Molecular Aspects and Prognostic Significance of Microcalcifications in Human Pathology: A Narrative Review

 ,
,  and
and 
Abstract
:1. Introduction
Molecular Mechanisms Involved in the Formation of Microcalcifications
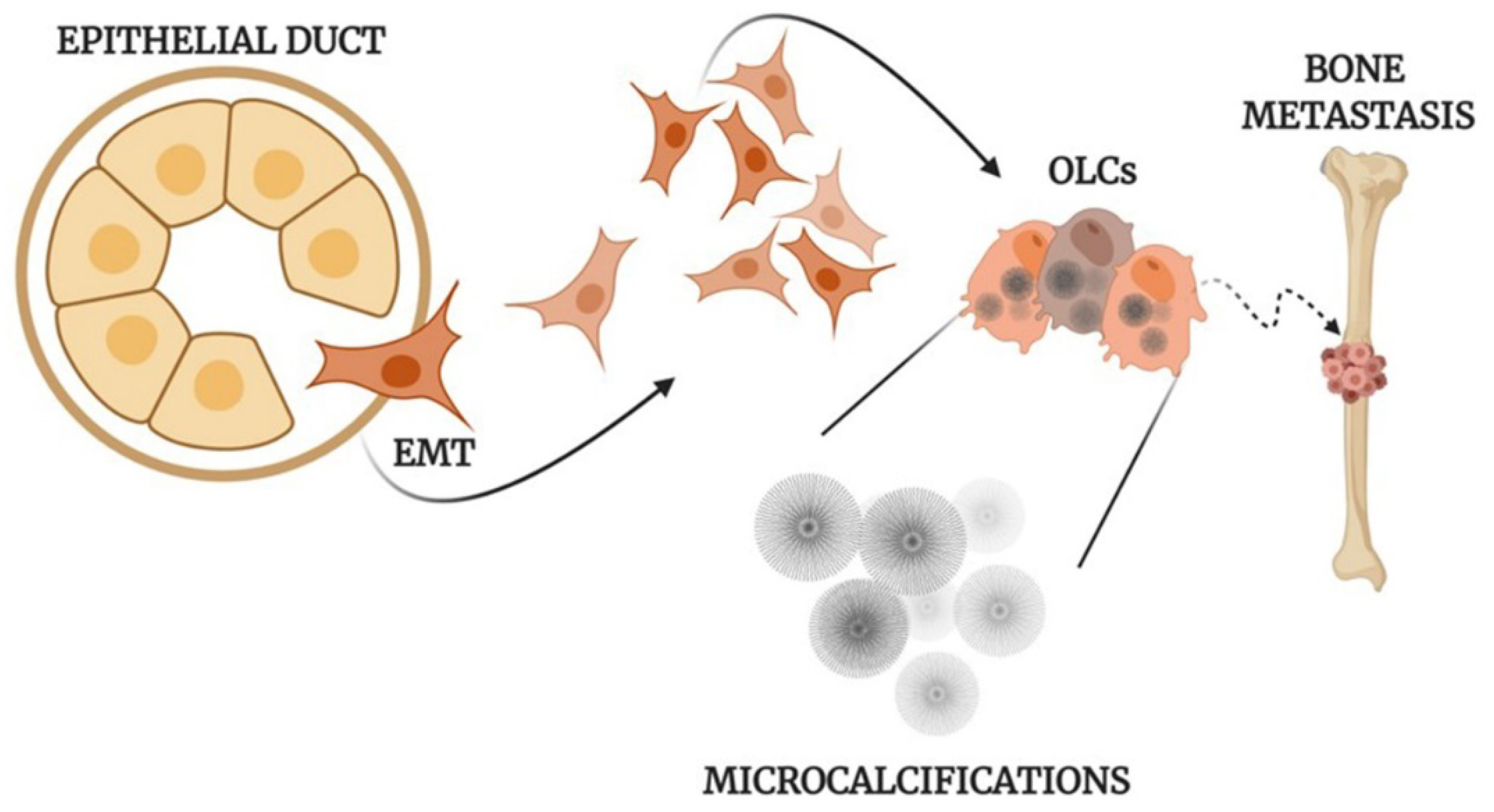
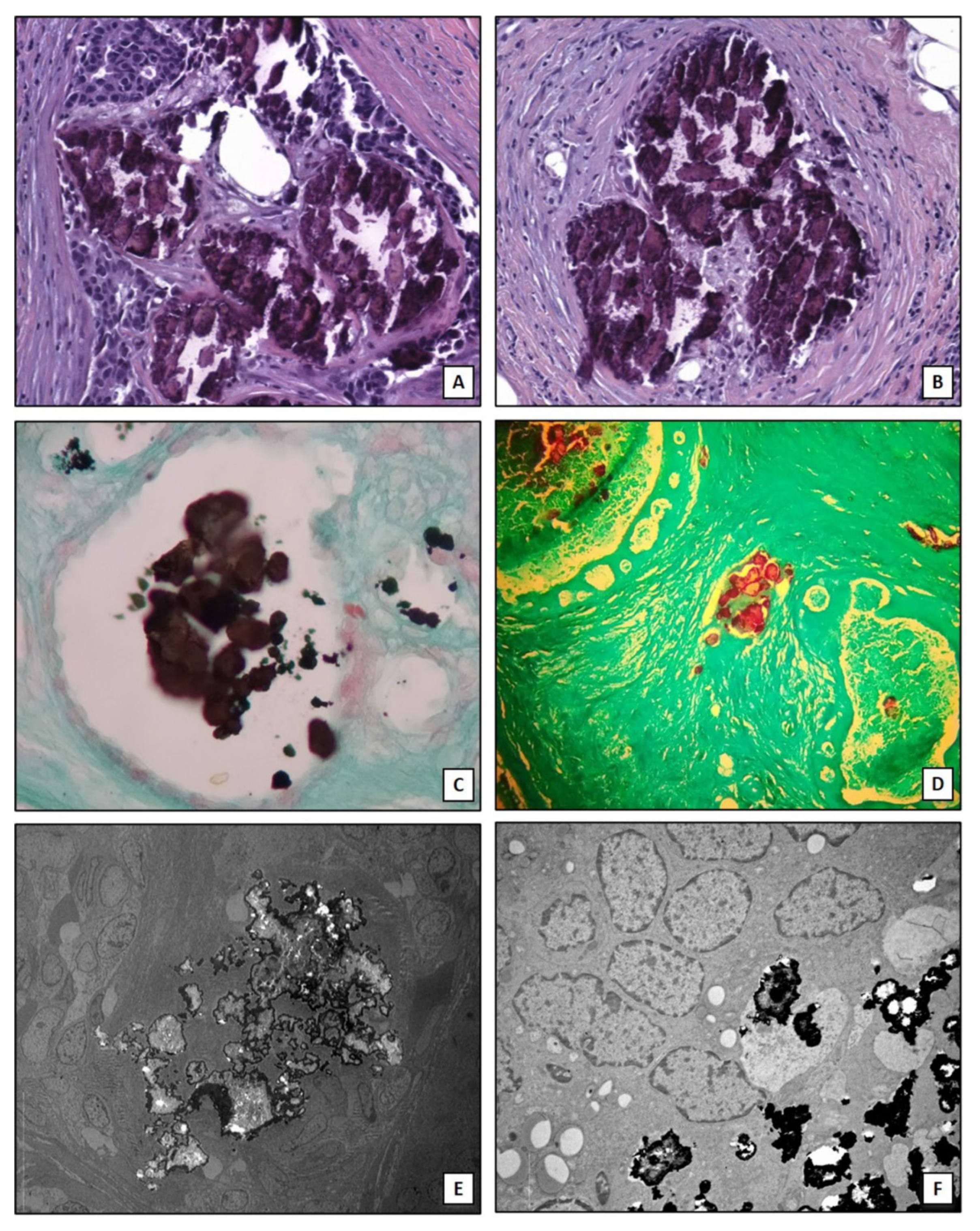
2. Microcalcifications in Breast Cancer
3. Microcalcifications in Thyroid Lesions
4. Microcalcifications in Lung Pathogenesis
5. Microcalcifications in Ovary Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| BMPs | Bone morphogenetic proteins |
| BOLCs | Breast Osteoblast-Like Cells |
| CO | Calcium oxalate |
| CT | Computed tomography |
| EDX | Energy dispersive X-ray analysis |
| EMT | Epithelial-to-mesenchymal transition |
| ER | Estrogen receptor |
| HOXA9 | Homeobox A9 |
| HA | Hydroxyapatite |
| HER2 | Human epidermal growth factor receptor-2 |
| LCIS | Lobular carcinoma in situ |
| Mg-HAp | Magnesium-substituted hydroxyapatite |
| micro-FTIR | Micro-Fourier transform infrared |
| OPN | Osteopontin |
| PTC | Papillary thyroid carcinoma |
| PTX3 | Pentraxin-related protein 3 |
| POLCs | Prostate osteoblast-like cells |
| PBs | Psammoma bodies |
| RANKL | Receptor activator of nuclear factor kappa-B ligand |
| RUNX2 | Runt-related transcription factor 2 |
| SEM | Scanning electron microscope |
| TRPM7 | Transient receptor potential cation channel subfamily M |
| US | Ultrasonography |
| VSMCs | Vascular smooth muscle cells |
| VOLCs | Vascular osteoblast-like cells |
References
- Baldwin, P. Breast calcification imaging. Radiol. Technol. 2013, 84, 383M–404M, quiz 405M–408M. [Google Scholar]
- Horvat, J.V.; Keating, D.M.; Rodrigues-Duarte, H.; Morris, E.A.; Mango, V.L. Calcifications at Digital Breast Tomosynthesis: Imaging Features and Biopsy Techniques. RadioGraphics 2019, 39, 307–318. [Google Scholar] [CrossRef]
- Li, X.; Gao, H.; Chen, Z.; Zhang, L.; Zhu, X.; Wang, S.; Peng, W. Diagnosis of breast cancer based on microcalcifications using grating-based phase contrast CT. Eur. Radiol. 2018, 28, 3742–3750. [Google Scholar] [CrossRef] [PubMed]
- Spak, D.; Plaxco, J.; Santiago, L.; Dryden, M.; Dogan, B. BI-RADS ® fifth edition: A summary of changes. Diagn. Interv. Imaging 2017, 98, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Malguria, N.; Zimmerman, S.; Fishman, E.K. Coronary Artery Calcium Scoring. J. Comput. Assist. Tomogr. 2018, 42, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xu, W.; Zeng, H.; He, Z.; Lu, X.; Zuo, D.; Qin, G.; Chen, W. OXPHOS-dependent metabolic reprogramming prompts metastatic potential of breast cancer cells under osteogenic differentiation. Br. J. Cancer 2020, 123, 1644–1655. [Google Scholar] [CrossRef]
- Sanchez-Duffhues, G.; De Vinuesa, A.G.; Dijke, P.T. Endothelial-to-mesenchymal transition in cardiovascular diseases: Developmental signaling pathways gone awry. Dev. Dyn. 2018, 247, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Sharma, T.; Radosevich, J.A.; Pachori, G.; Mandal, C.C. A Molecular View of Pathological Microcalcification in Breast Cancer. J. Mammary Gland. Biol. Neoplasia 2016, 21, 25–40. [Google Scholar] [CrossRef]
