Kisspeptin and Testicular Function—Is It Necessary?

 ,
,
Abstract
:1. Introduction
2. Testicular Function
3. Distribution of Kisspeptin and Its Receptor
4. Regulation of Testicular Kisspeptin Signaling
5. Effects of Kisspeptin on Sperm Function
6. Effects of Kisspeptin on Testicular Steroidogenesis:
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GnRH | gonadotrophin-releasing hormone |
| LH | luteinising hormone |
| FSH | follicle-stimulating hormone |
| ST | seminiferous tubules |
| HPG | hypothalamic-pituitary-gonadal axis |
| NKB | Neurokinin B |
| GnIH | Gonadotrophin inhibiting hormone |
| RT-PCR | Reverse transcriptase-polymerase chain reaction |
| ARC | arcuate nucleus |
| E2 | oestradiol |
| kp | kisspeptin |
| s/c | subcutaneous |
| ip | intraperitoneal |
| hCG | human chorionic gonadotropin |
| GnRH-R | GnRH receptor |
| CYP11A1 | cytochrome P450 cholesterol side-chain cleavage |
| Esr1 | oestrogen receptor |
| LHCGR | luteinizing hormone choriogonadotropin receptor |
| StAR | steroidogenic acute regulatory protein |
| Ca2+ | calcium |
| IVF | in vitro fertilisation |
| GSI | gonadosomatic index |
| HSD3B1 | 3-bhydroxysteroid dehydrogenase |
| IV | intravenous |
References
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G. The GPR54 Gene as a Regulator of Puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahab, M.; Mastronardi, C.; Seminara, S.B.; Crowley, W.F.; Ojeda, S.R.; Plant, T.M. Increased Hypothalamic GPR54 Signaling: A Potential Mechanism for Initiation of Puberty in Primates. Proc. Natl. Acad. Sci. USA 2005, 102, 2129–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Gottsch, M.L.; Lee, K.J.; Popa, S.M.; Smith, J.T.; Jakawich, S.K.; Clifton, D.K.; Steiner, R.A.; Herbison, A.E. Activation of Gonadotropin-Releasing Hormone Neurons by Kisspeptin as a Neuroendocrine Switch for the Onset of Puberty. J. Neurosci. 2005, 25, 11349–11356. [Google Scholar] [CrossRef] [PubMed]
- Dhillo, W.S. Kisspeptin: A Novel Regulator of Reproductive Function. J. Neuroendocrinol. 2008, 20, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and Reproduction: Physiological Roles and Regulatory Mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef]
- Dhillo, W.S.; Chaudhri, O.B.; Patterson, M.; Thompson, E.L.; Murphy, K.G.; Badman, M.K.; McGowan, B.M.; Amber, V.; Patel, S.; Ghatei, M.A.; et al. Kisspeptin-54 Stimulates the Hypothalamic-Pituitary Gonadal Axis in Human Males. J. Clin. Endocrinol. Metab. 2005, 90, 6609–6615. [Google Scholar] [CrossRef] [Green Version]
- De Roux, N.; Genin, E.; Carel, J.C.; Matsuda, F.; Chaussain, J.L.; Milgrom, E. Hypogonadotropic Hypogonadism Due to Loss of Function of the KISS1-Derived Peptide Receptor GPR54. Proc. Natl. Acad. Sci. USA 2003, 100, 10972–10976. [Google Scholar] [CrossRef] [Green Version]
