Caviar Extract and Its Constituent DHA Inhibits UVB-Irradiated Skin Aging by Inducing Adiponectin Production

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Caviar Extract on Cell Viability
2.2. Effects of Caviar Extract on Adipocyte Differentiation
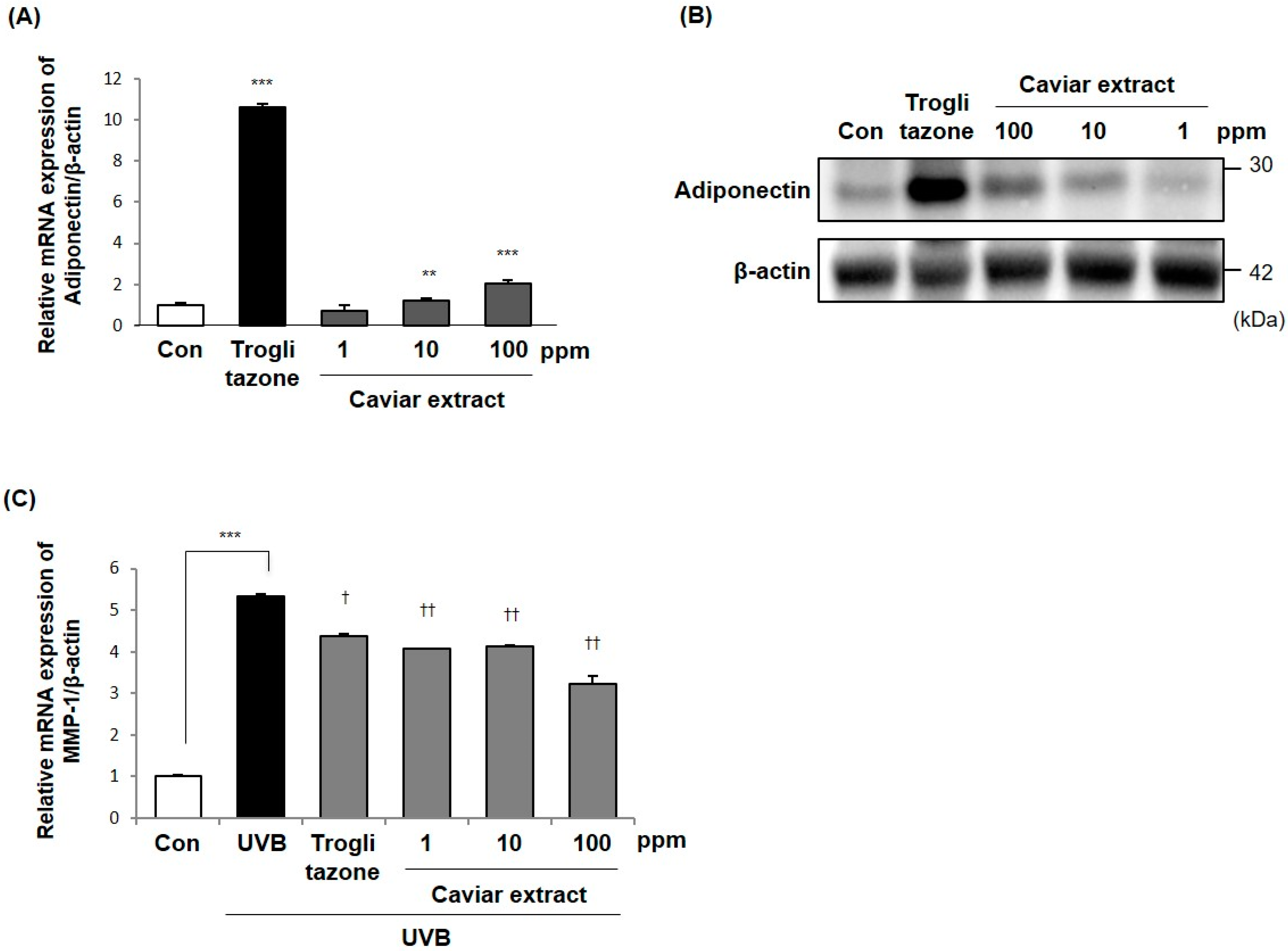
2.3. Effects of Caviar Extract on Adiponectin Production
2.4. Analysis of Lipid Composition of Caviar Extract
2.5. Effects of DHA on Adipocyte Differentiation
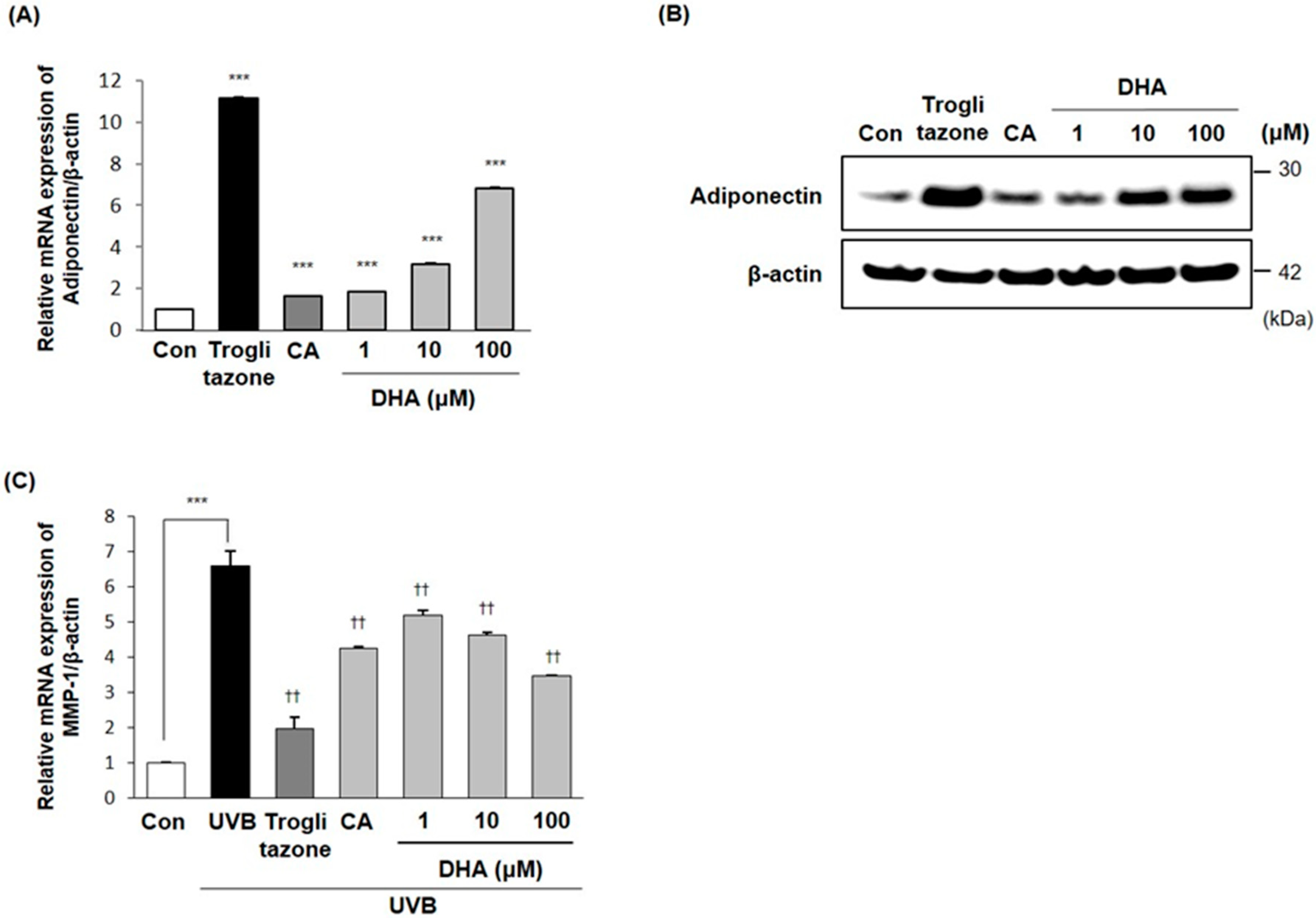
2.6. Effects of DHA on Adiponectin Production
2.7. Inhibitory Effects of DHA on MMP-1 Expression in Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Adipocyte Differentiation
4.2. Cell Viability Assay
4.3. Oil Red O Staining
4.4. RT-qPCR
4.5. Immunoblotting Analysis
4.6. Proximate Composition and Fatty Acid Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Antibiotic Antimycotic Solution |
| ATCC | American Type Culture Collection |
| C/EBPα | CCAAT-enhancer-binding protein alpha |
| DHA | Docosahexaenoic acid |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethylsulfoxide |
| EPA | Eicosapentaenoic acid |
| ECM | Extracellular matrix |
| FBS | Fetal bovine serum |
| HDFs | Human dermal fibroblasts |
| IBMX | 3-Isobutyl-1-methylxanthine |
| MMP-1 | Matrix metalloproteinase-1 |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NBS | Newborn bovine serum |
| OD | Optical density |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| PUFA | Polyunsaturated fatty acid |
| RIPA | Radioimmunoprecipitation assay |
| RT-qPCR | Quantitative real-time polymerase chain reaction |
| SREBP-1a | Sterol regulatory element binding protein-1a |
| UVB | Ultraviolet B |
References
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef]
- Boer, M.; Duchnik, E.; Maleszka, R.; Marchlewicz, M. Structural and biophysical characteristics of human skin in maintaining proper epidermal barrier function. Postepy Dermatol. Alergol. 2016, 33, 1–5. [Google Scholar] [CrossRef]
- Lim, J.Y.; Kim, O.K.; Lee, J.; Lee, M.J.; Kang, N.; Hwang, J.K. Protective effect of the standardized green tea seed extract on UVB-induced skin photoaging in hairless mice. Nutr. Res. Pract. 2014, 8, 398–403. [Google Scholar] [CrossRef] [Green Version]