- Xie, F.; Li, F.; Li, R.; Liu, Z.; Shi, J.; Zhang, C.; Dong, N. Inhibition of PP2A enhances the osteogenic differentiation of human aortic valvular interstitial cells via ERK and p38 MAPK pathways. Life Sci. 2020, 257, 118086. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, X.; Wu, H. Arterial Stiffness: A Focus on Vascular Calcification and Its Link to Bone Mineralization. Arter. Thromb. Vasc. Biol. 2020, 40, 1078–1093. [Google Scholar] [CrossRef]
- Cho, K.I.; Sakuma, I.; Sohn, I.S.; Jo, S.H.; Koh, K.K. Inflammatory and metabolic mechanisms underlying the calcific aortic valve disease. Atherosclerosis 2018, 277, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.F.; Morgan, M.P. Microcalcifications in breast cancer: Lessons from physiological mineralization. Bone 2013, 53, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Weinbaum, S. Microcalcifications, Their Genesis, Growth, and Biomechanical Stability in Fibrous Cap Rupture. Adv. Exp. Med. Biol. 2018, 1097, 129–155. [Google Scholar] [CrossRef] [PubMed]
- Bischetti, S.; Scimeca, M.; Bonanno, E.; Federici, M.; Anemona, L.; Menghini, R.; Casella, S.; Cardellini, M.; Ippoliti, A.; Mauriello, A. Carotid plaque instability is not related to quantity but to elemental composition of calcification. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, I.; Perri, E. Ultrastructural, Elemental and Mineralogical Analysis of Vascular Calcification in Atherosclerosis. Microsc. Microanal. 2017, 23, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, R.; Scimeca, M.; Toschi, N.; Pistolese, C.A.; Giannini, E.; Antonacci, C.; Ciuffa, S.; Tancredi, V.; Tarantino, U.; Albonici, L.; et al. Radiological, Histological and Chemical Analysis of Breast Microcalcifications: Diagnostic Value and Biological Significance. J. Mammary Gland. Biol. Neoplasia 2018, 23, 89–99. [Google Scholar] [CrossRef]
- Chang, H.H.; Cheng, C.L.; Huang, P.J.; Lin, S.Y. Application of scanning electron microscopy and X-ray microanalysis: FE-SEM, ESEM-EDS, and EDS mapping for studying the characteristics of topographical microstructure and elemental mapping of human cardiac calcified deposition. Anal. Bioanal Chem. 2014, 406, 359–366. [Google Scholar] [CrossRef]
- Marra, S.P.; Daghlian, C.P.; Fillinger, M.F.; Kennedy, F.E. Elemental composition, morphology and mechanical properties of calcified deposits obtained from abdominal aortic aneurysms. Acta Biomater. 2006, 2, 515–520. [Google Scholar] [CrossRef]
- Scimeca, M.; Giannini, E.; Antonacci, C.; Pistolese, C.A.; Spagnoli, L.G.; Bonanno, E. Microcalcifications in breast cancer: An active phenomenon mediated by epithelial cells with mesenchymal characteristics. BMC Cancer 2014, 14, 286. [Google Scholar] [CrossRef] [Green Version]
- O’Grady, S.; Morgan, M.P. Deposition of calcium in an in vitro model of human breast tumour calcification reveals functional role for ALP activity, altered expression of osteogenic genes and dysregulation of the TRPM7 ion channel. Sci. Rep. 2019, 9, 542. [Google Scholar] [CrossRef]