- Colledge, W.H. Kisspeptins and GnRH neuronal signaling. Trends Endocrinol. Metab. 2008, 20, 115–121. [Google Scholar] [CrossRef]
- Topaloglu, A.K.; Tello, J.A.; Kotan, L.D.; Ozbek, M.N.; Yilmaz, M.B.; Erdogan, S.; Gurbuz, F.; Temiz, F.; Millar, R.P.; Yuksel, B. Inactivating KISS1 Mutation and Hypogonadotropic Hypogonadism. N. Engl. J. Med. 2012, 366, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Ratnasabapathy, R.; Dhillo, W.S. The Effects of Kisspeptin in Human Reproductive Function—Therapeutic Implications. Curr. Drug Targets 2013, 14, 365–371. [Google Scholar] [CrossRef]
- Ohtaki, T.; Shintani, Y.; Honda, S.; Matsumoto, H.; Hori, A.; Kanehashi, K.; Terao, Y.; Kumano, S.; Takatsu, Y.; Masuda, Y.; et al. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001, 411, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Doran, J.; Kyle, V.; Yeo, S.; Colledge, W.H. Does Kisspeptin Signaling have a Role in the Testes? Front. Endocrinol. 2013, 4, 198. [Google Scholar] [CrossRef] [PubMed]
- Terao, Y.; Kumano, S.; Takatsu, Y.; Hattori, M.; Nishimura, A.; Ohtaki, T.; Shintani, Y. Expression of KiSS-1, a metastasis suppressor gene, in trophoblast giant cells of the rat placenta. Bba Gene Struct. Expr. 2004, 1678, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhu, E.; Yang, C.; Bentley, G.E.; Tsutsui, K.; Kriegsfeld, L.J. RFamide-Related Peptide and Messenger Ribonucleic Acid Expression in Mammalian Testis: Association with the Spermatogenic Cycle. Endocrinology 2010, 151, 617–627. [Google Scholar] [CrossRef] [Green Version]
- León, S.; Barroso, A.; Vázquez, M.J.; García-Galiano, D.; Manfredi-Lozano, M.; Ruiz-Pino, F.; Heras, V.; Romero-Ruiz, A.; Roa, J.; Schutz, G.; et al. Direct Actions of Kisspeptins on GnRH Neurons Permit Attainment of Fertility but are Insufficient to Fully Preserve Gonadotropic Axis Activity. Sci. Rep. 2016, 6, 19206. [Google Scholar] [CrossRef]
- Pinto, F.M.; Cejudo-Román, A.; Ravina, C.G.; Fernández-Sánchez, M.; Martín-Lozano, D.; Illanes, M.; Tena-Sempere, M.; Candenas, M.L. Characterization of the kisspeptin system in human spermatozoa. Int. J. Androl. 2012, 35, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Shahab, M.; Clarke, I.; Pereira, A.; Smith, J.; Khan, S.; Sultan, J.; Javed, S.; Anwar, T. KISS1 and KISS1 receptor expression in the rhesus monkey testis: A possible local regulator of testicular function. Cent. Eur. J. Biol. 2013, 8, 968–974. [Google Scholar] [CrossRef] [Green Version]
- Irfan, S.; Ehmcke, J.; Shahab, M.; Wistuba, J.; Schlatt, S. Immunocytochemical localization of kisspeptin and kisspeptin receptor in the primate testis. J. Med. Primatol. 2016, 45, 105–111. [Google Scholar] [CrossRef]
