High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer

 ,
,  , ,
, , 


Abstract
:1. Introduction
2. Results
2.1. Clinicopathological Features Were Not Significantly Different between miR-34a High and miR-34a Low Expressing Tumors Except for HER2 Expression
2.2. Nottingham Pathological Grade Was Associated with miR-34a Expression and miR-34a High Expressing Tumors Were Associated with Lower Expression of MKI67 in METABRIC
2.3. High MiR-34a Breast Cancer Associated with Lower Gene Set Variation Analysis (GSVA) Scores of the Gene Sets Related with Cell Cycle or Cell Proliferation
2.4. High MiR-34a Expression Tumors Enriched p53 Pathway and Apoptosis Gene Sets and Was Associated with Expression of Apoptosis Related Genes
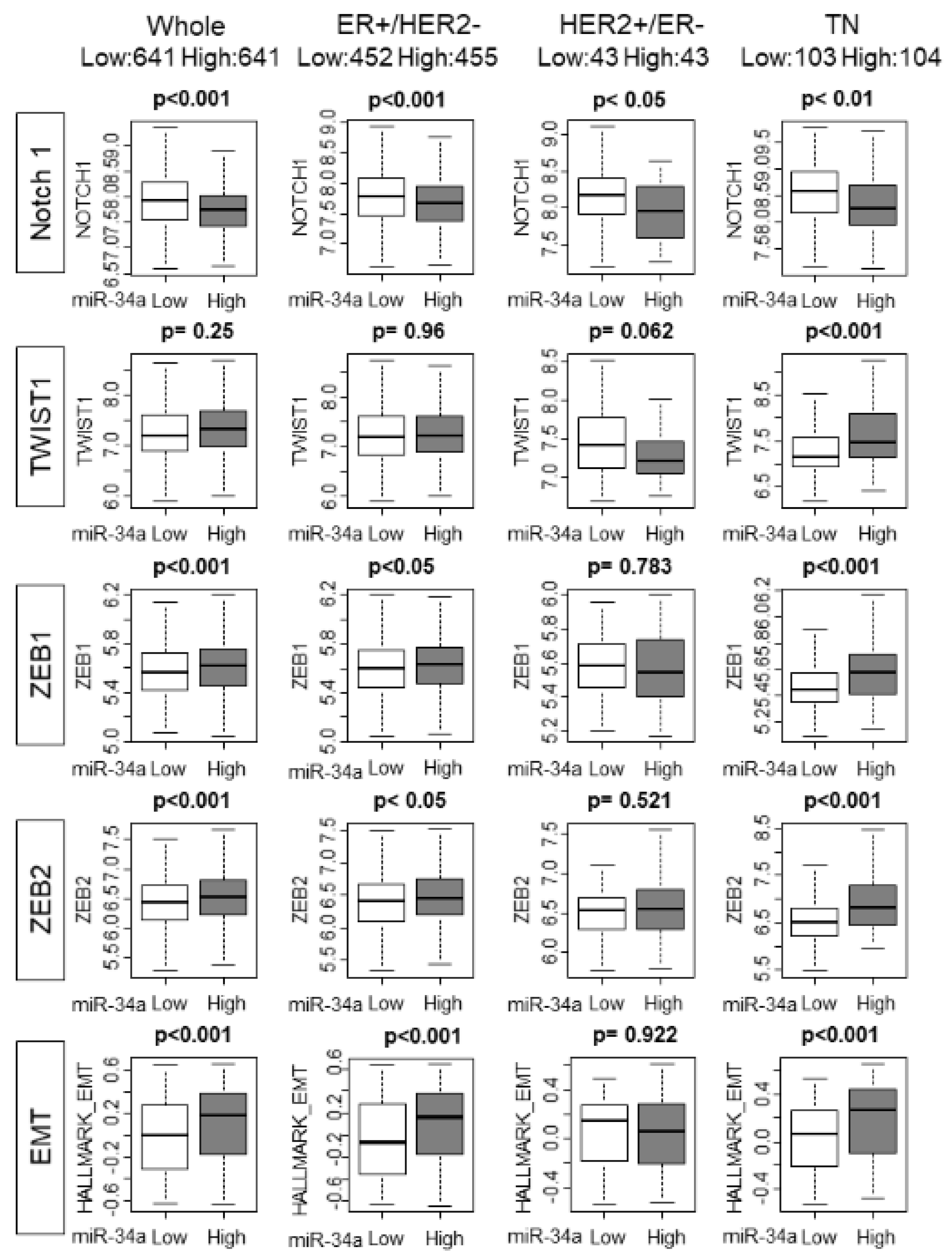
2.5. High MiR-34a Expression Tumors Associated with Higher Epithelial–Mesenchymal Transition (EMT) Score
2.6. High miR-34a Expression Tumor Was Not Associated with Distant Metastasis and Was Not Associated with Better Survival Outcome
3. Discussion
4. Material and Methods
4.1. Obtaining Data of TCGA and METABRIC
4.2. Gene Set Variation Analysis (GSVA) and Gene Set Enrichment Analysis (GSEA)
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ER | Estrogen receptor |
| HER2 | Human epidermal growth factor receptor 2 |
| TN | Triple-negative |
| TCGA | The Cancer Genome Atlas |
| METABRIC | Molecular Taxonomy of Breast Cancer International Consortium |
| GSEA | Gene Set Enrichment Analysis |
| GSVA | Gene Set Variant Analysis |
| BCL2 | B cell lymphoma 2 |
| ANXA5 | Annexin A5 |
| CASP8 | Caspase 8 |
| CASP9 | Caspase 9 |
| ZEB1 | Zinc finger E-box binding homeobox 1 |
| ZEB2 | Zinc finger E-box binding homeobox 2 |
| TWIST1 | Twist family bHLH transcription factor 1 |
| EMT | Epithelial-mesenchymal transition |
References
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Miller, K.D.; Goding Sauer, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Joyce, D.; Takabe, K. Current status and limitations of immunotherapy for breast cancer. Surgery 2020, 167, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Giuliano, M.; Trivedi, M.V.; Schiff, R.; Osborne, C.K. Metastasis dormancy in estrogen receptor-positive breast cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 6389–6397. [Google Scholar] [CrossRef] [Green Version]
- Tokumaru, Y.; Takabe, K.; Yoshida, K.; Akao, Y. Effects of MIR143 on RAS signaling networks in solid tumors: A brief overview. Cancer Sci. 2020, 10.1111/cas.14357. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Tajirika, T.; Sugito, N.; Kuranaga, Y.; Shinohara, H.; Tsujino, T.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K. Synthetic miR-143 Inhibits Growth of HER2−Positive Gastric Cancer Cells by Suppressing KRAS Networks Including DDX6 RNA Helicase. Int. J. Mol. Sci. 2019, 20, 1697. [Google Scholar] [CrossRef] [Green Version]
