Probing Hexaminolevulinate Mediated PpIX Fluorescence in Cancer Cell Suspensions in the Presence of Chemical Adjuvants



Abstract
:
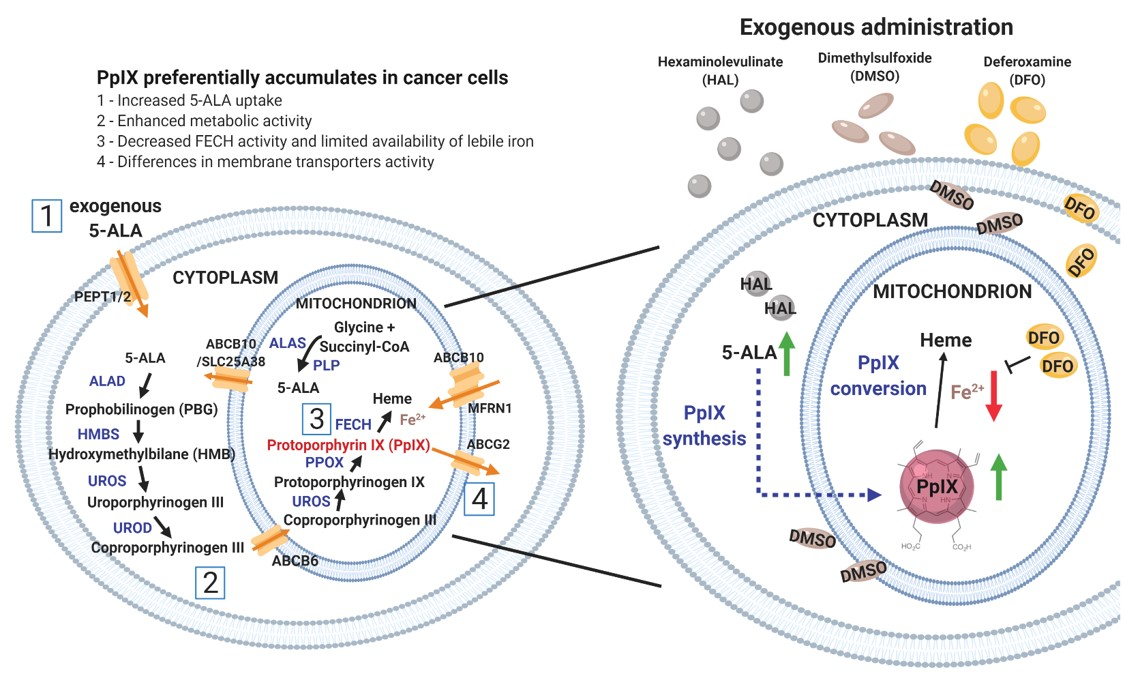
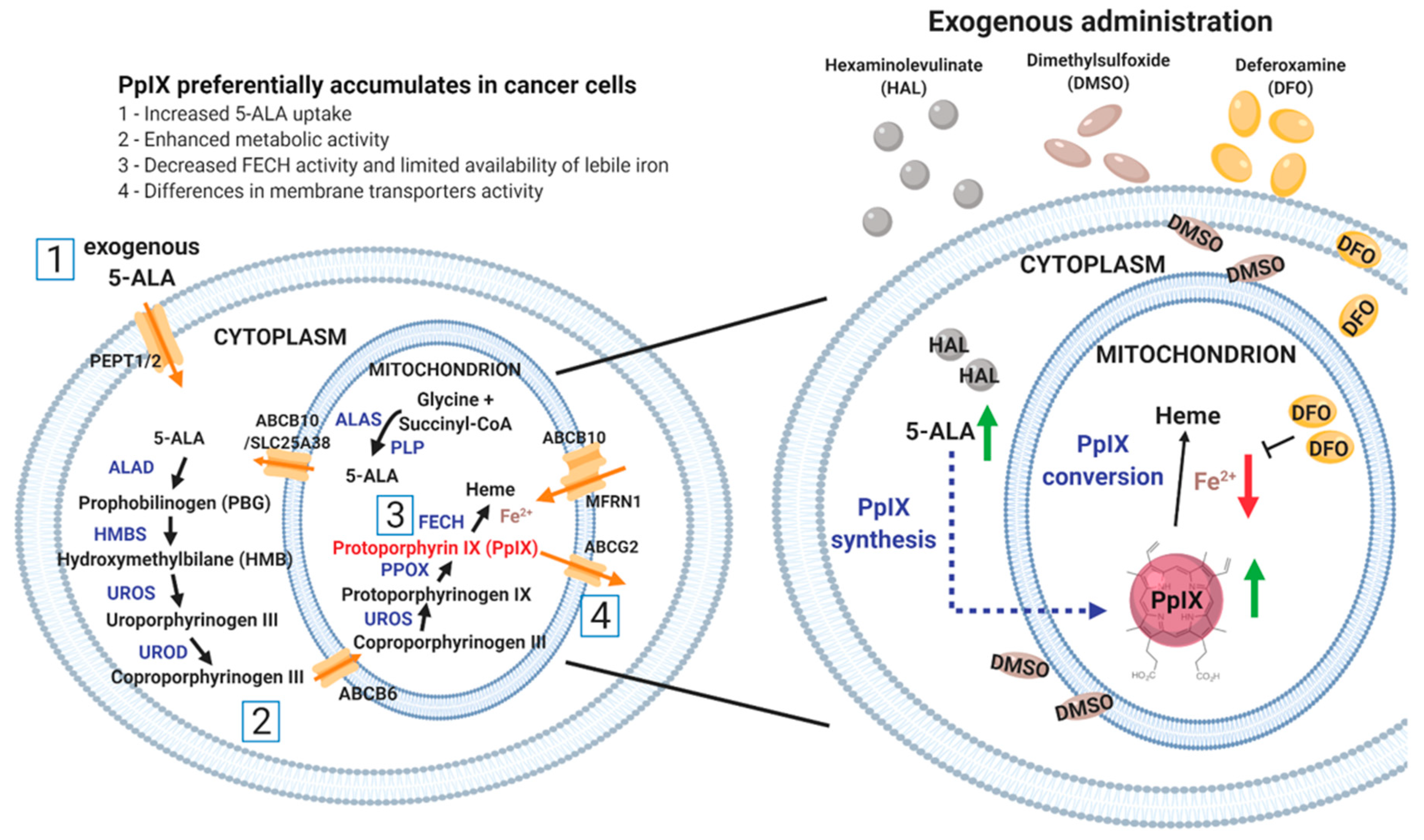
1. Introduction
2. Results
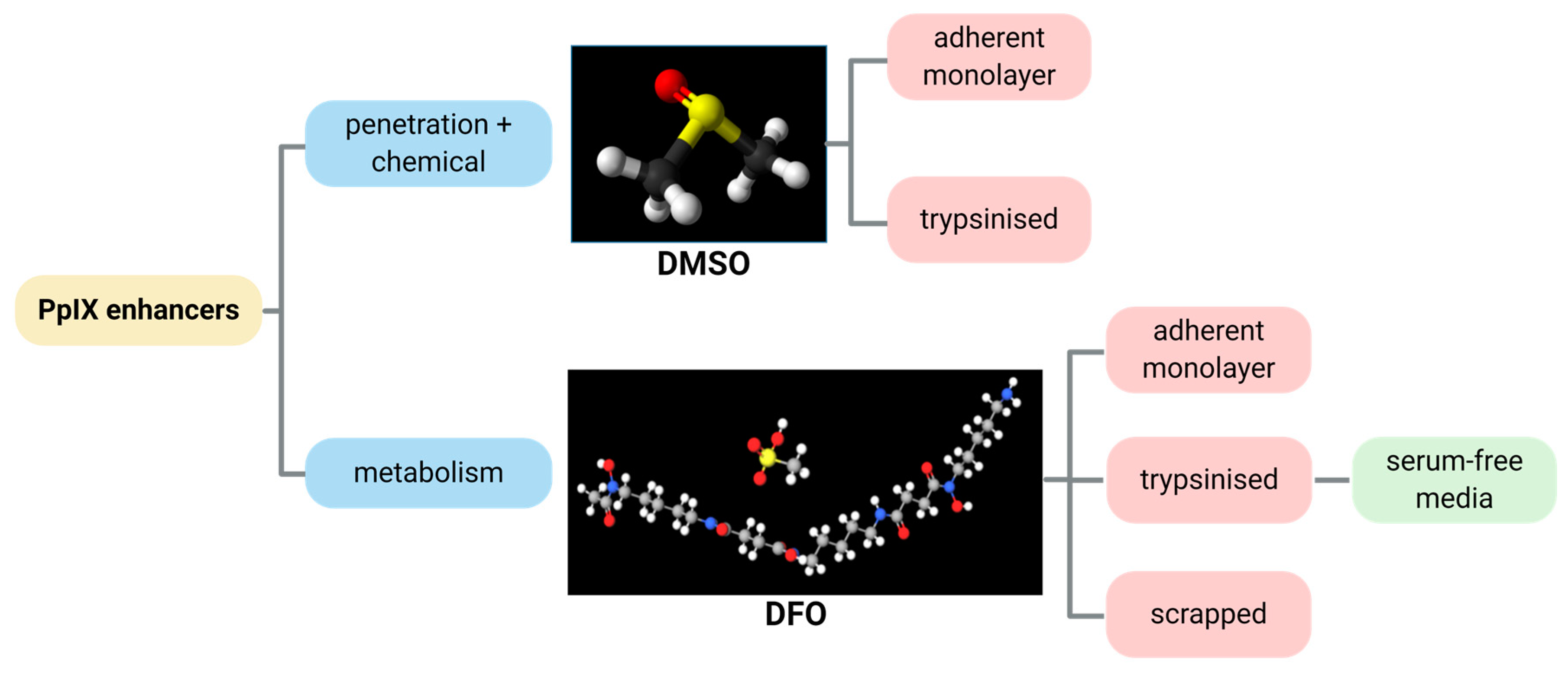
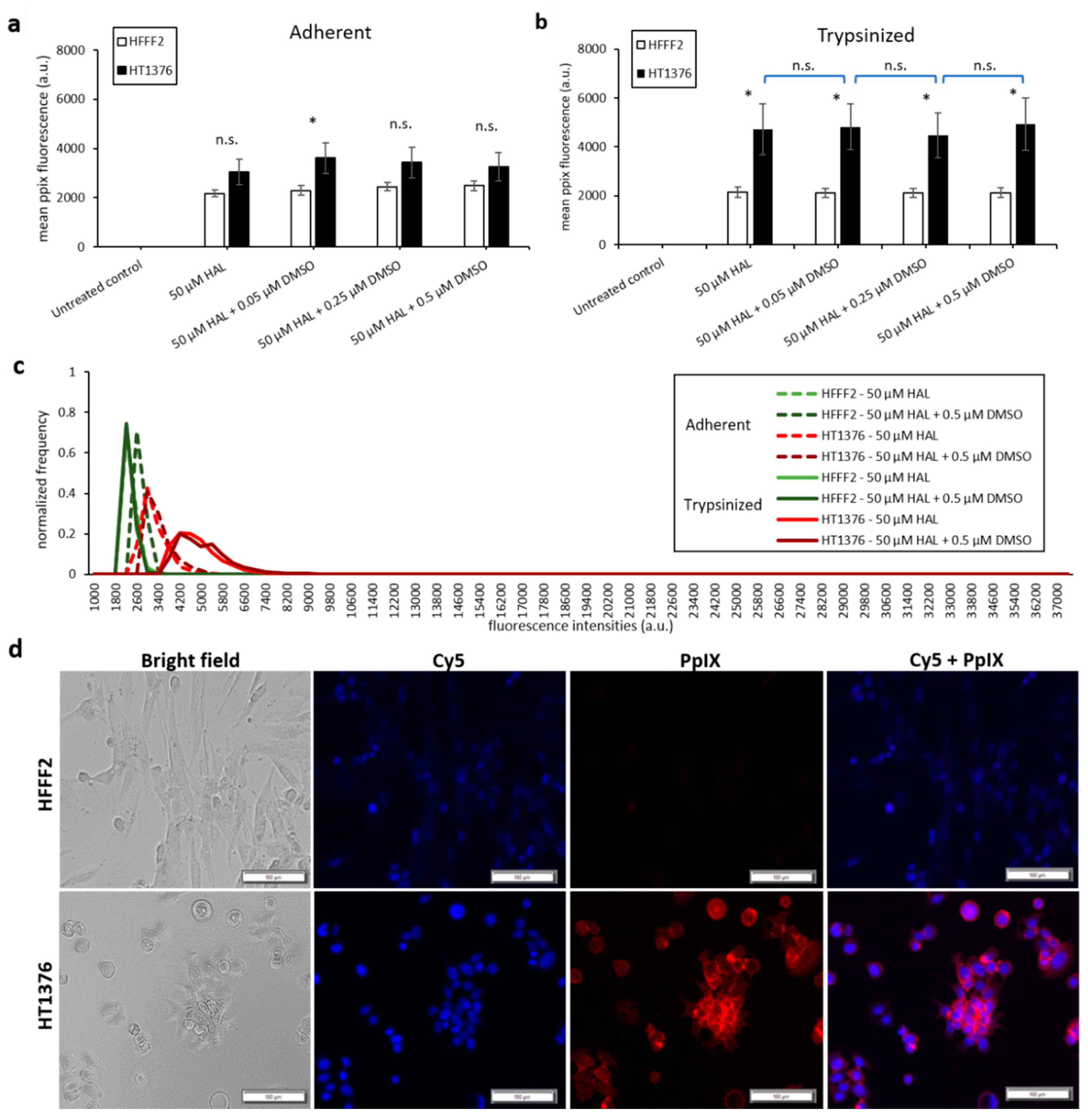
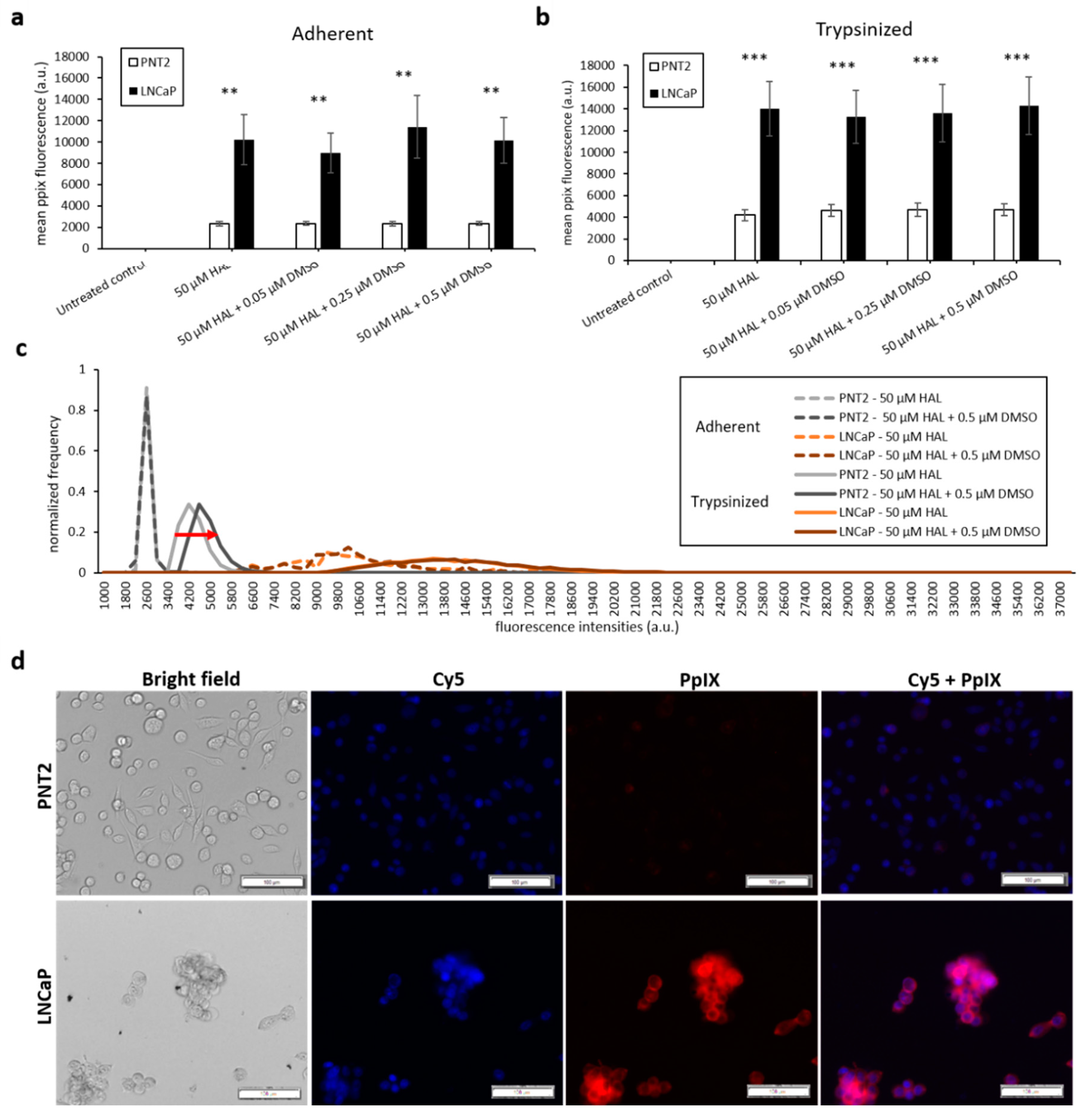
2.1. DMSO
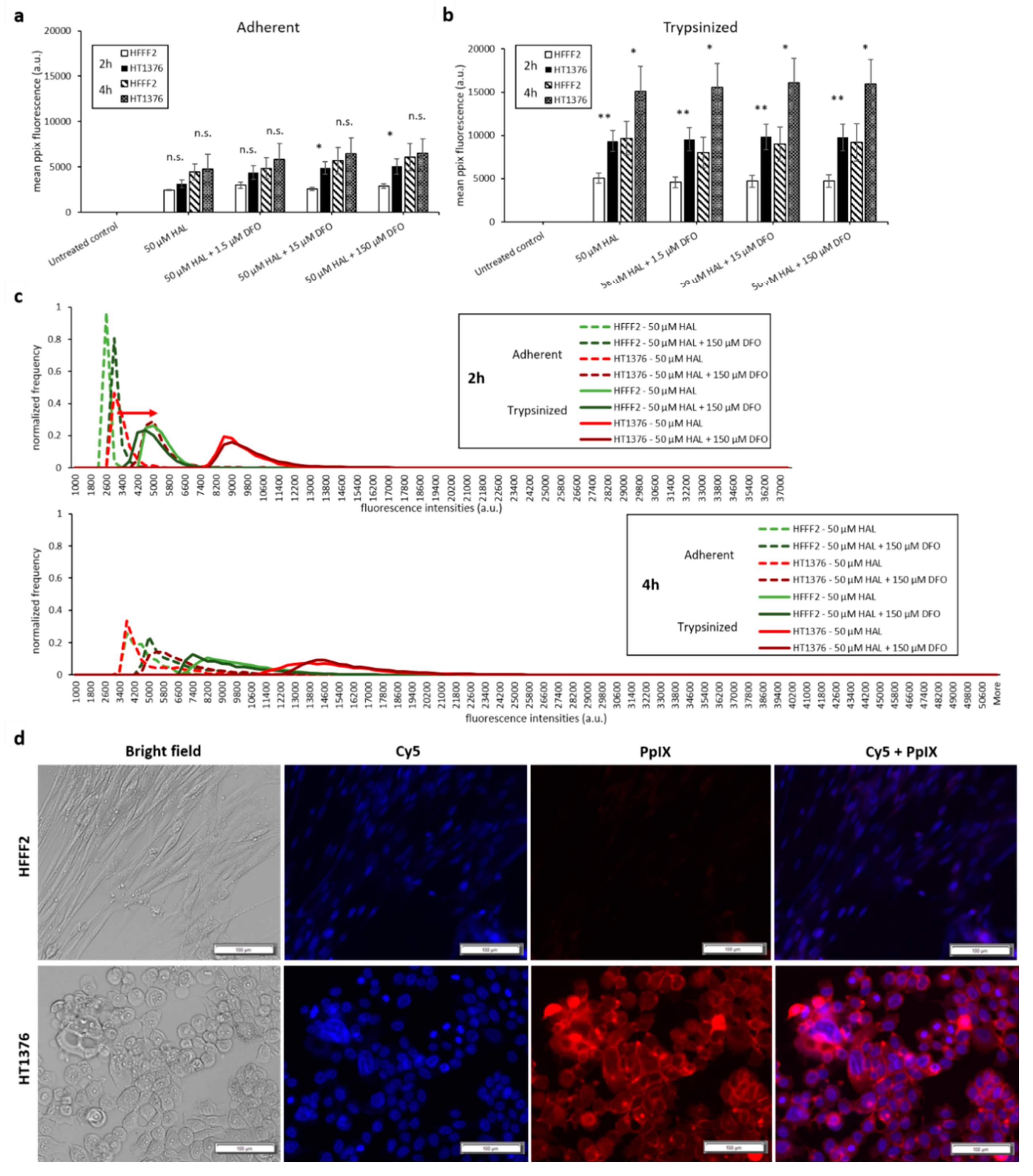
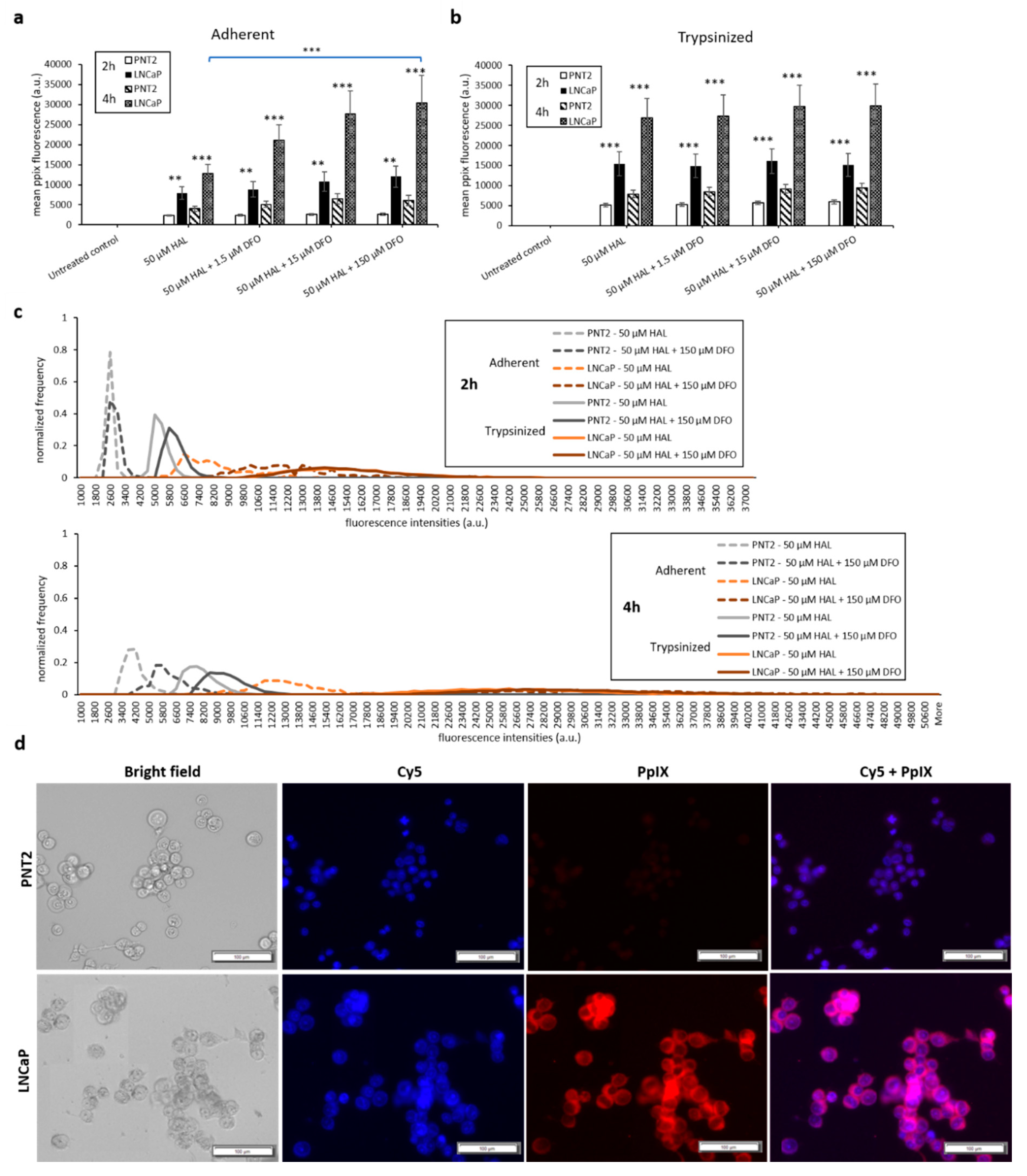
2.2. DFO
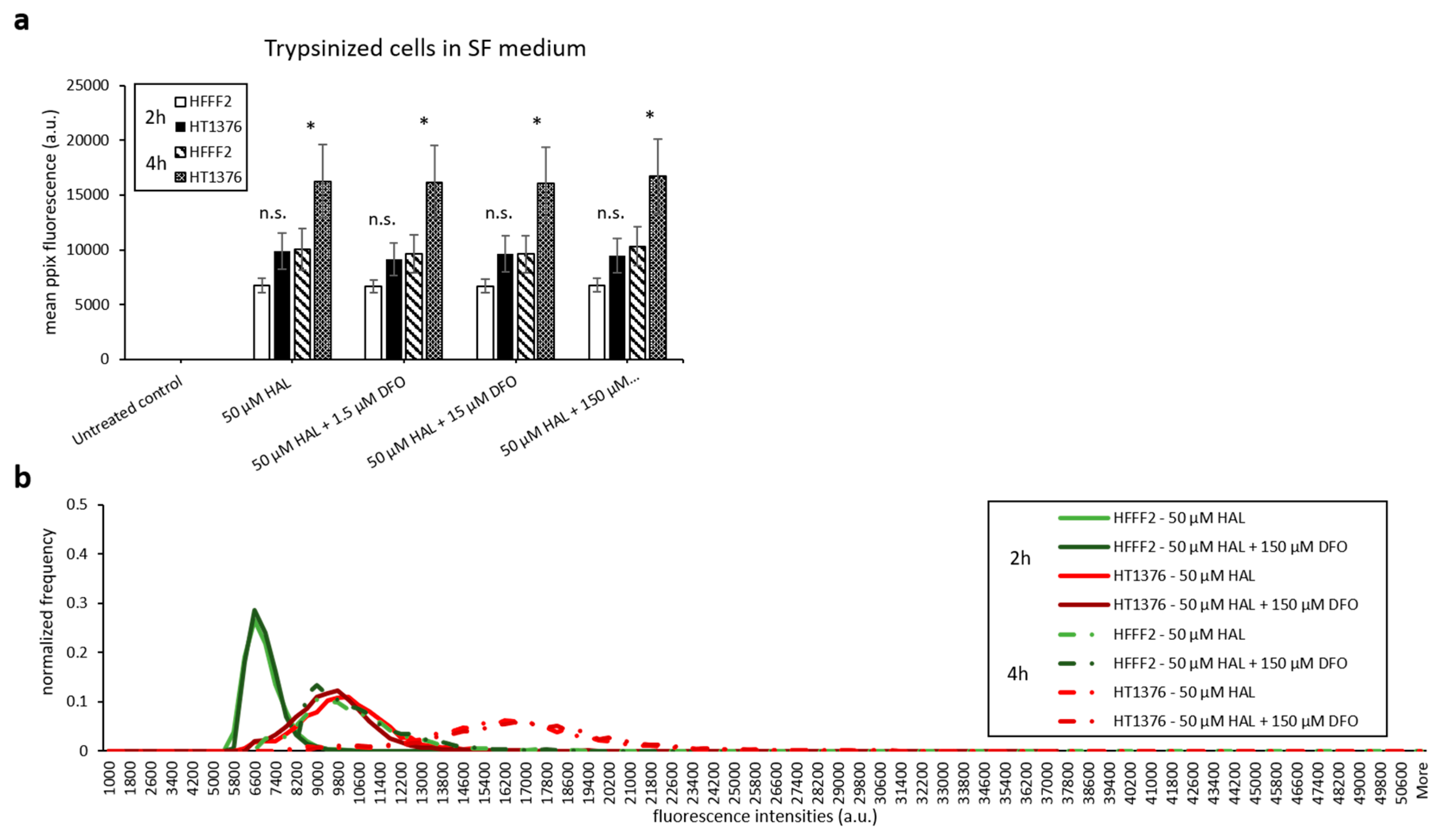
2.3. Serum-Free Media and Scraping
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines
4.3. HAL Based Treatment with DMSO
4.4. HAL Based Treatment with DFO
4.5. Nuclear Red
4.6. PpIX Fluorescence Measurement
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wachowska, M.; Muchowicz, A.; Firczuk, M.; Gabrysiak, M.; Winiarska, M.; Wańczyk, M.; Bojarczuk, K.; Golab, J. Aminolevulinic Acid (ALA) as a Prodrug in Photodynamic Therapy of Cancer. Molecules 2011, 16, 4140. [Google Scholar] [CrossRef] [Green Version]
- Ponka, P. Cell Biology of Heme. Am. J. Med Sci. 1999, 318, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Karl, A.; Tritschler, S.; Stanislaus, P.; Gratzke, C.; Tilki, D.; Strittmatter, F.; Knuchel, R.; Stief, C.G.; Zaak, D. Positive urine cytology but negative white-light cystoscopy: An indication for fluorescence cystoscopy? BJU Int. 2009, 103, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Valdes, P.A.; Bekelis, K.; Harris, B.T.; Wilson, B.C.; Leblond, F.; Kim, A.; Simmons, N.E.; Erkmen, K.; Paulsen, K.D.; Roberts, D.W. 5-Aminolevulinic acid-induced protoporphyrin IX fluorescence in meningioma: Qualitative and quantitative measurements in vivo. Neurosurgery 2014, 10 (Suppl. 