Effects of Prenatal Exposure to a Low-Dose of Bisphenol A on Sex Differences in Emotional Behavior and Central Alpha2-Adrenergic Receptor Binding



Abstract
:1. Introduction
2. Results
2.1. Preliminary Analysis
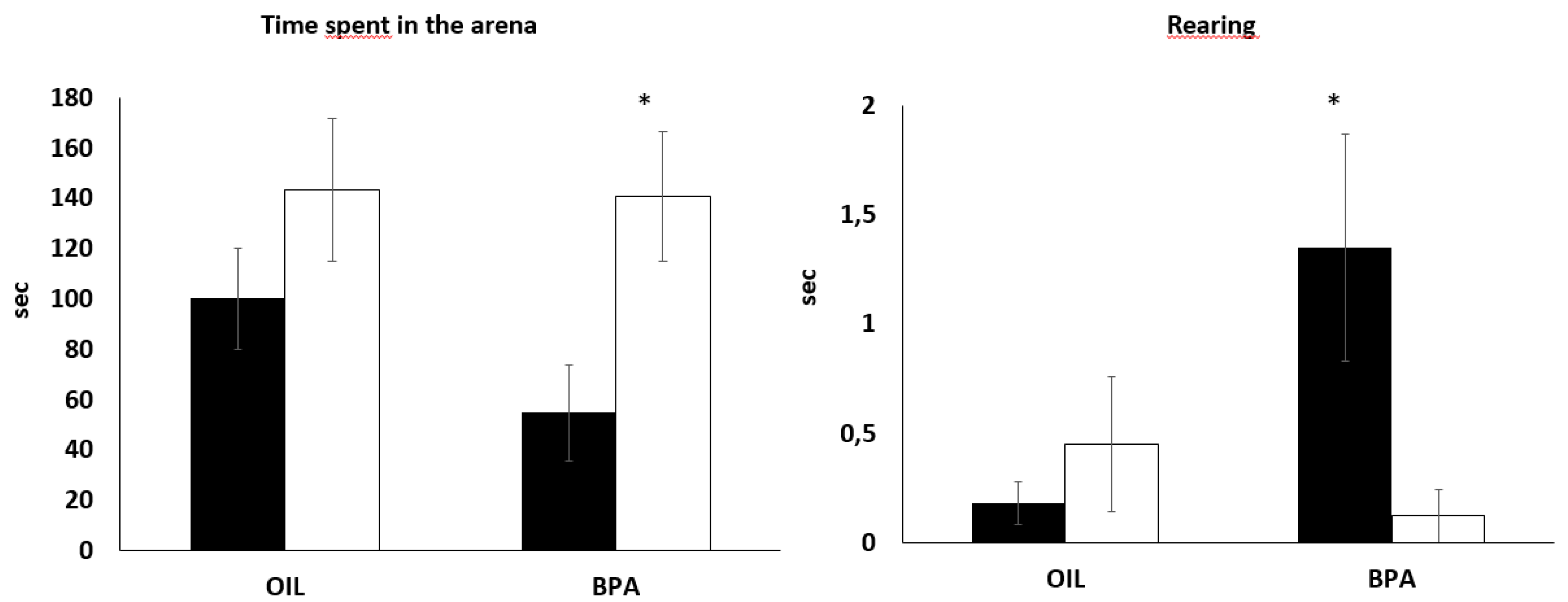
2.2. Effects of Prenatal Exposure to BPA on Behaviors during the Free-Exploratory Open Field (FOF)
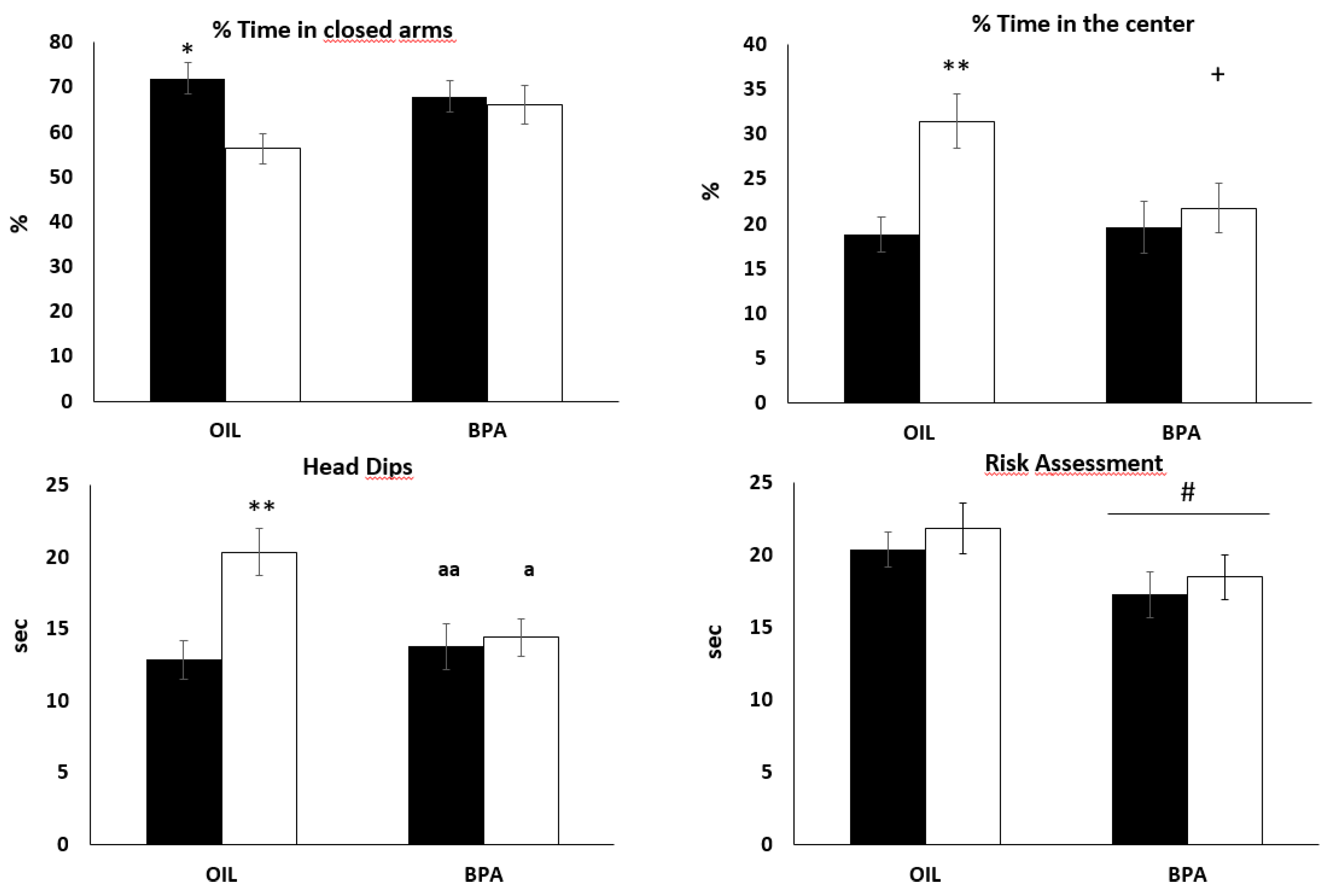
2.3. Effects of Prenatal Exposure to BPA on Behaviors in the Elevated Plus Maze (EPM)
2.4. Effects of Prenatal Exposure to BPA on Reproductive Behaviors of Males
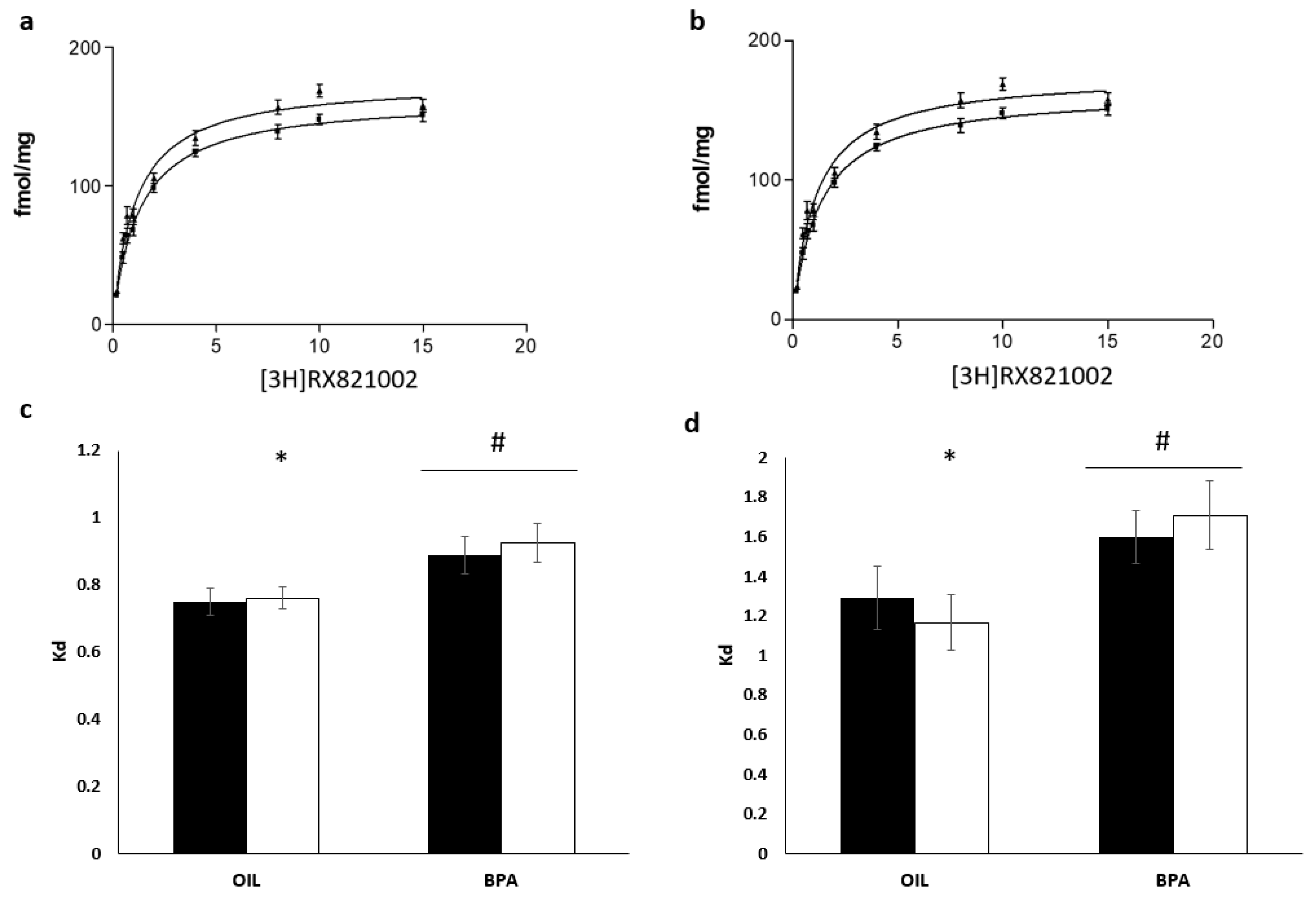
2.5. Effects of Prenatal Exposure to BPA on Receptor Density and Affinity in the Locus Coeruleus
2.6. Effects of Prenatal Exposure to BPA on Receptor Density and Affinity in the Medial Preoptic Area
3. Discussion
4. Materials and Methods
4.1. Maternal Treatment
4.2. Birth and Weaning
4.3. Free-Exploratory Open Field Test (FOF)
4.4. Elevated Plus Maze
4.5. Sociosexual Behavior
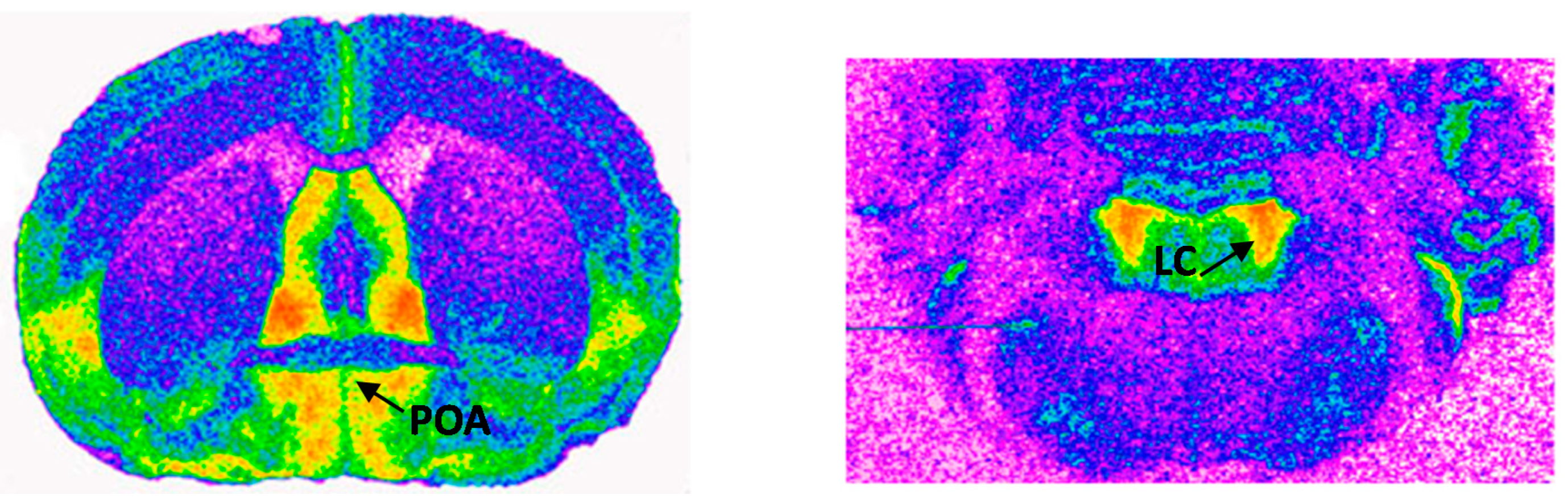
4.6. Receptor Autoradiography and Quantification
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Schönfelder, G.; Wittfoht, W.; Hopp, H.; Talsness, C.E.; Paul, M.; Chahoud, I. Parent bisphenol a accumulation in the human maternal-fetal-placental unit. Environ. Health Perspect. 2002, 110. [Google Scholar] [CrossRef] [Green Version]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 2002, 17, 2839–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Irie, M.; Kishikawa, N.; Wada, M.; Kuroda, N.; Nakashima, K. Determination of bisphenol A in human breast milk by HPLC with column-switching and fluorescence detection. Biomed. Chromatogr. 2004, 18, 501–507. [Google Scholar] [CrossRef] [PubMed]
- vom Saal, F.S.; Welshons, W.V. Evidence that bisphenol A (BPA) can be accurately measured without contamination in human serum and urine, and that BPA causes numerous hazards from multiple routes of exposure. Mol. Cell. Endocrinol. 2014, 398, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Welshons, W.V.; Nagel, S.C.; vom Saal, F.S. Large Effects from Small Exposures. III. Endocrine Mechanisms Mediating Effects of Bisphenol A at Levels of Human Exposure. Endocrinology 2006, 147, s56–s69. [Google Scholar] [CrossRef]
- Zoeller, R.T.; Bansal, R.; Parris, C. Bisphenol-A, an environmental contaminant that acts as a thyroid hormone receptor antagonist in vitro, increases serum thyroxine, and alters RC3/neurogranin expression in the developing rat brain. Endocrinology 2005, 146, 607–612. [Google Scholar] [CrossRef]