- Sporn, J.C.; Katsuta, E.; Yan, L.; Takabe, K. Expression of MicroRNA-9 is Associated With Overall Survival in Breast Cancer Patients. J. Surg. Res. 2019, 233, 426–435. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kawaguchi, T.; Yan, L.; Young, J.; Qi, Q.; Takabe, K. Clinical Relevance of microRNA Expressions in Breast Cancer Validated Using the Cancer Genome Atlas (TCGA). Ann. Surg. Oncol. 2017, 24, 2943–2949. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Yan, L.; Qi, Q.; Peng, X.; Gabriel, E.M.; Young, J.; Liu, S.; Takabe, K. Overexpression of suppressive microRNAs, miR-30a and miR-200c are associated with improved survival of breast cancer patients. Sci. Rep. 2017, 7, 15945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.; Kawaguchi, T.; Yan, L.; Qi, Q.; Liu, S.; Takabe, K. Tamoxifen sensitivity-related microRNA-342 is a useful biomarker for breast cancer survival. Oncotarget 2017, 8, 99978–99989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, T.; Yan, L.; Qi, Q.; Peng, X.; Edge, S.B.; Young, J.; Yao, S.; Liu, S.; Otsuji, E.; Takabe, K. Novel MicroRNA-Based Risk Score Identified by Integrated Analyses to Predict Metastasis and Poor Prognosis in Breast Cancer. Ann. Surg. Oncol. 2018, 25, 4037–4046. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 family: A potential tumor suppressor and therapeutic candidate in cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misso, G.; Di Martino, M.T.; De Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. Mir-34: A new weapon against cancer? Mol. Ther. Nucleic Acids 2014, 3, e194. [Google Scholar] [CrossRef] [PubMed]
- Raver-Shapira, N.; Marciano, E.; Meiri, E.; Spector, Y.; Rosenfeld, N.; Moskovits, N.; Bentwich, Z.; Oren, M. Transcriptional activation of miR-34a contributes to p53-mediated apoptosis. Mol. Cell 2007, 26, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.D.; Parsons, C.; Slack, F.J. The tumor-suppressive and potential therapeutic functions of miR-34a in epithelial carcinomas. Expert Opin. Ther. Targets 2016, 20, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Ahmad, A.; Sarkar, F.H. The role of microRNAs in breast cancer migration, invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 13414–13437. [Google Scholar] [CrossRef] [Green Version]
- Imani, S.; Wu, R.C.; Fu, J. MicroRNA-34 family in breast cancer: From research to therapeutic potential. J. Cancer 2018, 9, 3765–3775. [Google Scholar] [CrossRef] [Green Version]