1), 74–82; Discussion 82–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eljamel, S. 5-ALA Fluorescence Image Guided Resection of Glioblastoma Multiforme: A Meta-Analysis of the Literature. Int. J. Mol. Sci. 2015, 16, 10443–10456. [Google Scholar] [CrossRef] [Green Version]
- Hillemanns, P.; Wimberger, P.; Reif, J.; Stepp, H.; Klapdor, R. Photodynamic diagnosis with 5-aminolevulinic acid for intraoperative detection of peritoneal metastases of ovarian cancer: A feasibility and dose finding study. Lasers Surg. Med. 2017, 49, 169–176. [Google Scholar] [CrossRef]
- Harada, K.; Harada, Y.; Beika, M.; Koizumi, N.; Inoue, K.; Murayama, Y.; Kuriu, Y.; Nakanishi, M.; Minamikawa, T.; Yamaoka, Y.; et al. Detection of lymph node metastases in human colorectal cancer by using 5-aminolevulinic acid-induced protoporphyrin IX fluorescence with spectral unmixing. Int. J. Mol. Sci. 2013, 14, 23140–23152. [Google Scholar] [CrossRef] [Green Version]
- Ladner, D.P.; Steiner, R.A.; Allemann, J.; Haller, U.; Walt, H. Photodynamic diagnosis of breast tumours after oral application of aminolevulinic acid. Br. J. Cancer 2001, 84, 33. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Anai, S.; Fujimoto, K.; Hirao, Y.; Furuse, H.; Kai, F.; Ozono, S.; Hara, T.; Matsuyama, H.; Oyama, M.; et al. Oral 5-aminolevulinic acid mediated photodynamic diagnosis using fluorescence cystoscopy for non-muscle-invasive bladder cancer: A randomized, double-blind, multicentre phase II/III study. Photodiagnosis Photodyn. Ther. 2015, 12, 193–200. [Google Scholar] [CrossRef]
