Resveratrol Induces Expression of Metabolic and Antioxidant Machinery and Protects Tilapia under Cold Stress


Abstract
:1. Introduction
2. Results
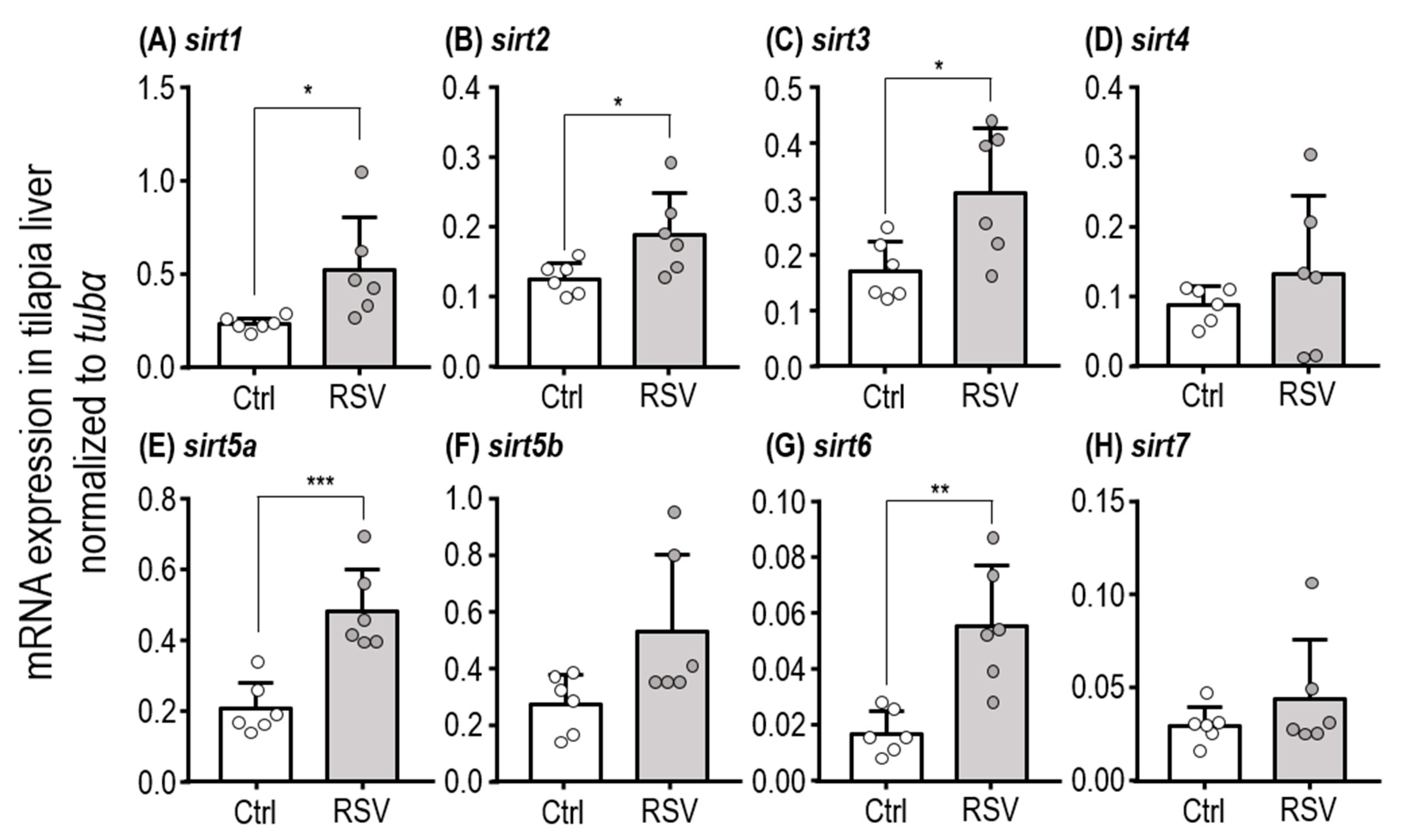
2.1. Subsection mRNA Expression of SIRT Homologues in the Liver
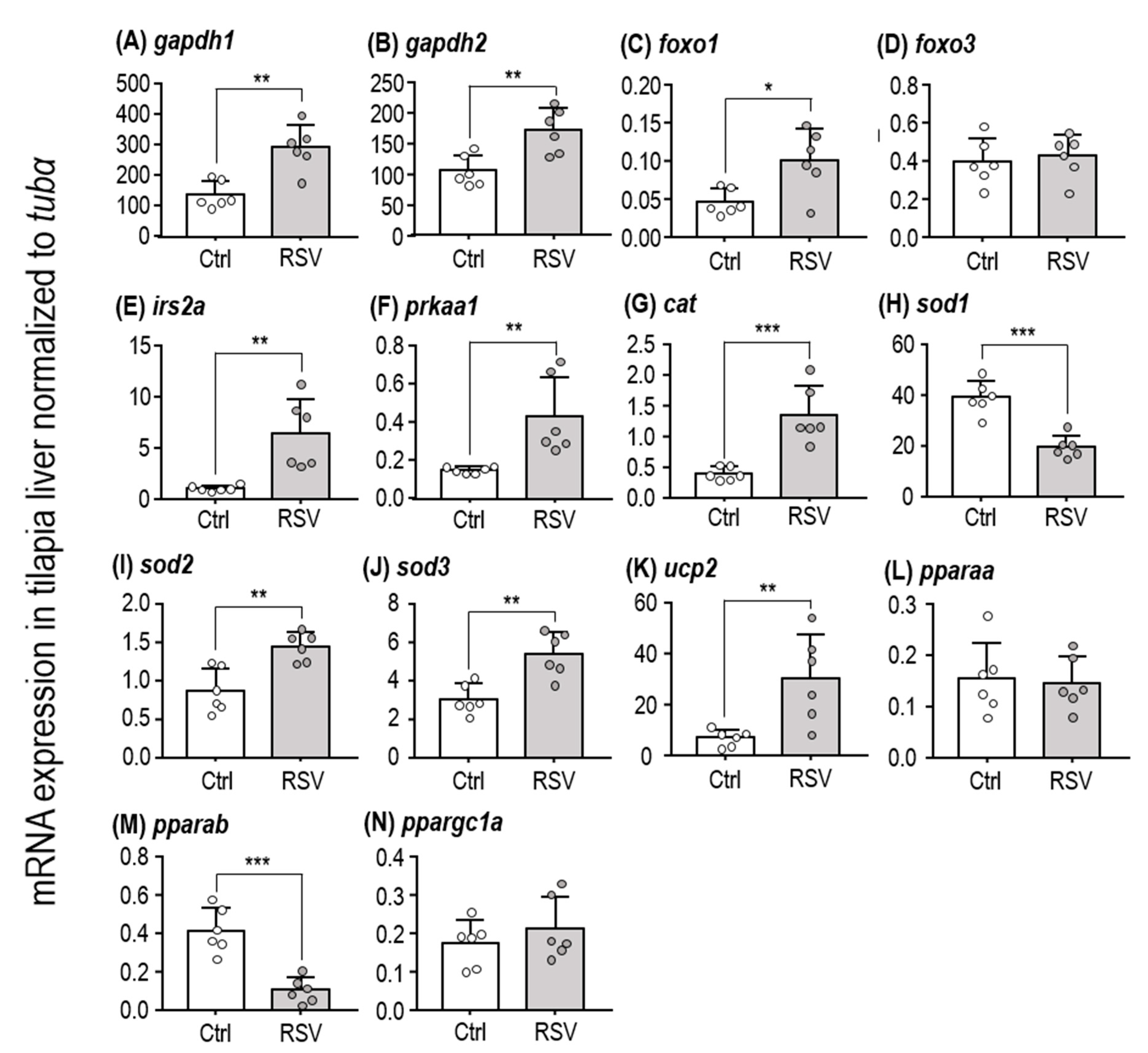
2.2. SIRT-Target Gene Expression After RSV Induction
2.3. Metabolic Response of Intact Tilapia to Cold Conditions
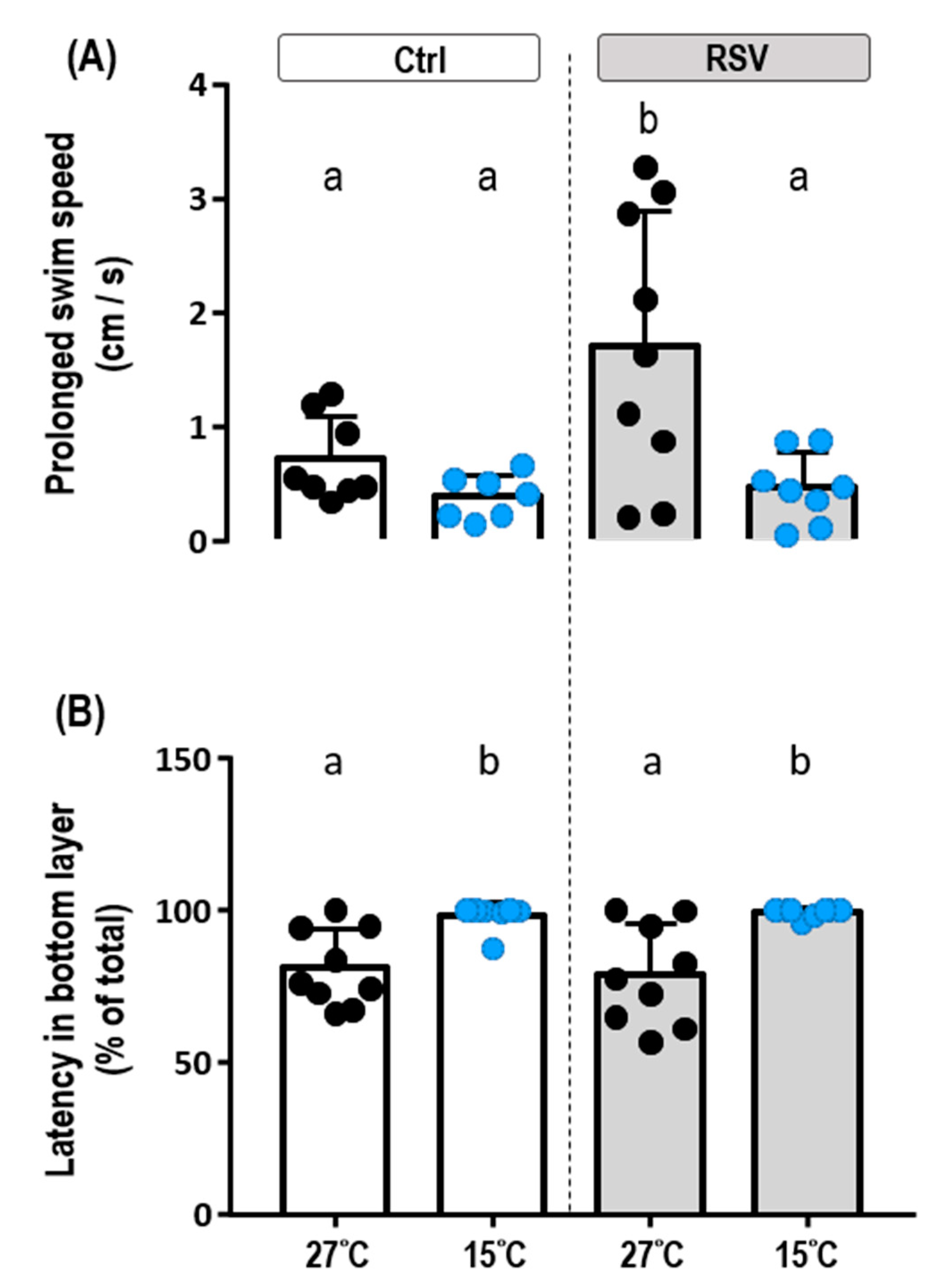
2.4. Locomotion under Cold Conditions
3. Discussion
4. Materials and Methods
4.1. Animal Breeding and Experimental Design
4.2. Purification of mRNA
4.3. Real-Time Quantitative PCR (qPCR)
4.4. Oxygen Consumption and Ammonium Excretion
4.5. Locomotion
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RSV | Resveratrol |
| ROS | Reactive oxygen species |
| SOD | superoxide dismutase |
| GPx | glutathione peroxidase |
| SIRTs | sirtuins |
| IRS | insulin receptor substrate |
| PPARs | peroxisome proliferator-activated receptors |
| FOXO | forkhead box protein |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| CAT | catalase |
| UCP | uncoupling protein |
References
- Friederich, M.; Olerud, J.; Fasching, A.; Liss, P.; Hansell, P.; Palm, F. Uncoupling protein-2 in diabetic kidneys: Increased protein expression correlates to increased non-transport related oxygen consumption. Adv. Exp. Med. Biol. 2008, 614, 37–43. [Google Scholar] [PubMed]
- Portner, H.O. Oxygen- and capacity-limitation of thermal tolerance: A matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 2010, 213, 881–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowyer, J.N.; Qin, J.G.; Stone, D.A.J. Protein, lipid and energy requirements of cultured marine fish in cold, temperate and warm water. Rev. Aquac. 2013, 5, 10–32. [Google Scholar] [CrossRef]
- Pörtner, H.-O. Physiological basis of temperature-dependent biogeography: Trade-offs in muscle design and performance in polar ectotherms. J. Exp. Biol. 2002, 205, 2217–2230. [Google Scholar]