- Anjum, S.; Krishna, A.; Sridaran, R.; Tsutsui, K. Localization of Gonadotropin-Releasing Hormone [GnRH], Gonadotropin-Inhibitory Hormone [GnIH], Kisspeptin and GnRH Receptor and Their Possible Roles in Testicular Activities From Birth to Senescence in Mice. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2012, 317, 630–644. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.; Wang, J.; Lee, Y.; Jong, D.; Tsui, K.; Chiu, C. Kisspeptin modulates fertilization capacity of mouse spermatozoa. Reproduction 2014, 147, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Salehi, S.; Adeshina, I.; Chen, H.; Zirkin, B.R.; Hussain, M.A.; Wondisford, F.; Wolfe, A.; Radovick, S. Developmental and Endocrine Regulation of Kisspeptin Expression in Mouse Leydig Cells. Endocrinology 2015, 156, 1514–1522. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, R.; Chianese, R.; Chioccarelli, T.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Cobellis, G. Intra-Testicular Signals Regulate Germ Cell Progression and Production of Qualitatively Mature Spermatozoa in Vertebrates. Front. Endocrinol. 2014, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Chianese, R.; Cobellis, G.; Chioccarelli, T.; Ciaramella, V.; Migliaccio, M.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptins, Estrogens and Male Fertility. Curr. Med. Chem. 2016, 23, 4070. [Google Scholar] [CrossRef]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin receptor, GPR54, as a candidate for the regulation of testicular activity in the frog Rana esculenta. Biol. Reprod. 2013, 88, 73. [Google Scholar] [CrossRef]
- Selvaraj, S.; Ohga, H.; Kitano, H.; Nyuji, M.; Yamaguchi, A.; Matsuyama, M. Peripheral Administration of KISS1 Pentadecapeptide Induces Gonadal Development in Sexually Immature Adult Scombroid Fish. Zool. Sci. 2013, 30, 446–454. [Google Scholar] [CrossRef]
- Selvaraj, S.; Ohga, H.; Nyuji, M.; Kitano, H.; Nagano, N.; Yamaguchi, A.; Matsuyama, M. Subcutaneous administration of KISS1 pentadecapeptide accelerates spermatogenesis in prepubertal male chub mackerel [Scomber japonicus]. Comp. Biochem. Physiol. Part A 2013, 166, 228–236. [Google Scholar] [CrossRef]
- Gilbert, S.F. Developmental Biology, 7th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2003. [Google Scholar]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr. Biol. 2017, 27, R988–R994. [Google Scholar] [CrossRef] [Green Version]
- Naz, R.K.; Rajesh, P.B. Role of tyrosine phosphorylation in sperm capacitation/acrosome reaction. Reprod. Biol. Endocrinol. 2004, 2, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Kawano, N.; Yoshida, K. Control of sperm motility and fertility: Diverse factors and common mechanisms. Cell. Mol. Life Sci. 2008, 65, 3446–3457. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.M.; Bremner, W.J. Endocrinology of the hypothalamic-pituitary-testicular axis with particular reference to the hormonal control of spermatogenesis. Bailliere’s Clin. Endocrinol. Metab. 1987, 1, 71. [Google Scholar] [CrossRef]