- McNicholas, K.; MacGregor, M.N.; Gleadle, J.M. In order for the light to shine so brightly, the darkness must be present-why do cancers fluoresce with 5-aminolaevulinic acid? Br. J. Cancer 2019. [Google Scholar] [CrossRef]
- Nakai, Y.; Tatsumi, Y.; Miyake, M.; Anai, S.; Kuwada, M.; Onishi, S.; Chihara, Y.; Tanaka, N.; Hirao, Y.; Fujimoto, K. Expression of ferrochelatase has a strong correlation in protoporphyrin IX accumulation with photodynamic detection of bladder cancer. Photodiagnosis Photodyn. Ther. 2016, 13 (Suppl. C), 225–232. [Google Scholar] [CrossRef] [PubMed]
- Krieg, R.C.; Fickweiler, S.; Wolfbeis, O.S.; Knuechel, R. Cell-type specific protoporphyrin IX metabolism in human bladder cancer in vitro. Photochem. Photobiol. 2000, 72, 226–233. [Google Scholar] [CrossRef]
- Anayo, L.; Magnussen, A.; Perry, A.; Wood, M.; Curnow, A. An experimental investigation of a novel iron chelating protoporphyrin IX prodrug for the enhancement of photodynamic therapy. Lasers Surg. Med. 2018, 50, 52–565. [Google Scholar] [CrossRef] [PubMed]
- Mateasik, A.; Trnka, M.; Kajo, K.; Vallova, M.; Cunderlikova, B. Cell-type dependent response to photodynamic treatment in 3D collagen cell cultures. J. Photochem. Photobiol. B Biol. 2017, 166, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Ozawa, T.; Mizuno, F.; Onishi, S.; Owari, T.; Hori, S.; Morizawa, Y.; Tatsumi, Y.; Miyake, M.; Tanaka, N.; et al. Spectrophotometric photodynamic detection involving extracorporeal treatment with hexaminolevulinate for bladder cancer cells in voided urine. J. Cancer Res. Clin. Oncol. 