- Sonna, L.; Fujita, J.; Gaffin, S.; Lilly, C. Molecular Biology of Thermoregulation: Invited Review: Effects of heat and cold stress on mammalian gene expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef] [Green Version]
- Ibarz, A.; Padrós, F.; Gallardo, M.Á.; Fernández-Borràs, J.; Blasco, J.; Tort, L. Low-temperature challenges to gilthead sea bream culture: Review of cold-induced alterations and ‘Winter Syndrome’. Rev. Fish Biol. Fish. 2010, 20, 539–556. [Google Scholar] [CrossRef]
- Donaldson, M.; Cooke, S.; Patterson, D.; Macdonald, J. Cold shock and fish. J. Fish Biol. 2008, 73, 1491–1530. [Google Scholar]
- Tseng, Y.C.; Chen, R.D.; Lucassen, M.; Schmidt, M.M.; Dringen, R.; Abele, D.; Hwang, P.P. Exploring uncoupling proteins and antioxidant mechanisms under acute cold exposure in brains of fish. PLoS ONE 2011, 6, e18180. [Google Scholar] [CrossRef] [Green Version]
- Costas, B.; Aragao, C.; Ruiz-Jarabo, I.; Vargas-Chacoff, L.; Arjona, F.J.; Mancera, J.M.; Dinis, M.T.; Conceicao, L.E. Different environmental temperatures affect amino acid metabolism in the eurytherm teleost Senegalese sole (Solea senegalensis Kaup, 1858) as indicated by changes in plasma metabolites. Amino Acids 2012, 43, 327–335. [Google Scholar] [CrossRef]
- Windisch, H.S.; Frickenhaus, S.; John, U.; Knust, R.; Portner, H.O.; Lucassen, M. Stress response or beneficial temperature acclimation: Transcriptomic signatures in Antarctic fish (Pachycara brachycephalum). Mol. Ecol. 2014, 23, 3469–3482. [Google Scholar] [CrossRef]
- Tseng, Y.C.; Hu, M.Y.; Stumpp, M.; Lin, L.Y.; Melzner, F.; Hwang, P.P. CO2-driven seawater acidification differentially affects development and molecular plasticity along life history of fish (Oryzias latipes). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 165, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.; Dunn, J. In Temperature acclimation and metabolism in ectotherms with particular reference to teleost fish. Symp. Soc. Exp. Biol. 1987, 41, 67–93. [Google Scholar] [PubMed]