- Clavijo, R.I.; Hsiao, W. Update on male reproductive endocrinology. Transl. Androl. Urol. 2018, 7, S367–S372. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Tsunekawa, K.; Moon, M.J.; Um, H.N.; Hwang, J.; Osugi, T.; Otaki, N.; Sunakawa, Y.; Kim, K.; Vaudry, H.; et al. Molecular Evolution of Multiple Forms of Kisspeptins and GPR54 Receptors in Vertebrates. Endocrinology 2009, 150, 2837–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.S.; Lee, Y.R.; Lee, J.Y.; Oh, D.Y.; Hwang, J.I.; Kim, J.I.; Vaudry, H.; Kwon, H.B.; Seong, J.Y. Molecular cloning of the bullfrog kisspeptin receptor GPR54 with high sensitivity to Xenopus kisspeptin. Peptides 2009, 30, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Tena-Sempere, M.; Felip, A.; Gómez, A.; Zanuy, S.; Carrillo, M. Comparative insights of the kisspeptin/kisspeptin receptor system: Lessons from non-mammalian vertebrates. Gen. Comp. Endocrinol. 2011, 175, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Um, H.N.; Han, J.M.; Hwang, J.; Hong, S.I.; Vaudry, H.; Seong, J.Y. Molecular coevolution of kisspeptins and their receptors from fish to mammals. Ann. N. Y. Acad. Sci. 2010, 1200, 67–74. [Google Scholar] [CrossRef]
- Bedford, J.M.; Moore, H.D.M.; Franklin, L.E. Significance of the equatorial segment of the acrosome of the spermatozoon in eutherian mammals. Exp. Cell Res. 1979, 119, 119–126. [Google Scholar] [CrossRef]
- Flesch, F.M.; Gadella, B.M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. BBA Rev. Biomembr. 2000, 1469, 197–235. [Google Scholar] [CrossRef]
- Lehman, M.N.; Coolen, L.M.; Goodman, R.L. Minireview: Kisspeptin/Neurokinin B/Dynorphin [KNDy] Cells of the Arcuate Nucleus: A Central Node in the Control of Gonadotropin-Releasing Hormone Secretion. Endocrinology 2010, 151, 3479–3489. [Google Scholar] [CrossRef]
- Topaloglu, A.K.; Reimann, F.; Guclu, M.; Yalin, A.S.; Kotan, L.D.; Porter, K.M.; Serin, A.; Mungan, N.O.; Cook, J.R.; Ozbek, M.N.; et al. TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for Neurokinin B in the central control of reproduction. Nat. Genet. 2009, 41, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Moreno, C.; Boilard, M.; Sullivan, R.; Sirard, M.A. Characterization and Identification of Epididymal Factors That Protect Ejaculated Bovine Sperm During In Vitro Storage. Biol. Reprod. 2002, 66, 159. [Google Scholar] [CrossRef]
- Wang, J.; Hsu, M.; Tseng, T.; Wu, L.; Yang, K.; Chiu, C. Kisspeptin expression in mouse Leydig cells correlates with age. J. Chin. Med. Assoc. 2015, 78, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin regulates steroidogenesis and spermiation in anuran amphibian. Reproduction 2017, 154, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Li, Z.; Jiang, W.; Ling, Y.; Kuang, H. Reproductive functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod. Biol. Endocrinol. 2019, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Wang, X.; Chen, Q.; Yang, H.; Zhou, N.; Sun, L.; Chen, H.; Liu, J.; Ao, L.; Cui, Z.; et al. Kisspeptin Protein in Seminal Plasma Is Positively Associated with Semen Quality: Results from the MARHCS Study in Chongqing, China. Biomed Res. Int. 2019, 2019, 5129263–5129269. [Google Scholar] [CrossRef]