- Morgan, M.P.; Cooke, M.M.; McCarthy, G.M. Microcalcifications associated with breast cancer: An epiphenomenon or biologically significant feature of selected tumors? J. Mammary Gland Biol. Neoplasia 2005, 10, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.F.; Hernandez-Santana, A.; Ramdass, S.; McMahon, G.; Harmey, J.H.; Morgan, M.P. Microcalcifications in breast cancer: Novel insights into the molecular mechanism and functional consequence of mammary mineralisation. Br. J. Cancer 2012, 106, 525–537. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, S.; Morgan, M.P. Microcalcifications in breast cancer: From pathophysiology to diagnosis and prognosis. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, R.; Scimeca, M.; Urbano, N.; Bonanno, E.; Schillaci, O. Breast microcalcifications: Biological and diagnostic perspectives. Future Oncol. 2018, 14, 3097–3099. [Google Scholar] [CrossRef] [PubMed]
- Scimeca, M.; Bonfiglio, R.; Montanaro, M.; Bonanno, E. Osteoblast-like cells in human cancers: New cell type and reliable markers for bone metastasis. Future Oncol. 2018, 14, 9–11. [Google Scholar] [CrossRef]
- Scimeca, M.; Urbano, N.; Bonfiglio, R.; Schillaci, O.; Bonanno, E. Breast osteoblast-like cells: A new biomarker for the management of breast cancer. Br. J. Cancer 2018, 119, 1129–1132. [Google Scholar] [CrossRef] [Green Version]
- Scimeca, M.; Bonfiglio, R.; Menichini, E.; Albonici, L.; Urbano, N.; De Caro, M.T.; Mauriello, A.; Schillaci, O.; Gambacurta, A.; Bonanno, E. Microcalcifications Drive Breast Cancer Occurrence and Development by Macrophage-Mediated Epithelial to Mesenchymal Transition. Int. J. Mol. Sci. 2019, 20, 5633. [Google Scholar] [CrossRef] [Green Version]
- Scimeca, M.; Antonacci, C.; Toschi, N.; Giannini, E.; Bonfiglio, R.; Buonomo, C.O.; Pistolese, C.A.; Tarantino, U.; Bonanno, E. Breast Osteoblast-like Cells: A Reliable Early Marker for Bone Metastases from Breast Cancer. Clin. Breast Cancer 2018, 18, e659–e669. [Google Scholar] [CrossRef]
- Scimeca, M.; Bonfiglio, R.; Varone, F.; Ciuffa, S.; Mauriello, A.; Bonanno, E. Calcifications in prostate cancer: An active phenomenon mediated by epithelial cells with osteoblast-phenotype. Microsc. Res. Tech. 2018, 81, 745–748. [Google Scholar] [CrossRef]
- Scimeca, M.; Urbano, N.; Bonfiglio, R.; Mapelli, S.N.; Catapano, C.V.; Carbone, G.M.; Ciuffa, S.; Tavolozza, M.; Schillaci, O.; Mauriello, A.; et al. Prostate Osteoblast-Like Cells: A Reliable Prognostic Marker of Bone Metastasis in Prostate Cancer Patients. Contrast Media Mol. Imaging 2018, 2018, 9840962, Erratum in: Contrast Media Mol. Imaging 2019, 2019, 7843735. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.L.; Liu, J.; Del Castillo, M.; Shah, T. Role of Coronary Calcium Score to Identify Candidates for ASCVD Prevention. Curr. Atheroscler. Rep. 2019, 21, 53. [Google Scholar] [CrossRef] [PubMed]