- Abele, D.; Puntarulo, S. Formation of reactive species and induction of antioxidant defence systems in polar and temperate marine invertebrates and fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 138, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Malek, R.L.; Sajadi, H.; Abraham, J.; Grundy, M.A.; Gerhard, G.S. The effects of temperature reduction on gene expression and oxidative stress in skeletal muscle from adult zebrafish. Comp. Biochem. Phys. C Toxicol. Pharmacol. 2004, 138, 363–373. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol: Twenty Years of Growth, Development and Controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Kupis, W.; Palyga, J.; Tomal, E.; Niewiadomska, E. The role of sirtuins in cellular homeostasis. J. Physiol. Biochem. 2016, 72, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Oellerich, M.F.; Potente, M. FOXOs and sirtuins in vascular growth, maintenance, and aging. Circ. Res. 2012, 110, 1238–1251. [Google Scholar] [CrossRef] [Green Version]
- Meiliana, A.; Dewi, N.M.; Wijaya, A. Resveratrol: A sirtuin activator and the fountain of youth. Indones. Biomed. J. 2015, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Guarente, L. Sirtuins at a glance. J. Cell Sci. 2011, 124, 833–838. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Xiao, C.; Wang, R.H.; Lahusen, T.; Xu, X.; Vassilopoulos, A.; Vazquez-Ortiz, G.; Jeong, W.I.; Park, O.; Ki, S.H.; et al. Hepatic-specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis. Cell Metab. 2010, 12, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Nasrin, N.; Wu, X.; Fortier, E.; Feng, Y.; Bare, O.C.; Chen, S.; Ren, X.; Wu, Z.; Streeper, R.S.; Bordone, L. SIRT4 regulates fatty acid oxidation and mitochondrial gene expression in liver and muscle cells. J. Biol. Chem. 2010, 285, 31995–32002. [Google Scholar] [CrossRef] [Green Version]