- Rehman, R.; Alam, A.; Zafar, S. Kisspeptin levels in infertile male subjects with abnormal sperm parameters. J. Pak. Med Assoc. 2019, 69, 1227. [Google Scholar]
- Toolee, H.; Rastegar, T.; Solhjoo, S.; Mortezaee, K.; Mohammadipour, M.; Kashani, I.R.; Akbari, M. Roles for Kisspeptin in proliferation and differentiation of spermatogonial cells isolated from mice offspring when the cells are cocultured with somatic cells. J. Cell. Biochem. 2019, 120, 5042–5054. [Google Scholar] [CrossRef]
- Ramzan, F.; Qureshi, I.Z. Intraperitoneal kisspeptin-10 administration induces dose-dependent degenerative changes in maturing rat testes. Life Sci. 2011, 88, 246–256. [Google Scholar] [CrossRef]
- Thompson, E.L.; Amber, V.; Stamp, G.; Patterson, M.; Curtis, A.E.; Cooke, J.H.; Appleby, G.; Dhillo, W.; Ghatel, M.; Bloom, S.; et al. Kisspeptin-54 at high doses acutely induces testicular degeneration in adult male rats via central mechanisms. Br. J. Pharmacol. 2009, 156, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Aytürk, N.; Firat, T.; Kükner, A.; Özoğul, C.; Töre, F.; Kandirali, İ.E.; Yilmaz, B. The effect of kisspeptin on spermatogenesis and apoptosis in rats. Turk. J. Med Sci. 2017, 47, 334–342. [Google Scholar] [CrossRef]
- Tariq, A.R.; Shabab, M. Effect of kisspeptin challenge on testosterone and inhibin secretion from in vitro testicular tissue of adult male rhesus monkey [Macaca mulatta]. Andrologia 2017, 49, e12590. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, S.; Seminara, S.B.; Pohl, C.R.; DiPietro, M.J.; Crowley, W.F.; Plant, T.M. Effect of Continuous Intravenous Administration of Human Metastin 45–54 on the Neuroendocrine Activity of the Hypothalamic-Pituitary-Testicular Axis in the Adult Male Rhesus Monkey [Macaca mulatta]. Endocrinology 2007, 148, 3364–3370. [Google Scholar] [CrossRef]
- Irfan, S.; Ehmcke, J.; Wahab, F.; Shahab, M.; Schlatt, S. Intratesticular action of kisspeptin in rhesus monkey [Macaca mulatta]. Andrologia 2014, 46, 610–617. [Google Scholar] [CrossRef] [PubMed]
- McGuire, N.L.; Bentley, G.E. Neuropeptides in the gonads: From evolution to pharmacology. Front. Pharmacol. 2010, 1, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, J.; Herbison, A.E. Postnatal Development of Kisspeptin Neurons in Mouse Hypothalamus; Sexual Dimorphism and Projections to Gonadotropin-Releasing Hormone Neurons. Endocrinology 2006, 147, 5817–5825. [Google Scholar] [CrossRef] [PubMed]