2017, 143, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Miyake, M.; Anai, S.; Hori, S.; Tatsumi, Y.; Morizawa, Y.; Onisi, S.; Tanaka, N.; Fujimoto, K. Spectrophotometric photodynamic diagnosis of prostate cancer cells excreted in voided urine using 5-aminolevulinic acid. Lasers Med. Sci. 2018. [Google Scholar] [CrossRef]
- Fu, C.Y.; Ng, B.K.; Razul, S.G.; Chin, W.W.; Tan, P.H.; Lau, W.K.; Olivo, M. Fluorescence detection of bladder cancer using urine cytology. Int. J. Oncol. 2007, 31, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Čunderlíková, B.; Wahlqvist, R.; Berner, A.; Vasovič, V.; Warloe, T.; Nesland, J.M.; Peng, Q. Detection of urinary bladder cancer with flow cytometry and hexaminolevulinate in urine samples. Cytopathology 2007, 18, 87–95. [Google Scholar] [CrossRef]
- Yamamichi, G.; Nakata, W.; Tani, M.; Tsujimura, G.; Tsujimoto, Y.; Nin, M.; Mimura, A.; Miwa, H.; Tsujihata, M. High diagnostic efficacy of 5-aminolevulinic acid-induced fluorescent urine cytology for urothelial carcinoma. Int. J. Clin. Oncol. 2019, 24, 1075–1080. [Google Scholar] [CrossRef]
- Macgregor-Ramiasa, M.; McNicholas, K.; Ostrikov, K.; Li, J.; Michael, M.; Gleadle, J.M.; Vasilev, K. A platform for selective immuno-capture of cancer cells from urine. Biosens. Bioelectron. 2017. [Google Scholar] [CrossRef]
- Yang, X.; Palasuberniam, P.; Kraus, D.; Chen, B. Aminolevulinic Acid-Based Tumor Detection and Therapy: Molecular Mechanisms and Strategies for Enhancement. Int. J. Mol. Sci. 2015, 16, 5865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiya, Y.; Fukuhara, H.; Matsumoto, K.; Endo, Y.; Nakajima, M.; Tanaka, T.; Okura, I.; Kurabayashi, A.; Furihata, M.; Inoue, K.; et