- Howdeshell, K.L.; Hotchkiss, A.K.; Thayer, K.A.; Vandenbergh, J.G.; Vom Saal, F.S. Environmental toxins: Exposure to bisphenol A advances puberty. Nature 1999, 401, 763–764. [Google Scholar] [CrossRef]
- Palanza, P.; Gioiosa, L.; vom Saal, F.S.; Parmigiani, S. Effects of developmental exposure to bisphenol A on brain and behavior in mice. Environ. Res. 2008, 108, 150–157. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hunt, P.A.; Gore, A.C. Endocrine disruptors and the future of toxicology testing—Lessons from CLARITY–BPA. Nat. Rev. Endocrinol. 2019, 15, 366–374. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Arnold, A.P. Reframing sexual differentiation of the brain. Nat. Neurosci. 2011, 14, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, A. Synaptogenic action of sex steroids in developing and adult neuroendocrine brain. Psychoneuroendocrinology 1991, 16, 25–40. [Google Scholar] [CrossRef]
- Arnold, A.P.; Gorski, R.A. Gonadal Steroid Induction of Structural Sex Differences in the Central Nervous System. Annu. Rev. Neurosci. 1984, 7, 413–442. [Google Scholar] [CrossRef] [PubMed]
- Simerly, R.B. Wired for Reproduction: Organization and Development of Sexually Dimorphic Circuits in the Mammalian Forebrain. Annu. Rev. Neurosci. 2002, 25, 507–536. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; Konkle, A.T.M.; Zup, S.L.; McCarthy, M.M. Impact of sex and hormones on new cells in the developing rat hippocampus: A novel source of sex dimorphism? Eur. J. Neurosci. 2008, 27, 791–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillamón, A.; de Blas, M.R.; Segovia, S. Effects of sex steroids on the development of the locus coeruleus in the rat. Dev. Brain Res. 1988, 40, 306–310. [Google Scholar] [CrossRef]
- Cooke, B.M.; Tabibnia, G.; Breedlove, S.M. A brain sexual dimorphism controlled by adult circulating androgens. Proc. Natl. Acad. Sci. USA 1999, 96, 7538–7540. [Google Scholar] [CrossRef] [Green Version]
- Berridge, C.W.; Waterhouse, B.D. The locus coeruleus-noradrenergic system: Modulation of behavioral state and state-dependent cognitive processes. Brain Res. Rev. 2003, 42, 33–84. [Google Scholar] [CrossRef]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the Stress Response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Sara, S.J. The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci. 2009, 10, 211–223. [Google Scholar] [CrossRef]
- Pinos, H.; Collado, P.; Rodríguez-Zafra, M.; Rodríguez, C.; Segovia, S.; Guillamón, A. The development of sex differences in the locus coeruleus of the rat. Brain Res. Bull. 2001, 56, 73–78. [Google Scholar] [CrossRef]