- Pierantoni, R.; Cobellis, G.; Meccariello, R.; Fasano, S. Evolutionary Aspects Of Cellular Communication In The Vertebrate Hypothalamo–Hypophysio–Gonadal Axis. Int. Rev. Cytol. 2002, 218, 69–141. [Google Scholar] [CrossRef]
- Sharpe, R.M. Paracrine control of the testis. Clin. Endocrinol. Metab. 1986, 15, 185. [Google Scholar] [CrossRef]
- Morales, P.; Pizarro, E.; Kong, M.; Kerr, B.; Ceric, F.; Vigil, P. Gonadotropin-Releasing Hormone-Stimulated Sperm Binding to the Human Zona Is Mediated by a Calcium Influx. Biol. Reprod. 2000, 63, 635. [Google Scholar] [CrossRef] [Green Version]
- Morales, P.; Pasten, C.; Pizarro, E. Inhibition of In Vivo and In Vitro Fertilization in Rodents by Gonadotropin-Releasing Hormone Antagonists. Biol. Reprod. 2002, 67, 1360. [Google Scholar] [CrossRef] [Green Version]
- Tena-Sempere, M. GPR54 and kisspeptin in reproduction. Hum. Reprod. Update. 2006, 12, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Castellano, J.M.; Gaytan, M.; Roa, J.; Vigo, E.; Navarro, V.M.; Bellido, C.; Dieguez, C.; Aguilar, E.; Sanchez-Criado, J.E.; Pellicer, A.; et al. Expression of KiSS-1 in Rat Ovary: Putative Local Regulator of Ovulation? Endocrinology 2006, 147, 4852–4862. [Google Scholar] [CrossRef]
- Gaytán, F.; Gaytán, M.; Castellano, J.M.; Romero, M.; Roa, J.; Aparicio, B.; Garrido, N.; Sanchez-Criado, J.E.; Millar, R.P.; Pelicer, A.; et al. KiSS-1 in the mammalian ovary: Distribution of kisspeptin in human and marmoset and alterations in KiSS-1 mRNA levels in a rat model of ovulatory dysfunction. Am. J. Physiol. Endocrinol. Metab. 2009, 296, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin drives germ cell progression in the anuran amphibian Pelophylax esculentus: A study carried out in ex vivo testes. Gen. Comp. Endocrinol. 2014, 211, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Pierantoni, R.; Cobellis, R.; Meccariello, C.; Palmiero, G.; Minucci, F.S.; Fasano, S. The Amphibian Testis As Model To Study Germ Cell Progression During Spermatogenesis. Comparative Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 132, 131–139. [Google Scholar] [CrossRef]
- Roseweir, A.K.; Kauffman, A.S.; Smith, J.T.; Guerriero, K.A.; Morgan, K.; Pielecka-Fortuna, J.; Pineda, R. Discovery Of Potent Kisspeptin Antagonists Delineate Physiological Mechanisms Of Gonadotropin Regulation. J. Neurosci. 2009, 29, 3920–3929. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, C.N.; Nijher, G.M.K.; Chaudhri, O.B.; Murphy, K.G.; Ranger, A.; Lim, A.; Patel, D.; Mehta, A.; Todd, C.; Ramachandran, R. Subcutaneous Injection of Kisspeptin-54 Acutely Stimulates Gonadotropin Secretion in Women with Hypothalamic Amenorrhea, But Chronic Administration Causes Tachyphylaxis. J. Clin. Endocrinol. Metab. 2009, 94, 4315–4323. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Ramzan, F.; Qureshi, I.Z.; Ramzan, M.; Ramzan, M.H.; Ramzan, F. Immature rat seminal vesicles show histomorphological and ultrastructural alterations following treatment with kisspeptin-10. Reprod. Biol. Endocrinol. 2012, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, F.; Ramzan, F.; Khan, M.; Khan, M.; Ramzan, M.; Ramzan, M. The effect of chronic kisspeptin administration on seminal fructose levels in male mice. Endocrine 2014, 45, 144–147. [Google Scholar] [CrossRef]