- Park, P.J.; Cho, E.G. Kojyl Cinnamate Ester Derivatives Increase Adiponectin Expression and Stimulate Adiponectin-Induced Hair Growth Factors in Human Dermal Papilla Cells. Int. J. Mol. Sci. 2019, 20, 15. [Google Scholar] [CrossRef] [Green Version]
- Ezure, T.; Amano, S. Influence of subcutaneous adipose tissue mass on dermal elasticity and sagging severity in lower cheek. Skin Res. Technol. 2010, 16, 332–338. [Google Scholar] [CrossRef]
- Kim, W.S.; Park, B.S.; Sung, J.H. Protective role of adipose-derived stem cells and their soluble factors in photoaging. Arch. Dermatol. Res. 2009, 301, 329–336. [Google Scholar] [CrossRef]
- Wojciechowicz, K.; Markiewicz, E.; Jahoda, C.A. C/EBPalpha identifies differentiating preadipocytes around hair follicles in foetal and neonatal rat and mouse skin. Exp. Dermatol. 2008, 17, 675–680. [Google Scholar] [CrossRef]
- Rivera-Gonzalez, G.; Shook, B.; Horsley, V. Adipocytes in Skin Health and Disease. Cold Spring Harb. Perspect. Med. 2014, 4, a015271. [Google Scholar] [CrossRef]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef] [Green Version]
- Weiss, R.; Taksali, S.E.; Dufour, S.; Yeckel, C.W.; Papademetris, X.; Cline, G.; Tamborlane, W.V.; Dziura, J.; Shulman, G.I.; Caprio, S. The “obese insulin-sensitive” adolescent: Importance of adiponectin and lipid partitioning. J. Clin. Endocrinol. Metab. 2005, 90, 3731–3737. [Google Scholar] [CrossRef] [Green Version]
- Salathia, N.S.; Shi, J.; Zhang, J.; Glynne, R.J. An in vivo screen of secreted proteins identifies adiponectin as a regulator of murine cutaneous wound healing. J. Investig. Dermatol. 2013, 133, 812–821. [Google Scholar] [CrossRef] [Green Version]
- Shibata, S.; Tada, Y.; Asano, Y.; Hau, C.S.; Kato, T.; Saeki, H.; Yamauchi, T.; Kubota, N.; Kadowaki, T.; Sato, S. Adiponectin regulates cutaneous wound healing by promoting keratinocyte proliferation and migration via the ERK signaling pathway. J. Immunol. 2012, 189, 3231–3341. [Google Scholar] [CrossRef] [Green Version]