- Pendergast, J.S.; Tuesta, L.M.; Bethea, J.R. Oestrogen receptor β contributes to the transient sex difference in tyrosine hydroxylase expression in the mouse locus coeruleus. J. Neuroendocrinol. 2008, 20, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.; Arai, O.; Ogata, R.; Omura, M.; Hori, T.; Aou, S. Exposure to bisphenol A during the fetal and suckling periods disrupts sexual differentiation of the locus coeruleus and of behavior in the rat. Neurosci. Lett. 2001, 304, 73–76. [Google Scholar] [CrossRef]
- Kubo, K.; Arai, O.; Omura, M.; Watanabe, R.; Ogata, R.; Aou, S. Low dose effects of bisphenol A on sexual differentiation of the brain and behavior in rats. Neurosci. Res. 2003, 45, 345–356. [Google Scholar] [CrossRef]
- Tando, S.; Itoh, K.; Yaoi, T.; Ogi, H.; Goto, S.; Mori, M.; Fushiki, S. Bisphenol A exposure disrupts the development of the locus coeruleus-noradrenergic system in mice. Neuropathology 2014, 34, 527–534. [Google Scholar] [CrossRef]
- Gioiosa, L.; Fissore, E.; Ghirardelli, G.; Parmigiani, S.; Palanza, P. Developmental exposure to low-dose estrogenic endocrine disruptors alters sex differences in exploration and emotional responses in mice. Horm. Behav. 2007, 52, 307–316. [Google Scholar] [CrossRef]
- Gioiosa, L.; Parmigiani, S.; vom Saal, F.S.; Palanza, P. The effects of bisphenol A on emotional behavior depend upon the timing of exposure, age and gender in mice. Horm. Behav. 2013, 63, 598–605. [Google Scholar] [CrossRef]
- Panzica, G.C.; Viglietti-Panzica, C.; Balthazart, J. The sexually dimorphic medial preoptic nucleus of quail: A key brain area mediating steroid action on male sexual behavior. Front. Neuroendocrinol. 1996, 17, 51–125. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, D.W.; Baum, M.J. Hormone-dependent medial preoptic/lumbar spinal cord/autonomic coordination supporting male sexual behaviors. Mol. Cell. Endocrinol. 2018, 467, 21–30. [Google Scholar] [CrossRef]
- Dobolyi, A.; Grattan, D.R.; Stolzenberg, D.S. Preoptic inputs and mechanisms that regulate maternal responsiveness. J. Neuroendocrinol. 2014, 26, 627–640. [Google Scholar] [CrossRef] [Green Version]
- Gorski, R.A.; Gordon, J.H.; Shryne, J.E.; Southam, A.M. Evidence for a morphological sex difference within the medial preoptic area of the rat brain. Brain Res. 1978, 148, 333–346. [Google Scholar] [CrossRef]