al. Expression levels of PEPT1 and ABCG2 play key roles in 5-aminolevulinic acid (ALA)-induced tumor-specific protoporphyrin IX (PpIX) accumulation in bladder cancer. Photodiagnosis Photodyn. Ther. 2013, 10, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.; de Bruijn, H.S.; Di Venosa, G.; Mamone, L.; Robinson, D.J.; Juarranz, A.; Batlle, A.; Casas, A. Porphyrin synthesis from aminolevulinic acid esters in endothelial cells and its role in photodynamic therapy. J. Photochem. Photobiol. B Biol. 2009, 96, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xia, Y.; Liu, X.; Jiang, S.; Xiong, L. Desferrioxamine shows different potentials for enhancing 5-aminolaevulinic acid-based photodynamic therapy in several cutaneous cell lines. Lasers Med. Sci. 2010, 25, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Curnow, A.; Perry, A.; Wood, M. Improving in vitro photodynamic therapy through the development of a novel iron chelating aminolaevulinic acid prodrug. Photodiagnosis Photodyn. Ther. 2019, 25, 157–165. [Google Scholar] [CrossRef]
- Chan, K.M.; Vasilev, K.; Shirazi, H.S.; McNicholas, K.; Li, J.; Gleadle, J.; Macgregor, M. Biosensor device for the photo-specific detection of immuno-captured bladder cancer cells using hexaminolevulinate: An ex-vivo study. Photodiagnosis Photodyn. Ther. 2019. [Google Scholar] [CrossRef]