- Silvestre, M.F.P.; Viollet, B.; Caton, P.W.; Leclerc, J.; Sakakibara, I.; Foretz, M.; Holness, M.C.; Sugden, M.C. The AMPK-SIRT signaling network regulates glucose tolerance under calorie restriction conditions. Life Sci. 2014, 100, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Zhao, Z.; Wu, W.; Song, C.; Meng, S.; Fan, L.; Bing, X.; Chen, J. Effects of dietary resveratrol supplementation on hepatic and serum pro-/anti-inflammatory activity in juvenile GIFT tilapia, Oreochromis niloticus. Dev. Comp. Immunol. 2017, 73, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.J.; Xu, C.; Liu, M.Y.; Wang, B.K.; Liu, W.B.; Chen, D.H.; Zhang, L.; Xu, C.Y.; Li, X.F. Resveratrol improves the energy sensing and glycolipid metabolism of blunt snout bream Megalobrama amblycephala fed high-carbohydrate diets by activating the AMPK-SIRT1-PGC-1alpha network. Front. Physiol. 2018, 9, 1258. [Google Scholar] [CrossRef] [PubMed]
- Pardal, D.; Caro, M.; Tueros, I.; Barranco, A.; Navarro, V. Resveratrol and piceid metabolites and their fat-reduction effects in zebrafish larvae. Zebrafish 2014, 11, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.J.; Sun, C.Y.; Wen, C.C.; Chen, Y.H. Nephroprotective role of resveratrol and ursolic acid in aristolochic acid intoxicated zebrafish. Toxins 2015, 7, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Qi, H.; Ma, L.; Liu, Z.; Fu, H.; Zhu, W.; Song, T.; Yang, B.; Li, G. Resveratrol attenuates oxidative stress and extends life span in the annual fish Nothobranchius guentheri. Rejuvenation Res. 2015, 18, 225–233. [Google Scholar] [CrossRef]
- Pereira, T.C.; Rico, E.P.; Rosemberg, D.B.; Schirmer, H.; Dias, R.D.; Souto, A.A.; Bonan, C.D.; Bogo, M.R. Zebrafish as a model organism to evaluate drugs potentially able to modulate sirtuin expression. Zebrafish 2011, 8, 9–16. [Google Scholar] [CrossRef]
- Hatting, M.; Tavares, C.D.J.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. N. Y. Acad. Sci. 2018, 1411, 21–35. [Google Scholar] [CrossRef]