- Bleier, R.; Byne, W.; Siggelkow, I. Cytoarchitectonic sexual dimorphisms of the medial preoptic and anterior hypothalamic areas in guinea pig, rat, hamster, and mouse. J. Comp. Neurol. 1982, 212, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Tillet, Y.; Batailler, M.; Thibault, J. Neuronal projections to the medial preoptic area of the sheep, with special reference to monoaminergic afferents: Immunohistochemical and retrograde tract tracing studies. J. Comp. Neurol. 1993, 330, 195–220. [Google Scholar] [CrossRef]
- Simerly, R.B.; Zee, M.C.; Pendleton, J.W.; Lubahn, D.B.; Korach, K.S. Estrogen receptor-dependent sexual differentiation of dopaminergic neurons in the preoptic region of the mouse. Proc. Natl. Acad. Sci. USA 1997, 94, 14077–14082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbison, A.E. Noradrenergic regulation of cyclic GnRH secretion. Rev. Reprod. 1997, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vathy, I.; Etgen, A.M. Hormonal Activation of Female Sexual Behavior is Accompanied by Hypothalamic Norepinephrine Release. J. Neuroendocrinol. 1989, 1, 383–388. [Google Scholar] [CrossRef]
- Cornil, C.A.; Balthazart, J.; Motte, P.; Massotte, L.; Seutin, V. Dopamine activates noradrenergic receptors in the preoptic area. J. Neurosci. 2002, 22, 9320–9330. [Google Scholar] [CrossRef] [Green Version]
- Cornil, C.A.; Ball, G.F. Interplay among catecholamine systems: Dopamine binds to α 2-adrenergic receptors in birds and mammals. J. Comp. Neurol. 2008, 511, 610–627. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Paule, M.G.; Ferguson, S.A. Low oral doses of bisphenol A increase volume of the sexually dimorphic nucleus of the preoptic area in male, but not female, rats at postnatal day 21. Neurotoxicol. Teratol. 2012, 34, 331–337. [Google Scholar] [CrossRef]
- Rubin, B.S.; Lenkowski, J.R.; Schaeberle, C.M.; Vandenberg, L.N.; Ronsheim, P.M.; Soto, A.M. Evidence of altered brain sexual differentiation in mice exposed perinatally to low, environmentally relevant levels of bisphenol A. Endocrinology 2006, 147, 3681–3691. [Google Scholar] [CrossRef] [Green Version]
- Palanza, P.; Howdeshell, K.L.; Parmigiani, S.; vom Saal, F.S. Exposure to a low dose of bisphenol A during fetal life or in adulthood alters maternal behavior in mice. Environ. Health Perspect. 2002, 110, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.A.; Shimell, J.J.; Watson, N.V. Pre- and postnatal bisphenol A treatment results in persistent deficits in the sexual behavior of male rats, but not female rats, in adulthood. Horm. Behav. 2011, 59, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Farabollini, F.; Porrini, S.; Della Seta, D.; Bianchi, F.; Dessì-Fulgheri, F. Effects of perinatal exposure to bisphenol A on sociosexual behavior of female and male rats. Environ. Health Perspect. 2002, 110, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Laviola, G.; Gioiosa, L.; Adriani, W.; Palanza, P. D-Amphetamine-related reinforcing effects are reduced in mice exposed prenatally to estrogenic endocrine disruptors. Brain Res. Bull. 2005, 65, 235–240. [Google Scholar]
