124–128. [Google Scholar] [CrossRef]
- Antoniel, M.; Giorgio, V.; Fogolari, F.; Glick, G.D.; Bernardi, P.; Lippe, G. The Oligomycin-Sensitivity Conferring Protein of Mitochondrial ATP Synthase: Emerging New Roles in Mitochondrial Pathophysiology. Int. J. Mol. Sci. 2014, 15, 7513–7536. [Google Scholar] [CrossRef] [Green Version]
- Devenish, R.J.; Prescott, M.; Boyle, G.M.; Nagley, P. The Oligomycin Axis of Mitochondrial ATP Synthase: OSCP and the Proton Channel. J. Bioenerg. Biomembr. 2000, 32, 507–516. [Google Scholar] [CrossRef]
- Mebratu, Y.; Tesfaigzi, Y. How ERK1/2 Activation Controls Cell Proliferation and Cell Death: Is Subcellular Localization the Answer? Cell Cycle 2009, 8, 1168–1175. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, Oxidative Stress and the Biology of Ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Nelson, K.C.; Wu, M.; Sternberg, P.; Jones, D.P. Oxidative Damage and Protection of the RPE. Prog. Retin. Eye Res. 2000, 19, 205–221. [Google Scholar] [CrossRef]
- Kijlstra, A.; Tian, Y.; Kelly, E.R.; Berendschot, T.T.J.M. Lutein: More than Just a Filter for Blue Light. Prog. Retin. Eye Res. 2012, 31, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Yan, W.; Beight, C. Lutein and Zeaxanthin Isomers Protect against Light-Induced Retinopathy via Decreasing Oxidative and Endoplasmic Reticulum Stress in BALB/CJ Mice. Nutrients 2018, 10, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aimjongjun, S.; Sutheerawattananonda, M.; Limpeanchob, N. Silk Lutein Extract and Its Combination with Vitamin E Reduce UVB-Mediated Oxidative Damage to Retinal Pigment Epithelial Cells. J. Photochem. Photobiol. B Biol. 2013, 124, 34–41. [Google Scholar] [CrossRef]
- Chan, C.-M.; Huang, J.-H.; Lin, H.-H.; Chiang, H.-S.; Chen, B.-H.; Hong, J.-Y.; Hung, C.-F. Protective Effects of (-)-Epigallocatechin Gallate on UVA-Induced Damage in ARPE19 Cells. Mol. Vis. 2008, 14, 2528–2534. [Google Scholar]
- Mookerjee, S.A.; Gerencser, A.A.; Nicholls, D.G.; Brand, M.D. Quantifying Intracellular Rates of Glycolytic and Oxidative ATP Production and Consumption Using Extracellular Flux Measurements. J. Biol. Chem. 2017, 292, 7189–7207. [Google Scholar] [CrossRef] [Green Version]
- Golestaneh, N.; Chu, Y.; Cheng, S.K.; Cao, H.; Poliakov, E.; Berinstein, D.M. Repressed SIRT1/PGC-1α Pathway and Mitochondrial Disintegration in IPSC-Derived RPE Disease Model of Age-Related Macular Degeneration. J. Transl. Med. 2016, 14, 344. [Google Scholar] [CrossRef] [Green Version]
- Fisher, C.R.; Ferrington, D.A. Perspective on AMD Pathobiology: A Bioenergetic Crisis in the RPE. Investig. Ophthalmol. Vis. Sci. 2018, 59, AMD41–AMD47. [Google Scholar] [CrossRef] [Green Version]
- Feher, J.; Kovacs, I.; Artico, M.; Cavallotti, C.; Papale, A.; Balacco Gabrieli, C. Mitochondrial Alterations of Retinal Pigment Epithelium in Age-Related Macular Degeneration. Neurobiol. Aging 2006, 27, 983–993. [Google Scholar] [CrossRef]
- Ferrington, D.A.; Ebeling, M.C.; Kapphahn, R.J.; Terluk, M.R.; Fisher, C.R.; Polanco, J.R.; Roehrich, H.; Leary, M.M.; Geng, Z.; Dutton, J.R.; et al. Altered Bioenergetics and Enhanced Resistance to Oxidative Stress in Human Retinal Pigment Epithelial Cells from Donors with Age-Related Macular Degeneration. Redox Biol. 2017, 13, 255–265. [Google Scholar] [CrossRef]
- Terluk, M.R.; Kapphahn, R.J.; Soukup, L.M.; Gong, H.; Gallardo, C.; Montezuma, S.R.; Ferrington, D.A. Investigating Mitochondria as a Target for Treating Age-Related Macular Degeneration. J. Neurosci. 2015, 35, 7304–7311. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Gottlieb, E.; Brooks, D.G.; Murphy, M.P.; Dunaief, J.L. Mitochondria-Derived Reactive Oxygen Species Mediate Blue Light–Induced Death of Retinal Pigment Epithelial Cells. Photochem. Photobiol. 2004, 79, 470. [Google Scholar] [CrossRef]
- Meyer, J.G.; Garcia, T.Y.; Schilling, B.; Gibson, B.W.; Lamba, D.A. Proteome and Secretome Dynamics of Human Retinal Pigment Epithelium in Response to Reactive Oxygen Species. Sci. Rep. 2019, 9, 15440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravingerová, T.; Barančík, M.; Strnisková, M. Mitogen-Activated Protein Kinases: A New Therapeutic Target in Cardiac Pathology. Mol. Cell. Biochem. 2003, 247, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R. ERK1/2 MAP Kinases: Structure, Function, and Regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- di Mari, J.F.; Davis, R.; Safirstein, R.L. MAPK Activation Determines Renal Epithelial Cell Survival during Oxidative Injury. Am. J. Physiol. Physiol. 1999, 277, F195–F203. [Google Scholar] [CrossRef] [PubMed]