- Tokumaru, Y.; Oshi, M.; Katsuta, E.; Yan, L.; Satyananda, V.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; Takabe, K. KRAS signaling enriched triple negative breast cancer is associated with favorable tumor immune microenvironment and better survival. Am. J. Cancer Res. 2020, 10, 897–907. [Google Scholar]
- Ungerleider, N.A.; Rao, S.G.; Shahbandi, A.; Yee, D.; Niu, T.; Frey, W.D.; Jackson, J.G. Breast cancer survival predicted by TP53 mutation status differs markedly depending on treatment. Breast Cancer Res. 2018, 20, 115. [Google Scholar] [CrossRef]
- Javeri, A.; Ghaffarpour, M.; Taha, M.F.; Houshmand, M. Downregulation of miR-34a in breast tumors is not associated with either p53 mutations or promoter hypermethylation while it correlates with metastasis. Med. Oncol. 2013, 30, 413. [Google Scholar] [CrossRef]
- Kim, W.J.; Beardsley, D.I.; Adamson, A.W.; Brown, K.D. The monofunctional alkylating agent N-methyl-N’-nitro-N-nitrosoguanidine triggers apoptosis through p53-dependent and -independent pathways. Toxicol. Appl. Pharmacol. 2005, 202, 84–98. [Google Scholar] [CrossRef]
- Peurala, H.; Greco, D.; Heikkinen, T.; Kaur, S.; Bartkova, J.; Jamshidi, M.; Aittomäki, K.; Heikkilä, P.; Bartek, J.; Blomqvist, C.; et al. MiR-34a expression has an effect for lower risk of metastasis and associates with expression patterns predicting clinical outcome in breast cancer. PLoS ONE 2011, 6, e26122. [Google Scholar] [CrossRef]
- Chen, J.; Chang, H.; Peng, X.; Gu, Y.; Yi, L.; Zhang, Q.; Zhu, J.; Mi, M. 3,6-dihydroxyflavone suppresses the epithelial-mesenchymal transition in breast cancer cells by inhibiting the Notch signaling pathway. Sci. Rep. 2016, 6, 28858. [Google Scholar] [CrossRef] [PubMed]
- Imani, S.; Wei, C.; Cheng, J.; Khan, M.A.; Fu, S.; Yang, L.; Tania, M.; Zhang, X.; Xiao, X.; Zhang, X.; et al. MicroRNA-34a targets epithelial to mesenchymal transition-inducing transcription factors (EMT-TFs) and inhibits breast cancer cell migration and invasion. Oncotarget 2017, 8, 21362–21379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.T.; Zhong, H.T.; Li, G.W.; Shen, J.X.; Ye, Q.Q.; Zhang, M.L.; Liu, J. Oncogenic functions of the EMT-related transcription factor ZEB1 in breast cancer. J. Transl. Med. 2020, 18, 51. [Google Scholar] [CrossRef]
- Adams, B.D.; Wali, V.B.; Cheng, C.J.; Inukai, S.; Booth, C.J.; Agarwal, S.; Rimm, D.L.; Győrffy, B.; Santarpia, L.; Pusztai, L.; et al. miR-34a Silences c-SRC to Attenuate Tumor Growth in Triple-Negative Breast Cancer. Cancer Res. 2016, 76, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, S.; Hanna, J.; Sherman, M.E.; Figueroa, J.; Rimm, D.L. Quantitative assessment of miR34a as an independent prognostic marker in breast cancer. Br. J. Cancer 2015, 112, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayraktar, R.; Ivan, C.; Bayraktar, E.; Kanlikilicer, P.; Kabil, N.N.; Kahraman, N.; Mokhlis, H.A.; Karakas, D.; Rodriguez-Aguayo, C.; Arslan, A.; et al. Dual Suppressive Effect of miR-34a on the FOXM1/eEF2-Kinase Axis Regulates Triple-Negative Breast Cancer Growth and Invasion. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 4225–4241. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Lee, J.; Kang, J.; Kim, S.H.; Yoo, T.K.; Oh, S.; Lee, A. Notch1 in Tumor Microvascular Endothelial Cells and Tumoral miR-34a as Prognostic Markers in Locally Advanced Triple-Negative Breast Cancer. J. Breast Cancer 2019, 22, 562–578. [Google Scholar] [CrossRef]
- Takeshita, T.; Asaoka, M.; Katsuta, E.; Photiadis, S.J.; Narayanan, S.; Yan, L.; Takabe, K. High expression of polo-like kinase 1 is associated with TP53 inactivation, DNA repair deficiency, and worse prognosis in ER positive Her2 negative breast cancer. Am. J. Transl. Res. 2019, 11, 6507–6521. [Google Scholar]
- Takahashi, H.; Asaoka, M.; Yan, L.; Rashid, O.M.; Oshi, M.; Ishikawa, T.; Nagahashi, M.; Takabe, K. Biologically Aggressive Phenotype and Anti-cancer Immunity Counterbalance in Breast Cancer with High Mutation Rate. Sci. Rep. 2020, 10, 1852. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsuta, E.; Yan, L.; Takeshita, T.; McDonald, K.-A.; Dasgupta, S.; Opyrchal, M.; Takabe, K. High MYC mRNA Expression Is More Clinically Relevant than MYC DNA Amplification in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2019, 21, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, M.; Oshi, M.; Butash, A.L.; Asaoka, M.; Katsuta, E.; Peng, X.; Qi, Q.; Yan, L.; Takabe, K. Estrogen Receptor Positive Breast Cancer with High Expression of Androgen Receptor has Less Cytolytic Activity and Worse Response to Neoadjuvant Chemotherapy but Better Survival. Int. J. Mol. Sci. 2019, 20, 2655. [Google Scholar] [CrossRef] [Green Version]
- Okano, M.; Oshi, M.; Butash, A.L.; Katsuta, E.; Tachibana, K.; Saito, K.; Okayama, H.; Peng, X.; Yan, L.; Kono, K.; et al. Triple-Negative Breast Cancer with High Levels of Annexin A1 Expression Is Associated with Mast Cell Infiltration, Inflammation, and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshita, T.; Yan, L.; Asaoka, M.; Rashid, O.; Takabe, K. Late recurrence of breast cancer is associated with pro-cancerous immune microenvironment in the primary tumor. Sci. Rep. 2019, 9, 16942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. Bioinformatics 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Katsuta, E.; Maawy, A.A.; Yan, L.; Takabe, K. High expression of bone morphogenetic protein (BMP) 6 and BMP7 are associated with higher immune cell infiltration and better survival in estrogen receptor--positive breast cancer. Oncol. Rep. 2019, 42, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Kawaguchi, T.; Yan, L.; Peng, X.; Qi, Q.; Takabe, K. Cytolytic Activity Score to Assess Anticancer Immunity in Colorectal Cancer. Ann. Surg. Oncol. 2018, 25, 2323–2331. [Google Scholar] [CrossRef]