- Lotan, Y.; Bivalacqua, T.J.; Downs, T.; Huang, W.; Jones, J.; Kamat, A.M.; Konety, B.; Malmström, P.-U.; McKiernan, J.; O’Donnell, M.; et al. Blue light flexible cystoscopy with hexaminolevulinate in non-muscle-invasive bladder cancer: Review of the clinical evidence and consensus statement on optimal use in the USA—update 2018. Nat. Rev. Urol. 2019, 16, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Notman, R.; Noro, M.; O′Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef]
- De Rosa, F.S.; Marchetti, J.M.; Thomazini, J.A.; Tedesco, A.C.; Bentley, M.V.L.B. A vehicle for photodynamic therapy of skin cancer: Influence of dimethylsulphoxide on 5-aminolevulinic acid in vitro cutaneous permeation and in vivo protoporphyrin IX accumulation determined by confocal microscopy. J. Control. Release 2000, 65, 359–366. [Google Scholar] [CrossRef]
- Conder, L.H.; Woodard, S.I.; Dailey, H.A. Multiple mechanisms for the regulation of haem synthesis during erythroid cell differentiation. Possible role for coproporphyrinogen oxidase. Biochem. J. 1991, 275 Pt 2, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.; Anholt, H.; Bech, O.; Moan, J. The influence of iron chelators on the accumulation of protoporphyrin IX in 5-aminolaevulinic acid-treated cells. Br. J. Cancer 1996, 74, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, P.A.; Samkoe, K.; O′Hara, J.A.; Roberts, D.W.; Paulsen, K.D.; Pogue, B.W. Deferoxamine iron chelation increases delta-aminolevulinic acid induced protoporphyrin IX in xenograft glioma model. Photochem. Photobiol. 2010, 86, 471–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uekusa, M.; Omura, K.; Nakajima, Y.; Hasegawa, S.; Harada, H.; Morita, K.I.; Tsuda, H. Uptake and kinetics of 5-aminolevulinic acid in oral squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2010, 39, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Juzenas, P.; Juzeniene, A.; Moan, J. Deferoxamine photosensitizes cancer cells in vitro. Biochem. Biophys. Res. Commun. 2005, 332, 388–391. [Google Scholar] [CrossRef]