- Flügge, G. Alterations in the central nervous α2-adrenoceptor system under chronic psychosocial stress. Neuroscience 1996, 75, 187–196. [Google Scholar] [CrossRef]
- Starke, K. Presynaptic autoreceptors in the third decade: Focus on α2-adrenoceptors. J. Neurochem. 2001, 78, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Macmillan, L.B.; Fremeau, R.T.; Magnuson, M.A.; Lindner, J.; Limbird, L.E. Expression of α2-adrenergic receptor subtypes in the mouse brain: Evaluation of spatial and temporal information imparted by 3 kb of 5′ regulatory sequence for the α2A AR-receptor gene in transgenic animals. Neuroscience 1996, 74, 199–218. [Google Scholar] [CrossRef]
- Scheinin, M.; Lomasney, J.W.; Hayden-Hixson, D.M.; Schambra, U.B.; Caron, M.G.; Lefkowitz, R.J.; Fremeau, R.T. Distribution of α2-adrenergic receptor subtype gene expression in rat brain. Mol. Brain Res. 1994, 21, 133–149. [Google Scholar] [CrossRef]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Küppers, E.; Ivanova, T.; Karolczak, M.; Beyer, C. Estrogen: A multifunctional messenger to nigrostriatal dopaminergic neurons. J. Neurocytol. 2000, 29, 375–385. [Google Scholar] [CrossRef]
- Ivanova, T.; Beyer, C. Estrogen regulates tyrosine hydroxylase expression in the neonate mouse midbrain. J. Neurobiol. 2003, 54, 638–647. [Google Scholar] [CrossRef]
- Luque, J.M.; de Blas, M.R.; Segovia, S.; Guillamón, A. Sexual dimorphism of the dopamine-β-hydroxylase-immunoreactive neurons in the rat locus ceruleus. Dev. Brain Res. 1992, 67, 211–215. [Google Scholar] [CrossRef]
- Thanky, N.R.; Son, J.H.; Herbison, A.E. Sex differences in the regulation of tyrosine hydroxylase gene transcription by estrogen in the locus coeruleus of TH9-LacZ transgenic mice. Brain Res. Mol. Brain Res. 2002, 104, 220–226. [Google Scholar] [CrossRef]
- Johnson, A.E.; Nock, B.; McEwen, B.S.; Feder, H.H. A1- and A2-Noradrenergic Receptor Binding in Guinea Pig Brain: Sex Differences and Effects of Ovarian Steroids. Brain Res. 1988, 442, 205–213. [Google Scholar] [CrossRef]
- Karkanias, G.B.; Etgen, A.M. A thermodynamic analysis of estrogen regulation ofα2-adrenoceptor binding. Brain Res. 1995, 674, 26–32. [Google Scholar] [CrossRef]