- Durham, A.L.; Speer, M.Y.; Scatena, M.; Giachelli, C.M.; Shanahan, C.M. Role of smooth muscle cells in vascular calcification: Implications in atherosclerosis and arterial stiffness. Cardiovasc. Res. 2018, 114, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Barrett, H.E.; Van der Heiden, K.; Farrell, E.; Gijsen, F.J.H.; Akyildiz, A.C. Calcifications in atherosclerotic plaques and impact on plaque biomechanics. J. Biomech. 2019, 87, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.L.; Weinbaum, S.; Aikawa, E.; Hutcheson, J.D. Zooming in on the genesis of atherosclerotic plaque microcalcifications. J. Physiol. 2016, 594, 2915–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinnouchi, H.; Sato, Y.; Sakamoto, A.; Cornelissen, A.; Mori, M.; Kawakami, R.; Gadhoke, N.V.; Kolodgie, F.D.; Virmani, R.; Finn, A.V. Calcium deposition within coronary atherosclerotic lesion: Implications for plaque stability. Atherosclerosis 2020, 306, 85–95. [Google Scholar] [CrossRef]
- Shanahan, C.M. Inflammation ushers in calcification: A cycle of damage and protection? Circulation 2007, 116, 2782–2785. [Google Scholar] [CrossRef]
- Lomashvili, K.A.; Cobbs, S.; Hennigar, R.A.; Hardcastle, K.I.; O’Neill, W.C. Phosphate-induced vascular calcification: Role of pyrophosphate and osteopontin. J. Am. Soc. Nephrol. 2004, 15, 1392–1401. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, F.; Sakakura, K.; Yahagi, K.; Joner, M.; Virmani, R. Has our understanding of calcification in human coronary atherosclerosis progressed? Arter. Thromb. Vasc. Biol. 2014, 34, 724–736. [Google Scholar] [CrossRef] [Green Version]
- Pugliese, G.; Iacobini, C.; Blasetti Fantauzzi, C.; Menini, S. The dark and bright side of atherosclerotic calcification. Atherosclerosis 2015, 238, 220–230. [Google Scholar] [CrossRef]
- New, S.E.; Aikawa, E. Role of extracellular vesicles in de novo mineralization: An additional novel mechanism of cardiovascular calcification. Arter. Thromb. Vasc Biol. 2013, 33, 1753–1758. [Google Scholar] [CrossRef] [Green Version]
- Anderson, H.C. Mechanisms of pathologic calcification. Rheum. Dis. Clin. North Am. 1988, 14, 303–319. [Google Scholar] [PubMed]
- Bobryshev, Y.V.; Killingsworth, M.C.; Lord, R.S.; Grabs, A.J. Matrix vesicles in the fibrous cap of atherosclerotic plaque: Possible contribution to plaque rupture. J. Cell. Mol. Med. 2008, 12, 2073–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- New, S.E.; Goettsch, C.; Aikawa, M.; Marchini, J.F.; Shibasaki, M.; Yabusaki, K.; Libby, P.; Shanahan, C.M.; Croce, K.; Aikawa, E. Macrophage-derived matrix vesicles: An alternative novel mechanism for microcalcification in atherosclerotic plaques. Circ. Res. 2013, 113, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, A.N.; Davies, J.D.; Reynolds, J.L.; McNair, R.; Jones, G.T.; Sidibe, A.; Schurgers, L.J.; Skepper, J.N.; Proudfoot, D.; Mayr, M.; et al. Calcium regulates key components of vascular smooth muscle cell-derived matrix vesicles to enhance mineralization. Circ. Res. 2011, 109, e1–e12. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, A.; McGregor, D.H.; Anderson, H.C. Matrix vesicles in atherosclerotic calcification. Proc. Soc. Exp. Biol. Med. 1983, 172, 173–177. [Google Scholar] [CrossRef]
- Nakahara, T.; Dweck, M.R.; Narula, N.; Pisapia, D.; Narula, J.; Strauss, H.W. Coronary Artery Calcification: From Mechanism to Molecular Imaging. JACC Cardiovasc. Imaging 2017, 10, 582–593. [Google Scholar] [CrossRef]
- Anderson, H.C. Molecular biology of matrix vesicles. Clin. Orthop. Relat. Res. 1995, 314, 266–280. [Google Scholar] [CrossRef]