- Oh, K.J.; Han, H.S.; Kim, M.J.; Koo, S.H. CREB and FoxO1: Two transcription factors for the regulation of hepatic gluconeogenesis. BMB Rep. 2013, 46, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Cunha, I.; Galante-Oliveira, S.; Rocha, E.; Planas, M.; Urbatzka, R.; Castro, L.F. Dynamics of PPARs, fatty acid metabolism genes and lipid classes in eggs and early larvae of a teleost. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 164, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yu, S.; Ying, J.; Shi, T.; Wang, P. Resveratrol prevents ROS-induced apoptosis in high glucose-treated retinal capillary endothelial cells via the activation of AMPK/Sirt1/PGC-1alpha pathway. Oxid. Med. Cell. Longev. 2017, 2017, 7584691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Tang, X.; Chen, H.-Z. Sirtuins and insulin resistance. Front. Endocrinol. 2018, 9, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordone, L.; Motta, M.C.; Picard, F.; Robinson, A.; Jhala, U.S.; Apfeld, J.; McDonagh, T.; Lemieux, M.; McBurney, M.; Szilvasi, A.; et al. Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic beta cells. PLoS Biol. 2006, 4, e31. [Google Scholar] [CrossRef] [Green Version]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.Y.; Lim, J.H.; Youn, H.H.; Hong, Y.A.; Yang, K.S.; Park, H.S.; Chung, S.; Ko, S.H.; Shin, S.J.; Choi, B.S.; et al. Resveratrol prevents renal lipotoxicity and inhibits mesangial cell glucotoxicity in a manner dependent on the AMPK-SIRT1-PGC1alpha axis in db/db mice. Diabetologia 2013, 56, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, M.-L.; Zhou, Y.; Yi, L.; Gao, Y.-X.; Ran, L.; Chen, S.-H.; Zhang, T.; Zhou, X.; Zou, D.; et al. Resveratrol improves hepatic steatosis by inducing autophagy through the cAMP signaling pathway. Mol. Nutr. Food Res. 2015, 59, 1443–1457. [Google Scholar] [CrossRef]
- Dasgupta, B.; Milbrandt, J. Resveratrol stimulates AMP kinase activity in neurons. Proc. Natl. Acad. Sci. USA 2007, 104, 7217–7222. [Google Scholar] [CrossRef] [Green Version]