- Díaz-Cabiale, Z.; Olausson, H.; Sohlström, A.; Agnati, L.F.; Narváez, J.A.; Uvnäs-Moberg, K.; Fuxe, K. Long-term modulation by postnatal oxytocin of the α 2-adrenoceptor agonist binding sites in central autonomic regions and the role of prenatal stress. J. Neuroendocrinol. 2004, 16, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Caldji, C.; Tannenbaum, B.; Sharma, S.; Francis, D.; Plotsky, P.M.; Meaney, M.J. Maternal care during infancy regulates the development of neural systems mediating the expression of fearfulness in the rat. Proc. Natl. Acad. Sci. USA 1998, 95, 5335–5340. [Google Scholar] [CrossRef] [Green Version]
- Ansonoff, M.A.; Etgen, A.M. Receptor phosphorylation mediates estradiol reduction of α 2-adrenoceptor coupling to G protein in the hypothalamus of female rats. Endocrine 2001, 14, 165–174. [Google Scholar] [CrossRef]
- O’Rourke, M.F.; Blaxall, H.S.; Iversen, L.J.; Bylund, D.B. Characterization of [3H]RX821002 binding to alpha-2 adrenergic receptor subtypes. J. Pharmacol. Exp. Ther. 1994, 268, 1362–1367. [Google Scholar]
- Karkanias, G.B.; Etgen, A.M. Estradiol attenuates α2-adrenoceptor-mediated inhibition of hypothalamic norepinephrine release. J. Neurosci. 1993, 13, 3448–3455. [Google Scholar] [CrossRef] [Green Version]
- Etgen, A.M.; Karkanias, G.B. Estrogen regulation of noradrenergic signaling in the hypothalamus. Psychoneuroendocrinology 1994, 19, 603–610. [Google Scholar] [CrossRef]
- Meston, C.M.; Moe, I.V.; Gorzalka, B.B. Effects of sympathetic inhibition on receptive, proceptive, and rejection behaviors in the female rat. Physiol. Behav. 1996, 59, 537–542. [Google Scholar] [CrossRef]
- Herbison, A.E.; Simonian, S.X.; Thanky, N.R.; Bicknell, R.J. Oestrogen modulation of noradrenaline neurotransmission. Novartis Found. Symp. 2000, 230, 74–93. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.M.; Vetrivelan, R.; Mallick, H.N. Noradrenergic afferents and receptors in the medial preoptic area: Neuroanatomical and neurochemical links between the regulation of sleep and body temperature. Neurochem. Int. 2007, 50, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Picot, M.; Naulé, L.; Marie-Luce, C.; Martini, M.; Raskin, K.; Grange-Messent, V.; Franceschini, I.; Keller, M.; Mhaouty-Kodja, S. Vulnerability of the neural circuitry underlying sexual behavior to chronic adult exposure to oral bisphenol A in male mice. Endocrinology 2014, 155, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, S.A.; Law, C.D. elber.; Kissling, G.E. Developmental treatment with ethinyl estradiol, but not bisphenol A, causes alterations in sexually dimorphic behaviors in male and female Sprague Dawley rats. Toxicol. Sci. 2014, 140, 374–392. [Google Scholar] [CrossRef] [Green Version]