- Chen, N.X.; O’Neill, K.D.; Chen, X.; Moe, S.M. Annexin-mediated matrix vesicle calcification in vascular smooth muscle cells. J. Bone Miner. Res. 2008, 23, 1798–1805. [Google Scholar] [CrossRef] [Green Version]
- Warner, G.P.; Hubbard, H.L.; Lloyd, G.C.; Wuthier, R.E. 32Pi- and 45Ca-metabolism by matrix vesicle-enriched microsomes prepared from chicken epiphyseal cartilage by isosmotic Percoll density-gradient fractionation. Calcif. Tissue Int. 1983, 35, 327–338. [Google Scholar] [CrossRef]
- Nancollas, G.H.; LoRe, M.; Perez, L.; Richardson, C.; Zawacki, S.J. Mineral phases of calcium phosphate. Anat. Rec. 1989, 224, 234–241. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, C.; Wu, Y.; Zhu, Y.; Ren, Y.; Tong, Z. Microcalcification and BMP-2 in breast cancer: Correlation with clinicopathological features and outcomes. Oncotargets Ther. 2019, 12, 2023–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balleyguier, C.; Ayadi, S.; Van Nguyen, K.; Vanel, D.; Dromain, C.; Sigal, R. BIRADS classification in mammography. Eur. J. Radiol. 2007, 61, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Haka, A.S.; Shafer-Peltier, K.E.; Fitzmaurice, M.; Crowe, J.; Dasari, R.R.; Feld, M.S. Identifying microcalcifications in benign and malignant breast lesions by probing differences in their chemical composition using Raman spectroscopy. Cancer Res. 2002, 62, 5375–5380. [Google Scholar] [PubMed]
- Büsing, C.M.; Keppler, U.; Menges, V. Differences in microcalcification in breast tumors. Virchows Arch. A 1981, 393, 307–313. [Google Scholar] [CrossRef]
- Scimeca, M.; Trivigno, D.; Bonfiglio, R.; Ciuffa, S.; Urbano, N.; Schillaci, O.; Bonanno, E. Breast cancer metastasis to bone: From epithelial to mesenchymal transition to breast osteoblast-like cells. Semin. Cancer Biol. 2020, S1044-579X(20)30005-5. [Google Scholar] [CrossRef]
- Crescitelli, M.C.; Rauschemberger, M.B.; Cepeda, S.; Sandoval, M.; Massheimer, V.L. Role of estrone on the regulation of osteoblastogenesis. Mol. Cell. Endocrinol. 2019, 498, 110582. [Google Scholar] [CrossRef]
- Scimeca, M.; Giocondo, R.; Montanaro, M.; Granaglia, A.; Bonfiglio, R.; Tancredi, V.; Mauriello, A.; Urbano, N.; Schillaci, O.; Bonanno, E. BMP-2 Variants in Breast Epithelial to Mesenchymal Transition and Microcalcifications Origin. Cells 2020, 9, 1381. [Google Scholar] [CrossRef]
- Ali, M.A.; Czene, K.; Hall, P.; Humphreys, K. Association of Microcalcification Clusters with Short-term Invasive Breast Cancer Risk and Breast Cancer Risk Factors. Sci. Rep. 2019, 9, 14604. [Google Scholar] [CrossRef]
- Gao, B.; Zhang, H.; Zhang, S.D.; Cheng, X.Y.; Zheng, S.M.; Sun, Y.H.; Zhang, D.W.; Jiang, Y.; Tian, J.W. Mammographic and clinicopathological features of triple-negative breast cancer. Br. J. Radiol. 2014, 87, 20130496. [Google Scholar] [CrossRef] [Green Version]
- Fanizzi, A.; Basile, T.M.; Losurdo, L.; Bellotti, R.; Bottigli, U.; Campobasso, F.; Didonna, V.; Fausto, A.; Massafra, R.; Tagliafico, A.; et al. Ensemble Discrete Wavelet Transform and Gray-Level Co-Occurrence Matrix for Microcalcification Cluster Classification in Digital Mammography. Appl. Sci. 2019, 9, 5388. [Google Scholar] [CrossRef]