- Wolk, K.; Sabat, R. Adipokines in psoriasis: An important link between skin inflammation and metabolic alterations. Rev. Endocr. Metab. Disord. 2016, 17, 305–317. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, Y.K.; Kim, M.K.; Kim, S.; Kim, J.Y.; Lee, D.H.; Chung, J.H. UV-induced inhibition of adipokine production in subcutaneous fat aggravates dermal matrix degradation in human skin. Sci. Rep. 2016, 6, 25616. [Google Scholar] [CrossRef] [Green Version]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, A.; Sriwiriyanont, P.; Fujimura, T.; Ohuchi, A.; Kitahara, T.; Takema, Y.; Kitzmiller, W.J.; Visscher, M.O.; Tsuboi, R.; Boissy, R.E. Mechanistic effects of long-term ultraviolet B irradiation induce epidermal and dermal changes in human skin xenografts. Am. J. Pathol. 2009, 174, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.C.; Kiezel-Tsugunova, M.; Brownbridge, L.C.; Harwood, J.L.; Nicolaou, A. Lipid functions in skin: Differential effects of n-3 polyunsaturated fatty acids on cutaneous ceramides, in a human skin organ culture model. Biochim. Biophys. Acta Biomembr. 2017, 21, 1679–1689. [Google Scholar] [CrossRef]
- McDaniel, J.C.; Massey, K.; Nicolaou, A. Fish oil supplementation alters levels of lipid mediators of inflammation in microenvironment of acute human wounds. Wound Repair Regen. 2011, 19, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Yasutomo, K.; Watanabe, T. Treatment with DHA/EPA ameliorates atopic dermatitis-like skin disease by blocking LTB4 production. J. Med. Investig. 2016, 63, 187–191. [Google Scholar] [CrossRef]
- Liu, P.; Li, G.; Wu, J.; Zhou, X.; Wang, L.; Han, W.; Lv, Y.; Sun, C. Vaspin promotes 3T3-L1 preadipocyte differentiation. Exp. Biol. Med. (Maywood) 2015, 240, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Hadrich, F.; Sayadi, S. Apigetrin inhibits adipogenesis in 3T3-L1 cells by downregulating PPARγ and CEBP-α. Lipids Health Dis. 2018, 17, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Li, C.; Lv, Y.; Zhang, Y.; Amakye, W.K.; Mao, L. DHA increases adiponectin expression more than effectively than EPA at relative low concentrations by regulating PPARγ and its phosphorylation at Ser273 in 3T3-L1 adipocytes. Nutr. Metab. (London) 2017, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali, G.; Desouza, C.V.; Clevenger, M.E.; Ramalingam, R.; Sarawathi, V. Differential