- Naulé, L.; Picot, M.; Martini, M.; Parmentier, C.; Hardin-Pouzet, H.; Keller, M.; Franceschini, I.; Mhaouty-Kodja, S. Neuroendocrine and behavioral effects of maternal exposure to oral bisphenol A in female mice. J. Endocrinol. 2014, 220, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Bonthuis, P.J.; Cox, K.H.; Searcy, B.T.; Kumar, P.; Tobet, S.; Rissman, E.F. Of mice and rats: Key species variations in the sexual differentiation of brain and behavior. Front. Neuroendocrinol. 2010, 31, 341–358. [Google Scholar] [CrossRef] [Green Version]
- Beach, F.A. Sexual attractivity, proceptivity, and receptivity in female mammals. Horm. Behav. 1976, 7, 105–138. [Google Scholar] [CrossRef]
- McGill, T.E. Sexual Behavior in Three Inbred Strains of Mice. Behaviour 2008, 19, 341–350. [Google Scholar] [CrossRef]
- Flügge, G.; Ahrens, O.; Fuchs, E. Monoamine receptors in the prefrontal cortex of Tupaia belangeri during chronic psychosocial stress. Cell Tissue Res. 1997, 288, 1–10. [Google Scholar] [CrossRef]
- Palanza, P.; Nagel, S.C.; Parmigiani, S.; vom Saal, F.S. Perinatal exposure to endocrine disruptors: Sex, timing and behavioral endpoints. Curr. Opin. Behav. Sci. 2016, 7, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howdeshell, K.L.; Peterman, P.H.; Judy, B.M.; Taylor, J.A.; Orazio, C.E.; Ruhlen, R.L.; vom Saal, F.S.; Welshons, W.V. Bisphenol A is released from used polycarbonate animal cages into water at room temperature. Environ. Health Perspect. 2003, 111, 1180–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanza, P.; Gioiosa, L.; Parmigiani, S. Social stress in mice: Gender differences and effects of estrous cycle and social dominance. Physiol. Behav. 2001, 73, 411–420. [Google Scholar] [CrossRef]
- Dadomo, H.; Volpi, R.; Ferrari, M.; Vignali, A.; Bartolomucci, A.; Palanza, P.; Parmigiani, S. Sildenafil counteracts the inhibitory effect of social subordination on competitive aggression and sexual motivation in male mice. Behav. Brain Res. 2011, 216, 193–199. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The MouseBrain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Rich-Edwards, J.W.; Kaiser, U.B.; Chen, G.L.; Manson, J.A.E.; Goldstein, J.M. Sex and gender differences research design for basic, clinical, and population studies: Essentials for investigators. Endocr. Rev. 2018, 39, 424–439. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OIL | BPA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Females | Males | Females | Males | |||||||
| Outcome | M | SD | M | SD | d | M | SD | M | Sd | d |