- Russ, G.; Bonnema, S.J.; Erdogan, M.F.; Durante, C.; Ngu, R.; Leenhardt, L. European Thyroid Association Guidelines for Ultrasound Malignancy Risk Stratification of Thyroid Nodules in Adults: The EU-TIRADS. Eur. Thyroid. J. 2017, 6, 225–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wienke, J.R.; Chong, W.K.; Fielding, J.R.; Zou, K.H.; Mittelstaedt, C.A. Sonographic features of benign thyroid nodules: Interobserver reliability and overlap with malignancy. J. Ultrasound Med. 2003, 22, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hambleton, C.; Kandil, E. Appropriate and accurate diagnosis of thyroid nodules: A review of thyroid fine-needle aspiration. Int. J. Clin. Exp. Med. 2013, 6, 413–422. [Google Scholar] [PubMed]
- Shin, J.H.; Baek, J.H.; Chung, J.; Ha, E.J.; Kim, J.H.; Lee, Y.H.; Lim, H.K.; Moon, W.J.; Na, D.G.; Park, J.S.; et al. Korean Society of Thyroid Radiology (KSThR) and Korean Society of Radiology. Ultrasonography Diagnosis and Imaging-Based Management of Thyroid Nodules: Revised Korean Society of Thyroid Radiology Consensus Statement and Recommendations. Korean J. Radiol. 2016, 17, 370–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Q.; Zhang, Q.; Chen, S.; Niu, C. Petal-Like Calcifications in Thyroid Nodules on Ultrasonography: A Rare Morphologic Characteristic of Calcification Associated with Aggressive Biological Behavior. Front. Endocrinol. (Lausanne) 2020, 11, 271. [Google Scholar] [CrossRef]
- Na, D.G.; Kim, D.S.; Kim, S.J.; Ryoo, J.W.; Jung, S.L. Thyroid nodules with isolated macrocalcification: Malignancy risk and diagnostic efficacy of fine-needle aspiration and core needle biopsy. Ultrasonography 2016, 35, 212–219. [Google Scholar] [CrossRef]
- Grani, G.; Sponziello, M.; Pecce, V.; Ramundo, V.; Durante, C. Contemporary Thyroid Nodule Evaluation and Management. J. Clin. Endocrinol. Metab. 2020, 105, 2869–2883. [Google Scholar] [CrossRef]
- Woon, T.K.; Sanamandra, S.K.; Salkade, P.R.; Venkatanarasimha, N.; Ganguly, R.; Ho, C.L.; Bakar, R.A.; Chua, J.M.; Tan, B.S.; Damodharan, K. Subcentimetre thyroid nodules: Sonographic features associated with malignancy. Ultrasound 2020, 28, 155–163. [Google Scholar] [CrossRef]
- Guerlain, J.; Perie, S.; Lefevre, M.; Perez, J.; Vandermeersch, S.; Jouanneau, C.; Huguet, L.; Frochot, V.; Letavernier, E.; Weil, R.; et al. Localization and characterization of thyroid microcalcifications: A histopathological study. PLoS ONE 2019, 14, e0224138. [Google Scholar] [CrossRef] [Green Version]
- Das, D.K. Psammoma body: A product of dystrophic calcification or of a biologically active process that aims at limiting the growth and spread of tumor? Diagn. Cytopathol. 2009, 37, 534–541. [Google Scholar] [CrossRef]