- de Souza, A.L.; Marra, K.; Gunn, J.; Samkoe, K.S.; Kanick, S.C.; Davis, S.C.; Chapman, M.S.; Maytin, E.V.; Hasan, T.; Pogue, B.W. Comparing desferrioxamine and light fractionation enhancement of ALA-PpIX photodynamic therapy in skin cancer. Br. J. Cancer 2016, 115, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Palasuberniam, P.; Kraus, D.; Mansi, M.; Braun, A.; Howley, R.; Myers, K.A.; Chen, B. Ferrochelatase Deficiency Abrogated the Enhancement of Aminolevulinic Acid-mediated Protoporphyrin IX by Iron Chelator Deferoxamine. Photochem. Photobiol. 2019. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Cell Line | 50 µM HAL (Alone) | 50 µM HAL + 0.05 µM DMSO | 50 µM HAL + 0.25 µM DMSO | 50 µM HAL + 0.5 µM DMSO |
|---|---|---|---|---|---|
| Adherent monolayer | HFFF2 | – | – | – | – |
| HT1376 | – | ↑ | – | – | |
| PNT2 | – | – | – | – | |
| LNCaP | – | ↓ | ↑ | – | |
| Trypsinised cells in PBS | HFFF2 | – | – | – | – |
| HT1376 | – | – | – | – | |
| PNT2 | – | – | – | – | |
| LNCaP | – | ↓ | – | – |
| 2 h | 4 h | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Condition | Cell Line | 50 µM HAL (Alone) | 50 µM HAL + 1.5 µM DFO | 50 µM HAL + 15 µM DFO | 50 µM HAL + 150 µM DFO | 50 µM HAL (Alone) | 50 µM HAL + 1.5 µM DFO | 50 µM HAL + 15 µM DFO | 50 µM HAL + 150 µM DFO |
| Adherent monolayer | HFFF2 | – | ↑ | – | ↓ | – | – | ↑ | ↑ |
| HT1376 | – | ↑ | ↑ | ↑ | – | ↑ | ↑ | ↑ | |
| PNT2 | – | – | – | – | – | ↑ | ↑ | ↑ | |