- Bhullar, K.S.; Hubbard, B.P. Lifespan and healthspan extension by resveratrol. Biochim. Biophys. Acta 2015, 1852, 1209–1218. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D.; Rowe, M.; Butler, M.W.; McGraw, K.J. From molecules to living systems: Historical and contemporary issues in oxidative stress and antioxidant ecology. Funct. Ecol. 2010, 24, 950–959. [Google Scholar] [CrossRef]
- Lingappan, K. NF-kappaB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Blanquart, C.; Barbier, O.; Fruchart, J.C.; Staels, B.; Glineur, C. Peroxisome proliferator-activated receptors: Regulation of transcriptional activities and roles in inflammation. J. Steroid Biochem. Mol. Biol. 2003, 85, 267–273. [Google Scholar] [CrossRef]
- Randall, D.; Brauner, C. Effects of environmental factors on exercise in fish. J. Exp. Biol. 1991, 160, 113–126. [Google Scholar]
- Evans, D.H.; Claiborne, J.B. The Physiology of Fishes; Taylor & Francis: London, UK, 2005. [Google Scholar]
- Penghan, L.Y.; Cao, Z.D.; Fu, S.J. Effect of temperature and dissolved oxygen on swimming performance in crucian carp. Aquat. Biol. 2014, 21, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.; Wright, P. Nitrogen Excretion, 1st ed.; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Dolomatov, S.I.; Shekk, P.V.; Zukow, W.; Kryukova, M.I. Features of nitrogen metabolism in fishes. Rev. Fish Biol. Fish. 2011, 21, 733–737. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.C.; Hwang, P.P. Some insights into energy metabolism for osmoregulation in fish. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 148, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Ndong, D.; Chen, Y.Y.; Lin, Y.H.; Vaseeharan, B.; Chen, J.C. The immune response of tilapia Oreochromis mossambicus and its susceptibility to Streptococcus iniae under stress in low and high temperatures. Fish Shellfish Immunol. 2007, 22, 686–694. [Google Scholar] [CrossRef]
- Russell, D.J.; Thuesen, P.A.; Thomson, F.E. A review of the biology, ecology, distribution and control of Mozambique tilapia, Oreochromis mossambicus (Peters 1852) (Pisces: Cichlidae) with particular emphasis on invasive Australian populations. Rev. Fish Biol. Fish. 2012, 22, 533–554. [Google Scholar] [CrossRef]