- De Santis, S.; Sotgiu, G.; Crescenzi, A.; Taffon, C.; Felici, A.C.; Orsini, M. On the chemical composition of psammoma bodies microcalcifications in thyroid cancer tissues. J. Pharm. Biomed. Anal. 2020, 190, 113534. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Ohta, K.; Kobayashi, T. Expression and function of Cbfa-1/Runx2 in thyroid papillary carcinoma cells. J. Clin. Endocrinol. Metab. 2008, 93, 2409–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Fan, T.C.; Yu, J.C.; Liao, G.S.; Lin, Y.C.; Shih, A.C.; Li, W.H.; Yu, A.L. The prognostic significance of RUNX2 and miR-10a/10b and their inter-relationship in breast cancer. J. Transl. Med. 2014, 12, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonare, L.D.; Frigo, A.; Francia, G.; Davì, M.V.; Donatelli, L.; Stranieri, C.; Brazzarola, P.; Zatelli, M.C.; Menestrina, F.; Valenti, M.T. Runx2 mRNA expression in the tissue, serum, and circulating non-hematopoietic cells of patients with thyroid cancer. J. Clin. Endocrinol. Metab. 2012, 97, E1249–E1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Kim, H.K.; Lee, J.; Soh, E.Y.; Kim, J.H.; Song, I.; Chung, Y.S.; Choi, Y.J. Transcription Factor HOXA9 is Linked to the Calcification and Invasion of Papillary Thyroid Carcinoma. Sci. Rep. 2019, 9, 6773. [Google Scholar] [CrossRef] [Green Version]
- Grier, D.G.; Thompson, A.; Kwasniewska, A.; McGonigle, G.J.; Halliday, H.L.; Lappin, T.R. The pathophysiology of HOX genes and their role in cancer. J. Pathol. 2005, 205, 154–171. [Google Scholar] [CrossRef]
- Brassard, M.; Borget, I.; Edet-Sanson, A.; Giraudet, A.L.; Mundler, O.; Toubeau, M.; Bonichon, F.; Borson-Chazot, F.; Leenhardt, L.; Schvartz, C.; et al. THYRDIAG Working Group. Long-term follow-up of patients with papillary and follicular thyroid cancer: A prospective study on 715 patients. J. Clin. Endocrinol. Metab. 2011, 96, 1352–1359. [Google Scholar] [CrossRef]
- Park, K.W.; Shin, J.H.; Han, B.K.; Ko, E.Y.; Chung, J.H. Inoperable symptomatic recurrent thyroid cancers: Preliminary result of radiofrequency ablation. Ann. Surg. Oncol. 2011, 18, 2564–2568. [Google Scholar] [CrossRef]
- Wang, L.; Ge, M.; Xu, D.; Chen, L.; Qian, C.; Shi, K.; Liu, J.; Chen, Y. Ultrasonography-guided percutaneous radiofrequency ablation for cervical lymph node metastasis from thyroid carcinoma. J. Cancer Res. Ther. 2014, 10 (Suppl. C144–C149). [Google Scholar] [CrossRef]
- Chan, E.D.; Morales, D.V.; Welsh, C.H.; McDermott, M.T.; Schwarz, M.I. Calcium deposition with or without bone formation in the lung. Am. J. Respir. Crit. Care Med. 2002, 165, 1654–1669. [Google Scholar] [CrossRef] [Green Version]
- Bendayan, D.; Barziv, Y.; Kramer, M.R. Pulmonary calcifications: A review. Respir. Med. 2000, 94, 190–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.G.; MacMahon, H.; Vyborny, C.J.; Pollak, E.R. Dystrophic calcification in carcinoma of the lung: Demonstration by CT. AJR Am. J. Roentgenol. 1987, 148, 29–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkill, G.J.; Allen, S.D.; A’hern, R.P.; Gore, M.E.; King, D.M. Significance of tumour calcification in ovarian carcinoma. Br. J. Radiol. 2009, 82, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Inomata, M.; Hayashi, R.; Kambara, K.; Okazawa, S.; Imanishi, S.; Ichikawa, T.; Suzuki, K.; Yamada, T.; Miwa, T.; Kashii, T.; et al. Miliary brain metastasis presenting with calcification in a patient with lung cancer: A case report. J. Med. Case Rep. 2012, 6, 279. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, Y.; Homma, T.; Kohga, H.; Uki, J.; Shisa, H. A lung cancer case with numerous calcified metastatic nodules of the brain. Neuroradiology 1988, 30, 265–268. [Google Scholar] [CrossRef]