effects of eicosapentaenoic acid and docosahexaenoic acid in promoting the differentiation of 3T3-L1 preadipocytes. Prostaglandins Leukot Essent Fatty Acids 2014, 90, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Abbas, E.M.; Hrachya, G.H. Fatty acid composition of caviar and liver from cultured great sturgeon (Huso huso). Int. Food Res. J. 2015, 22, 1083–1086. [Google Scholar]
- Caprino, F.; Moretti, V.M.; Bellagmba, F.; Turchini, G.M.; Busetto, M.L.; Giani, I.; Paleari, M.A.; Pazzaglia, M. Fatty acid composition and volatile compounds of caviar from farmed white sturgeon (Acipenser transmontanus). Anal. Chim. Acta 2008, 617, 139–147. [Google Scholar] [CrossRef]
- Czesny, S.; Dbrowski, K.; Christensen, J.E.; Van Eenennaam, J.; Doroshov, S. Discrimination of wild and domestic origin of sturgeon ova on lipids and fatty acid analysis. Aquaculture 2000, 189, 145–153. [Google Scholar] [CrossRef]
- Mol, S.; Turan, S. Comparison of proximate, fatty acid and amino acid compositions of various types of fish roes. Int. J. Food Prop. 2008, 11, 669–677. [Google Scholar] [CrossRef]
- Shin, J.H.; Oliveira, A.C.; Rasco, R.A. Quality attributes and microbial storage stability of caviar from cultivated white sturgeon (Acipenser transmontanus). J. Food Sci. 2010, 75, 43–48. [Google Scholar] [CrossRef]
- Wirth, M.; Kirschbaum, F.; Gessner, J.; Willot, P.; Patriche, N.; Billard, R. Fatty acid composition in sturgeon caviar from different species: Comparing wild and farmed origins. Int. Rev. Hydrobiol. 2002, 87, 629–636. [Google Scholar] [CrossRef]
- Nordoy, A.; Marchioli, R.; Arnesen, H.; Videbaek, J. n-3 polyunsaturated fatty acids and cardiovascular diseases. Lipids 2001, 36, 127–129. [Google Scholar] [CrossRef]
- Talukdar, S.; Olefsky, J.M.; Osborn, O. Targeting GPR120 and other fatty acid-sensing GPCRs ameliorates insulin resistance and inflammatory diseases. Trends Pharmacol. Sci. 2011, 32, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiecolt-Glaser, J.K.; Belury, M.A.; Andridge, R.; Malarkey, W.B.; Hwang, B.S.; Gkaser, R. Omega-3 supplementation lowers inflammation in healthy middle-aged and older adults: A randomized controlled trial. Brain Behav. Immun. 