- Zheng, Y.; Wu, W.; Hu, G.; Zhao, Z.; Meng, S.; Fan, L.; Song, C.; Qiu, L.; Chen, J. Hepatic transcriptome analysis of juvenile GIFT tilapia (Oreochromis niloticus), fed diets supplemented with different concentrations of resveratrol. Ecotoxicol. Environ. Saf. 2018, 147, 447–454. [Google Scholar] [CrossRef]
- Hu, M.Y.; Guh, Y.J.; Stumpp, M.; Lee, J.R.; Chen, R.D.; Sung, P.H.; Chen, Y.C.; Hwang, P.P.; Tseng, Y.C. Branchial NH4+-dependent acid–base transport mechanisms and energy metabolism of squid (Sepioteuthis lessoniana) affected by seawater acidification. Front. Zool. 2014, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, C.Y.; Tseng, Y.C.; Chen, Y.J.; Yang, Y.; Hsu, Y. Personality and physiological traits predict contest interactions in Kryptolebias marmoratus. Behav. Process. 2020, 173, 104079. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | t | df | p-Value |
|---|---|---|---|
| sirt1 | 2.509 | 10 | 0.0309 * |
| sirt2 | 2.452 | 10 | 0.0342 * |
| sirt3 | 2.727 | 10 | 0.0213 * |
| sirt4 | 0.9317 | 10 | 0.3735 |
| sirt5a | 4.808 | 10 | 0.0007 *** |
| sirt5b | 2.196 | 10 | 0.0528 |
| sirt6 | 4.071 | 10 | 0.0022 ** |
| sirt7 | 1.052 | 10 | 0.3176 |
| gapdh1 | 4.416 | 10 | 0.0013 ** |
| gapdh2 | 3.71 | 10 | 0.004 ** |
| foxo1 | 3.041 | 10 | 0.0124 * |
| foxo3 | 0.448 | 10 | 0.6637 |
| irs2a | 3.843 | 10 | 0.0033 ** |
| prkaa1 | 3.345 | 10 | 0.0074 ** |
| cat | 4.934 | 10 | 0.0006 *** |
| sod1 | 6.1 | 10 | 0.0001 *** |
| sod2 | 4.099 | 10 | 0.0021 ** |
| sod3 | 4.195 | 10 | 0.0018 ** |
| ucp2 | 3.282 | 10 | 0.0083 ** |
| pparaa | 0.2313 | 10 | 0.8217 |
| pparab | 5.691 | 10 | 0.0002 *** |
| ppargc1a | 0.9309 | 10 | 0.3739 |
| df | MS | F | p | |
|---|---|---|---|---|
| Oxygen consumption rate | ||||
| Feeding | 1 | 816 | 7.811 | 0.0087 |
| Temperature | 1 | 35651 | 340.9 | <0.0001 |
| Feeding × Temperature | 1 | 3.317 | 0.0317 | 0.8598 |
| Error | 32 | 104.6 | ||
| Ammonium excretion rate | ||||