| FOF | ||||||||||
| Exp | 143.10 | 105.78 | 100.05 | 74.48 | 0.46 [−0.27; 1.21] | 140.81 | 96.81 | 54.73 | 70.98 | 1.01 [0.22; 1.79] * |
| RA | 35.5 | 27.63 | 33.9 | 31.18 | 0.46 [−0.27; 1.21] | 27.92 | 31.03 | 19.93 | 17.14 | 0.43 [−0.31; 1.17] |
| Rear | 0.45 | 1.19 | 0.18 | 0.41 | 0.29 [−0.43; 1.02] | 0.12 | 0.48 | 1.35 | 1.96 | 0.85 [0.08; 1.61] |
| EPM | ||||||||||
| %CA | 56.37 | 12.52 | 71.96 | 13.03 | 1.21 [0.41; 2.02]* | 66.10 | 16.43 | 68.00 | 13.55 | 0.12 [−0.84; 0.60] |
| %CE | 31.45 | 11.23 | 18.84 | 7.14 | 1.33 [0.53; 2.14]* | 21.77 | 10.44 | 19.65 | 10.78 | 0.20 [−0.52; 0.92] |
| HD | 20.35 | 6.23 | 12.85 | 5.03 | 1.32 [0.52; 2.12]* | 14.42 | 4.92 | 13.78 | 5.95 | 0.11 [−0.60; 0.84] |
| RA | 21.87 | 6.66 | 20.42 | 4.53 | 0.25 [−0.55; 1.05] | 18.5 | 5.74 | 17.28 | 5.92 | 0.20 [−0.51; 0.93] |
| POA | ||||||||||
| Density | 177.81 | 15.92 | 161.97 | 13.25 | 1.08 [0.04; 2.11] | 182.40 | 19.42 | 174.87 | 17.02 | 0.40 [−0.53; 1.34] |
| Kd | 1.65 | 0.40 | 1.29 | 0.45 | 0.29 [−1.23; 0.65] | 1.70 | 0.46 | 1.59 | 0.37 | 0.26 [−0.73; 1.26] |
| LC | ||||||||||
| Density | 195.72 | 7.99 | 190.06 | 9.99 | 0.62 [−0.37; 1.62] | 195.00 | 14.61 | 192.88 | 9.43 | 0.17 [−0.81; 1.15] |
| Kd | 0.76 | 0.9 | 0.74 | 0.11 | 0.10 [−0.87; 1.08] | 0.92 | 0.15 | 0.88 | 0.15 | 0.23 [−0.47; 1.21] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponzi, D.; Gioiosa, L.; Parmigiani, S.; Palanza, P. Effects of Prenatal Exposure to a Low-Dose of Bisphenol A on Sex Differences in Emotional Behavior and Central Alpha2-Adrenergic Receptor Binding. Int. J. Mol. Sci. 2020, 21, 3269. https://doi.org/10.3390/ijms21093269
Ponzi D, Gioiosa L, Parmigiani S, Palanza P. Effects of Prenatal Exposure to a Low-Dose of Bisphenol A on Sex Differences in Emotional Behavior and Central Alpha2-Adrenergic Receptor Binding. International Journal of Molecular Sciences. 2020; 21(9):3269. https://doi.org/10.3390/ijms21093269
Chicago/Turabian StylePonzi, Davide, Laura Gioiosa, Stefano Parmigiani, and Paola Palanza. 2020. "Effects of Prenatal Exposure to a Low-Dose of Bisphenol A on Sex Differences in Emotional Behavior and Central Alpha2-Adrenergic Receptor Binding" International Journal of Molecular Sciences 21, no. 9: 3269. https://doi.org/10.3390/ijms21093269
APA StylePonzi, D., Gioiosa, L., Parmigiani, S., & Palanza, P. (2020). Effects of Prenatal Exposure to a Low-Dose of Bisphenol A on Sex Differences in Emotional Behavior and Central Alpha2-Adrenergic Receptor Binding. International Journal of Molecular Sciences, 21(9), 3269. https://doi.org/10.3390/ijms21093269