2012, 26, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marotta, F.; Polimeni, A.; Solimene, U.; Lorenzetti, A.; Minelli, E.; Jain, S.; Rastmanesh, R.; Sedriep, S.; Soresi, Y. Beneficial modulation from a high-purity caviar-derived homogenate on chronological skin aging. Rejuvenation Res. 2012, 15, 174–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, R.T.; Tishinsky, J.M.; Yuan, Z.; Robinson, L.E. Docosahexanoic acid incrases cellular adiponectin mRNA and secreted adiponectin protein, as well as PPARγ mRNA, in 3T3-L1 adipocytes. Appl. Physiol. Nutr. Metab. 2010, 35, 783–789. [Google Scholar] [CrossRef]
- Itoh, M.; Suganami, T.; Satoh, N.; Tanimoto-Koyama, K.; Yuan, Z.; Tanaka, M.; Kawano, H.; Yano, H.; Aoe, S.; Takeya, M.; et al. Increased adiponectin secretion by highly purified eicosapentaenoic acid in rodent models of obesity and human obese subjects. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1918–1925. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Liu, F. Transcriptional and post-translational regulation of adiponectin. Biochem. J. 2009, 425, 41–52. [Google Scholar] [CrossRef]
- Seo, J.B.; Moon, H.M.; Noh, M.J.; Jeong, H.W.; Yoo, E.J.; Kim, W.S.; Park, J.; Youn, B.S.; Kim, J.W.; Park, S.D.; et al. Adipocyte determination- and differentiation-dependent factor 1/sterol regulatory element binding protein 1c regulates mouse adiponectin expression. J. Biol. Chem. 2004, 279, 22108–22117. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Lipid Numbers | Mean |
|---|---|---|
| Myristic acid | C14:0 | 0.06 g/100 g |
| Behenic Acid | C22:0 | 0.00 g/100 g |
| γ-Linolenic acid | C18:3, ω-6 | 0.09 g/100 g |
| Linolenic acid | C18:2, ω-6 | 0.66 g/100 g |
| Stearic Acid | C18.0 | 0.23 g/100 g |
| Arachidonic Acid | C20:4, ω-6 | 0.24 g/100 g |
| Arachidic Acid | C20.0 | 0.01 g/100 g |
| α-Linolenic acid | C18:3, ω-3 | 0.09 g/100 g |
| Palmitoleic acid | C16:1, ω-7 | 0.34 g/100 g |
| Heptadecanoic acid | C17:0 | 0.02 g/100 g |
| Elaidic acid | C18:1n9t | 0.04 g/100 g |
| Linolelaidic acid | C18:2 | 0.08 g/100 g |
| Cis-11-eicosenoic acid | C20:1 | 0.07 g/100 g |