- Arany, I.; Megyesi, J.K.; Kaneto, H.; Price, P.M.; Safirstein, R.L. Cisplatin-Induced Cell Death Is EGFR/Src/ERK Signaling Dependent in Mouse Proximal Tubule Cells. Am. J. Physiol. Physiol. 2004, 287, F543–F549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glotin, A.-L.; Calipel, A.; Brossas, J.-Y.; Faussat, A.-M.; Tre’ton, J.; Mascarelli, F. Sustained versus Transient ERK1/2 Signaling Underlies the Anti- and Proapoptotic Effects of Oxidative Stress in Human RPE Cells. Investig. Opthalmology Vis. Sci. 2006, 47, 4614. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qu, Y.; Shen, X.Y.; Ouyang, T.; Fu, W.B.; Luo, T.; Wang, H.Q. Multiple Signal Pathways Involved in Crocetin-Induced Apoptosis in KYSE-150 Cells. Pharmacology 2019, 103, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Qian, Z.; Wen, N.; Xi, L. Crocetin Suppresses Angiotensin II-Induced Vascular Smooth-Muscle Cell Proliferation through Inhibition of ERK1/2 Activation and Cell-Cycle Progression. J. Cardiovasc. Pharmacol. 2007, 50, 519–525. [Google Scholar] [CrossRef]
- Zhang, H.; Shang, Q.; An, J.; Wang, C.; Ma, J. Crocetin Inhibits PDGF-BB-Induced Proliferation and Migration of Retinal Pigment Epithelial Cells. Eur. J. Pharmacol. 2019, 842, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Kassoff, A.; Kassoff, J.; Buehler, J.; Eglow, M.; Kaufman, F.; Mehu, M.; Kieval, S.; Mairs, M.; Graig, B.; Quattrocchi, A.; et al. A Randomized, Placebo-Controlled, Clinical Trial of High-Dose Supplementation with Vitamins C and E, Beta Carotene, and Zinc for Age-Related Macular Degeneration and Vision Loss: AREDS Report No. 8. Arch. Ophthalmol. 2001, 119, 1417–1436. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Liu, Z.; Li, X.; Jia, H.; Sun, L.; Tian, C.; Jia, L.; Liu, J. α-Tocopherol Is an Effective Phase II Enzyme Inducer: Protective Effects on Acrolein-Induced Oxidative Stress and Mitochondrial Dysfunction in Human Retinal Pigment Epithelial Cells. J. Nutr. Biochem. 2010, 21, 1222–1231. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Winborn, C.S.; Thomas, F.; Tran, Q.T.; Lang, J.C.; Chaum, E. Modulation of Oxidative Stress Responses by Vitamin E, or Vitamin A plus Vitamin C Treatment in Human Retinal Pigment Epithelial Cells. Nat. Sci. 2015, 7, 558–570. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Linkert, M.; Rueden, C.T.; Allan, C.; Burel, J.-M.; Moore, W.; Patterson, A.; Loranger, B.; Moore, J.; Neves, C.; MacDonald, D.; et al. Metadata Matters: Access to Image Data in the Real World. J. Cell Biol. 2010, 189, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Forster, B.; Van De Ville, D.; Berent, J.; Sage, D.; Unser, M. Complex Wavelets for Extended Depth-of-Field: A New Method for the Fusion of Multichannel Microscopy Images. Microsc. Res. Tech. 2004, 65, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang, I.; Friman, O.; Guertin, D.A.; Chang, J.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image Analysis Software for Identifying and Quantifying Cell Phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimi, P.; Gheisari, A.; Gasparini, S.J.; Baharvand, H.; Shekari, F.; Satarian, L.; Ader, M. Crocetin Prevents RPE Cells from Oxidative Stress through Protection of Cellular Metabolic Function and Activation of ERK1/2. Int. J. Mol. Sci. 2020, 21, 2949. https://doi.org/10.3390/ijms21082949
Karimi P, Gheisari A, Gasparini SJ, Baharvand H, Shekari F, Satarian L, Ader M. Crocetin Prevents RPE Cells from Oxidative Stress through Protection of Cellular Metabolic Function and Activation of ERK1/2. International Journal of Molecular Sciences. 2020; 21(8):2949. https://doi.org/10.3390/ijms21082949
Chicago/Turabian StyleKarimi, Padideh, Ali Gheisari, Sylvia J Gasparini, Hossein Baharvand, Faezeh Shekari, Leila Satarian, and Marius Ader. 2020. "Crocetin Prevents RPE Cells from Oxidative Stress through Protection of Cellular Metabolic Function and Activation of ERK1/2" International Journal of Molecular Sciences 21, no. 8: 2949. https://doi.org/10.3390/ijms21082949
APA StyleKarimi, P., Gheisari, A., Gasparini, S. J., Baharvand, H., Shekari, F., Satarian, L., & Ader, M. (2020). Crocetin Prevents RPE Cells from Oxidative Stress through Protection of Cellular Metabolic Function and Activation of ERK1/2. International Journal of Molecular Sciences, 21(8), 2949. https://doi.org/10.3390/ijms21082949