- Wen, J.; Miao, Y.; Wang, S.; Tong, R.; Zhao, Z.; Wu, J. Calcification: A Disregarded or Ignored Issue in the Gynecologic Tumor Microenvironments. Int. J. Gynecol. Cancer 2018, 28, 486–492. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef]
- Outwater, E.K.; Siegelman, E.S.; Hunt, J.L. Ovarian teratomas: Tumor types and imaging characteristics. Radiographics 2001, 21, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Zhao, Z.; Huang, L.; Li, L.; Li, J.; Zeng, Y.; Wu, J.; Miao, Y. Switch of the ovarian cancer cell to a calcifying phenotype in the calcification of ovarian cancer. J. Cancer 2018, 9, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Scimeca, M.; Antonacci, C.; Bonanno, E. Breast microcalcifications: A focus. J. Cell Sci. Ther. 2015, S8, e101. [Google Scholar] [CrossRef]
- Silva, E.G.; Deavers, M.T.; Parlow, A.F.; Gershenson, D.M.; Malpica, A. Calcifications in ovary and endometrium and their neoplasms. Mod. Pathol. 2003, 16, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Biomarkers | Organs | Role | Localization | Refs. |
|---|---|---|---|---|
| BMP-2 | Breast, prostate, ovary | Induces cartilage and bone formation | Vesicles, extracellular | [26,52,55,89] |
| HOXA9 | Thyroid | Cell growth and differentiation | Nucleus | [75,79] |
| OPN | Breast, ovary | Provides the basic structure of the bone matrix | Golgi apparatus, extracellular | [52,55,89] |
| PTX3 | Breast, prostate | Osteoblast proliferation, differentiation and function | Membrane, extracellular | [26,34,55] |
| RANKL | Breast, prostate | Osteoclatogenesis | Membrane, extracellular | [26] |
| RUNX2 | Breast, prostate, thyroid | Osteoblast differentiation | Nucleus, Nucleoplasm | [26,52,55,72,73] |
| VDR | Breast, prostate | Modulation of calcium levels in the body | Nucleus | [26,34,55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonfiglio, R.; Granaglia, A.; Giocondo, R.; Scimeca, M.; Bonanno, E. Molecular Aspects and Prognostic Significance of Microcalcifications in Human Pathology: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 120. https://doi.org/10.3390/ijms22010120
Bonfiglio R, Granaglia A, Giocondo R, Scimeca M, Bonanno E. Molecular Aspects and Prognostic Significance of Microcalcifications in Human Pathology: A Narrative Review. International Journal of Molecular Sciences. 2021; 22(1):120. https://doi.org/10.3390/ijms22010120
Chicago/Turabian StyleBonfiglio, Rita, Annarita Granaglia, Raffaella Giocondo, Manuel Scimeca, and Elena Bonanno. 2021. "Molecular Aspects and Prognostic Significance of Microcalcifications in Human Pathology: A Narrative Review" International Journal of Molecular Sciences 22, no. 1: 120. https://doi.org/10.3390/ijms22010120
APA StyleBonfiglio, R., Granaglia, A., Giocondo, R., Scimeca, M., & Bonanno, E. (2021). Molecular Aspects and Prognostic Significance of Microcalcifications in Human Pathology: A Narrative Review. International Journal of Molecular Sciences, 22(1), 120. https://doi.org/10.3390/ijms22010120