| Eicosadienoic acid | C20:2, ω-6 | 0.04 g/100 g |
| Cis-8 11 14-eicosatrienoic acid | C20:3n6 | 0.05 g/100 g |
| Cis-11, 14, 17-eicosatrienoic acid | C20:3n3 | 0.01 g/100 g |
| Eicosapentaenoic acid | C20:5, ω-3 | 0.24 g/100 g |
| Docosahexaenoic acid | C22:6, ω-3 | 1.16 g/100 g |
| Cis-15-tetracosenoic acid | C24:1, ω-9 | 0.02 g/100 g |
| Oleic acid | C18:1, ω-9 | 2.72 g/100 g |
| Palmitic acid | C16:0 | 1.54 g/100 g |
| Pentadecanoic acid | C15:0 | 0.01 g/100 g |
| Erucic acid | C22:1, ω-9 | 0.01 g/100 g |
| Protein | 23.98% | |
| Lipid | 10.45% | |
| Moisture | 55.96% | |
| Ash | 5.12% | |
| Carbohydrate | 0.09% | |
| Dietary fiber | 4.40% |
| Item | Condition |
|---|---|
| Column | SPTM-2560 (100 m × 0.25 mm × 0.20 µm) |
| Detector | FID |
| Injection Temp. | 225 °C |
| Injection Volume | 1.0 µL |
| Detector Temp. | 285 °C |
| Oven Temp. | 100 °C (4 min), 208 °C (3 °C/min, 5 min), 244 °C (2 °C/min, 15 min) |
| Carrier Gas | He |
| Column Flow | 0.75 mL/min |
| Split Ratio | 200:1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-E.; Nho, Y.-H.; Yun, S.K.; Park, S.-M.; Kang, S.; Yeo, H. Caviar Extract and Its Constituent DHA Inhibits UVB-Irradiated Skin Aging by Inducing Adiponectin Production. Int. J. Mol. Sci. 2020, 21, 3383. https://doi.org/10.3390/ijms21093383
Lee K-E, Nho Y-H, Yun SK, Park S-M, Kang S, Yeo H. Caviar Extract and Its Constituent DHA Inhibits UVB-Irradiated Skin Aging by Inducing Adiponectin Production. International Journal of Molecular Sciences. 2020; 21(9):3383. https://doi.org/10.3390/ijms21093383
Chicago/Turabian StyleLee, Kyung-Eun, Youn-Hwa Nho, Seok Kyun Yun, Sung-Min Park, Seunghyun Kang, and Hyeonju Yeo. 2020. "Caviar Extract and Its Constituent DHA Inhibits UVB-Irradiated Skin Aging by Inducing Adiponectin Production" International Journal of Molecular Sciences 21, no. 9: 3383. https://doi.org/10.3390/ijms21093383
APA StyleLee, K.-E., Nho, Y.-H., Yun, S. K., Park, S.-M., Kang, S., & Yeo, H. (2020). Caviar Extract and Its Constituent DHA Inhibits UVB-Irradiated Skin Aging by Inducing Adiponectin Production. International Journal of Molecular Sciences, 21(9), 3383. https://doi.org/10.3390/ijms21093383