- Terakawa, T.; Katsuta, E.; Yan, L.; Turaga, N.; McDonald, K.-A.; Fujisawa, M.; Guru, K.A.; Takabe, K. High expression of SLCO2B1 is associated with prostate cancer recurrence after radical prostatectomy. Oncotarget 2018, 9, 14207–14218. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Factor | Whole Cohort (n = 1282) | p Value | |
|---|---|---|---|
| miR-34a High n = 641 | miR-34a Low n = 641 | ||
| Age | |||
| <65 y | 259 | 252 | 0.732 |
| ≥65 y | 382 | 389 | |
| Stage | |||
| 0 | 5 | 6 | 0.068 |
| 1 | 216 | 168 | |
| 2 | 296 | 310 | |
| 3 | 54 | 46 | |
| 4 | 8 | 2 | |
| Unknown | 62 | 109 | |
| Tumor Size | |||
| ≤2 cm | 48 | 47 | 0.621 |
| 2–5 cm | 300 | 314 | |
| >5 cm | 285 | 266 | |
| Unknown | 8 | 14 | |
| Lymph Node Metastasis | |||
| Negative | 319 | 338 | 0.954 |
| Positive | 276 | 289 | |
| Unknown | 46 | 14 | |
| ER Status | |||
| Negative | 141 | 159 | 0.262 |
| Positive | 500 | 482 | |
| PgR Status | |||
| Negative | 292 | 315 | 0.218 |
| Positive | 349 | 326 | |
| HER2 Status | |||
| Negative | 548 | 573 | 0.043 |
| Positive | 93 | 68 | |
| Histologic Subtype | |||
| Invasive Ductal Carcinoma | 481 | 502 | 0.254 |
| Invasive Lobular Carcinoma | 54 | 48 | |
| Mixed Ductal and Lobular Carcinoma | 75 | 72 | |
| Other | 11 | 4 | |
| Unknown | 20 | 15 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokumaru, Y.; Katsuta, E.; Oshi, M.; Sporn, J.C.; Yan, L.; Le, L.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; et al. High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 3045. https://doi.org/10.3390/ijms21093045
Tokumaru Y, Katsuta E, Oshi M, Sporn JC, Yan L, Le L, Matsuhashi N, Futamura M, Akao Y, Yoshida K, et al. High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. International Journal of Molecular Sciences. 2020; 21(9):3045. https://doi.org/10.3390/ijms21093045
Chicago/Turabian StyleTokumaru, Yoshihisa, Eriko Katsuta, Masanori Oshi, Judith C. Sporn, Li Yan, Lan Le, Nobuhisa Matsuhashi, Manabu Futamura, Yukihiro Akao, Kazuhiro Yoshida, and et al. 2020. "High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer" International Journal of Molecular Sciences 21, no. 9: 3045. https://doi.org/10.3390/ijms21093045
APA StyleTokumaru, Y., Katsuta, E., Oshi, M., Sporn, J. C., Yan, L., Le, L., Matsuhashi, N., Futamura, M., Akao, Y., Yoshida, K., & Takabe, K. (2020). High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. International Journal of Molecular Sciences, 21(9), 3045. https://doi.org/10.3390/ijms21093045