| LNCaP | – | ↑ | ↑ | ↑ | – | ↑ | ↑ | ↑ | |
| Trypsinised cells in PBS | HFFF2 | – | ↓ | – | – | – | ↓ | ↓ | – |
| HT1376 | – | – | – | – | – | – | ↑ | ↑ | |
| PNT2 | – | – | ↑ | ↑ | – | ↑ | ↑ | ↑ | |
| LNCaP | – | – | ↑ | – | – | – | ↑ | ↑ | |
| Tryspinised cells in serum-free media | PNT2 | – | – | – | ↑ | – | ↑ | – | ↑ |
| LNCaP | – | – | ↑ | ↑ | – | – | ↑ | ↑ | |
| Scrapped | PNT2 | – | ↓ | – | – | – | – | – | ↑ |
| LNCaP | – | – | ↑ | ↑ | – | ↓ | ↓ | – | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, K.M.; Gleadle, J.; Vasilev, K.; MacGregor, M. Probing Hexaminolevulinate Mediated PpIX Fluorescence in Cancer Cell Suspensions in the Presence of Chemical Adjuvants. Int. J. Mol. Sci. 2020, 21, 2963. https://doi.org/10.3390/ijms21082963
Chan KM, Gleadle J, Vasilev K, MacGregor M. Probing Hexaminolevulinate Mediated PpIX Fluorescence in Cancer Cell Suspensions in the Presence of Chemical Adjuvants. International Journal of Molecular Sciences. 2020; 21(8):2963. https://doi.org/10.3390/ijms21082963
Chicago/Turabian StyleChan, Kit Man, Jonathan Gleadle, Krasimir Vasilev, and Melanie MacGregor. 2020. "Probing Hexaminolevulinate Mediated PpIX Fluorescence in Cancer Cell Suspensions in the Presence of Chemical Adjuvants" International Journal of Molecular Sciences 21, no. 8: 2963. https://doi.org/10.3390/ijms21082963
APA StyleChan, K. M., Gleadle, J., Vasilev, K., & MacGregor, M. (2020). Probing Hexaminolevulinate Mediated PpIX Fluorescence in Cancer Cell Suspensions in the Presence of Chemical Adjuvants. International Journal of Molecular Sciences, 21(8), 2963. https://doi.org/10.3390/ijms21082963