| Feeding | 1 | 0.2527 | 0.0831 | 0.7749 |
| Temperature | 1 | 11.83 | 3.89 | 0.0573 |
| Feeding × Temperature | 1 | 0.1439 | 0.0473 | 0.8291 |
| Error | 32 | 3.04 | ||
| O: N ratio | ||||
| Feeding | 1 | 99.18 | 0.5132 | 0.4789 |
| Temperature | 1 | 3664 | 18.96 | 0.0001 |
| Feeding × Temperature | 1 | 95.17 | 0.4925 | 0.4879 |
| Error | 32 | 193.2 | ||
| Prolong swim speed | ||||
| Feeding | 1 | 2.296 | 4.873 | 0.0356 |
| Temperature | 1 | 4.916 | 10.43 | 0.0032 |
| Feeding × Temperature | 1 | 1.678 | 3.561 | 0.0696 |
| Error | 28 | 0.4712 | ||
| Latency in bottom layer | ||||
| Feeding | 1 | 1.911 | 0.0151 | 0.9028 |
| Temperature | 1 | 2856 | 22.67 | <0.0001 |
| Feeding × Temperature | 1 | 21.97 | 0.1744 | 0.6793 |
| Error | 29 | 126 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.-C.; Wang, Y.-C.; Peng, H.-W.; Hseu, J.-R.; Wu, G.-C.; Chang, C.-F.; Tseng, Y.-C. Resveratrol Induces Expression of Metabolic and Antioxidant Machinery and Protects Tilapia under Cold Stress. Int. J. Mol. Sci. 2020, 21, 3338. https://doi.org/10.3390/ijms21093338
Wang M-C, Wang Y-C, Peng H-W, Hseu J-R, Wu G-C, Chang C-F, Tseng Y-C. Resveratrol Induces Expression of Metabolic and Antioxidant Machinery and Protects Tilapia under Cold Stress. International Journal of Molecular Sciences. 2020; 21(9):3338. https://doi.org/10.3390/ijms21093338
Chicago/Turabian StyleWang, Min-Chen, Yu-Chun Wang, Hui-Wen Peng, Jinn-Rong Hseu, Guan-Chung Wu, Ching-Fong Chang, and Yung-Che Tseng. 2020. "Resveratrol Induces Expression of Metabolic and Antioxidant Machinery and Protects Tilapia under Cold Stress" International Journal of Molecular Sciences 21, no. 9: 3338. https://doi.org/10.3390/ijms21093338
APA StyleWang, M. -C., Wang, Y. -C., Peng, H. -W., Hseu, J. -R., Wu, G. -C., Chang, C. -F., & Tseng, Y. -C. (2020). Resveratrol Induces Expression of Metabolic and Antioxidant Machinery and Protects Tilapia under Cold Stress. International Journal of Molecular Sciences, 21(9), 3338. https://doi.org/10.3390/ijms21093338