- O’Donnell, L.; Nicholls, P.K.; O’Bryan, M.K.; McLachlan, R.I.; Stanton, P.G. Spermiation. Spermatogenesis 2011, 1, 14–35. [Google Scholar] [CrossRef]
- Mei, H.; Walters, C.; Carter, R.; Colledge, W.H. Gpr54−/− mice show more pronounced defects in spermatogenesis than KISS1−/− mice and improved spermatogenesis with age when exposed to dietary phytoestrogens. Reproduction 2011, 141, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Stocco, D.M. StAR Protein and the Regulation of Steroid Hormone Biosynthesis. Annu. Rev. Physiol. 2001, 63, 193–213. [Google Scholar] [CrossRef]
- Huma, T.; Ulla, F.; Hanif, F.; Rizak, J.D.; Shahab, M. Peripheral administration of kisspeptin antagonist does not alter basal plasma testosterone but decreases plasma adiponectin levels in adult male rhesus macaques. Turk. J. Biol. 2014, 38, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Pierik, F.; Burdorf, A.; de Jong, F.; Weber, R. Inhibin B: A novel marker of spermatogenesis. Ann. Med. 2003, 35, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, F.; Qureshi, I.Z.; Ramzan, M.; Ramzan, M.H.; Ramzan, F. Kisspeptin-10 induces dose dependent degeneration in prepubertal rat prostate gland. Prostate 2013, 73, 690–699. [Google Scholar] [CrossRef] [PubMed]


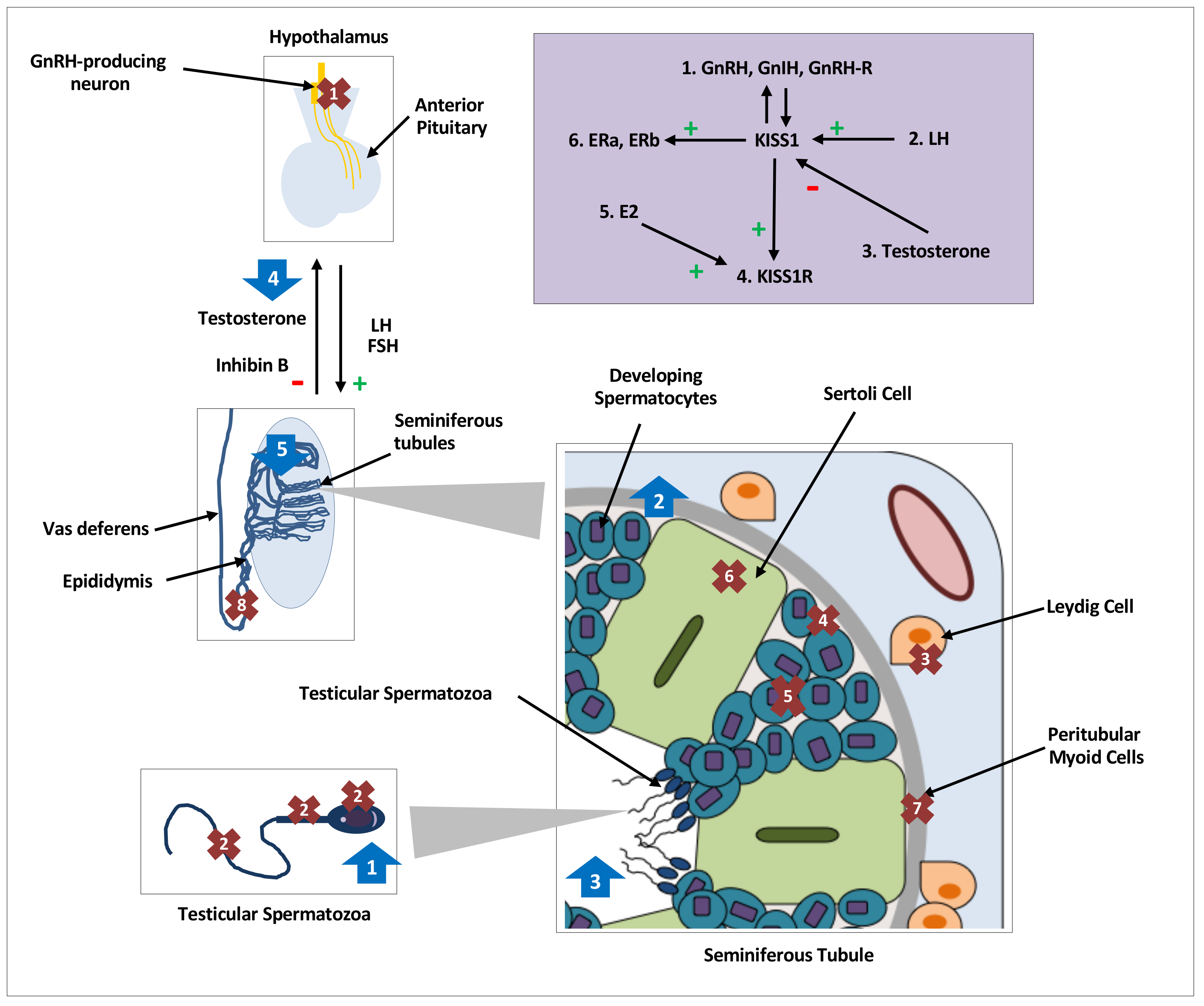{kind=link}
| Species | Distribution of KISS1 & KISSR | Regulation | Effects on Sperm Function | Effects on Testicular Steroidogenesis |
|---|---|---|---|---|
| Human | ||||
| Mice |
| |||
| Monkey | ||||
| Frog |
|
|
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Thaventhiran, T.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Kisspeptin and Testicular Function—Is It Necessary? Int. J. Mol. Sci. 2020, 21, 2958. https://doi.org/10.3390/ijms21082958
Sharma A, Thaventhiran T, Minhas S, Dhillo WS, Jayasena CN. Kisspeptin and Testicular Function—Is It Necessary? International Journal of Molecular Sciences. 2020; 21(8):2958. https://doi.org/10.3390/ijms21082958
Chicago/Turabian StyleSharma, Aditi, Thilipan Thaventhiran, Suks Minhas, Waljit S. Dhillo, and Channa N. Jayasena. 2020. "Kisspeptin and Testicular Function—Is It Necessary?" International Journal of Molecular Sciences 21, no. 8: 2958. https://doi.org/10.3390/ijms21082958
APA StyleSharma, A., Thaventhiran, T., Minhas, S., Dhillo, W. S., & Jayasena, C. N. (2020). Kisspeptin and Testicular Function—Is It Necessary? International Journal of Molecular Sciences, 21(8), 2958. https://doi.org/10.3390/ijms21082958