How to Build and to Protect the Neuromuscular Junction: The Role of the Glial Cell Line-Derived Neurotrophic Factor


Abstract
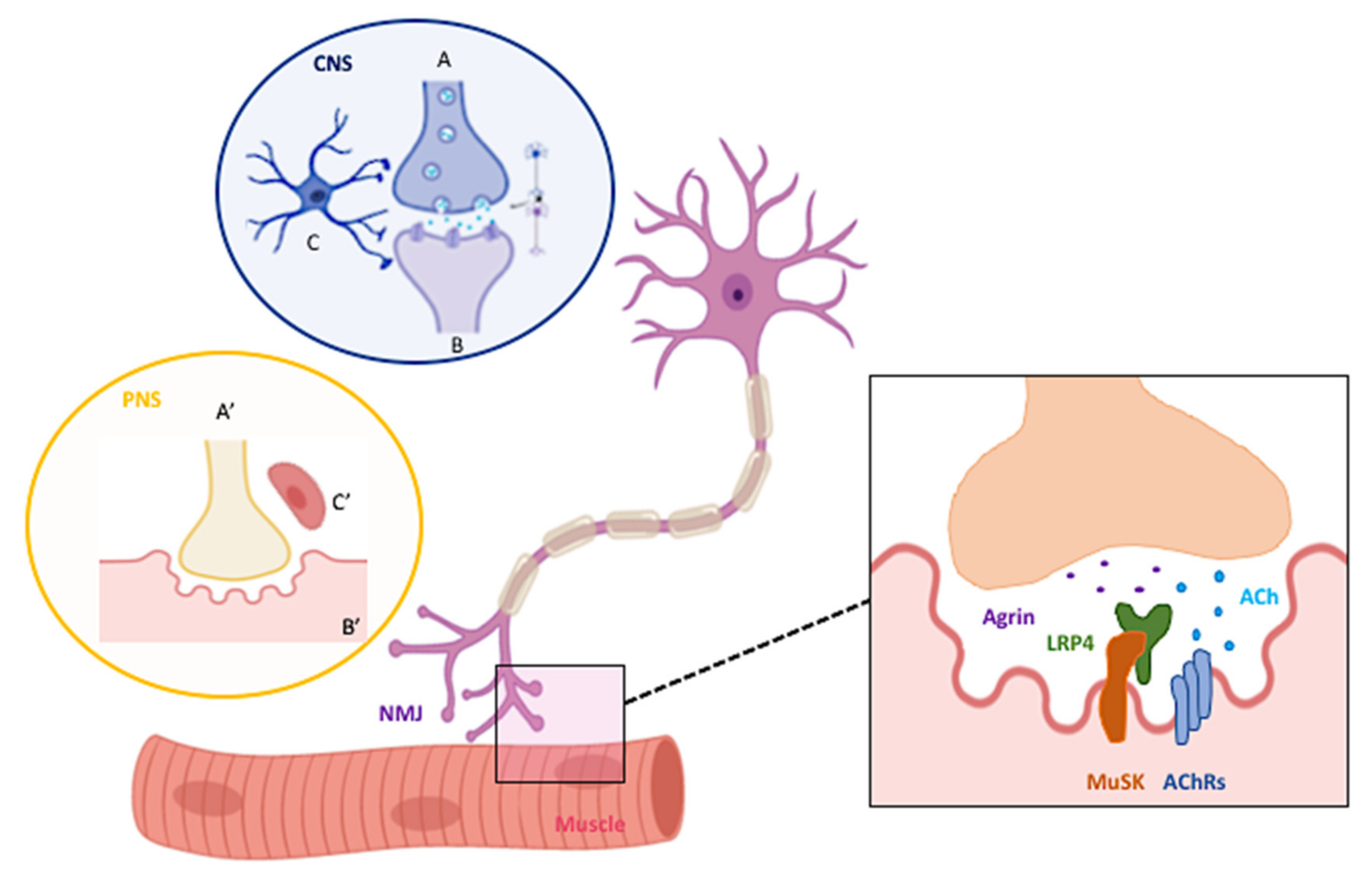
:1. NMJ Structural Architecture and Peculiarities
2. Why Is It Relevant to Study the Neuro-Muscular Interplay in Neurological Disorders?
3. Overview of the Neurotrophic Factor Support in the NS
4. The Role of GDNF at the NMJ
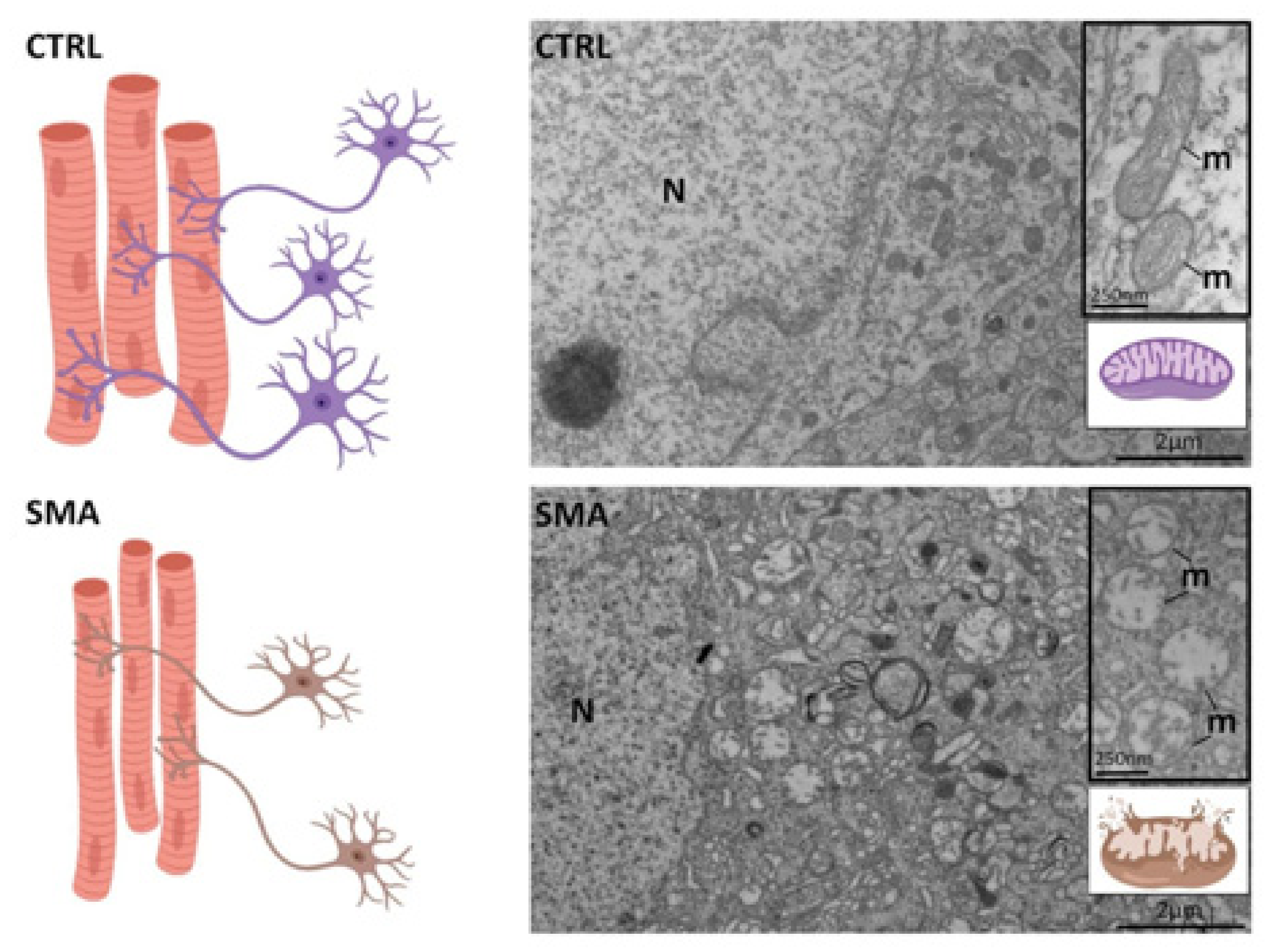
5. The NMJ-GDNF Cross-Talk: What Goes Wrong During Neurodegeneration?
6. 3D Neuromuscular Models for NMJ Reconstruction
- (i)
- 3D models performed with cell lines have the great advantage to be both handy and straightforward. Because of the dividing rate of the cells, NM contacts are rapidly obtained after 7/8 days of culture. This model is particularly suitable for studies requiring silencing and/or overexpression of specific genes in order to mimic a pathological condition. Indeed, muscle and neuronal cell lines can be efficiently transfected in co-culture models [54]. However, it is important to note that manipulation of the cells via transfection could affect itself the formation of the NM contact and, furthermore, that the transfection process is only transient.
- (ii)
- When NM contacts are performed with primary myoblasts and MNs or spinal cord slices, the timing for obtaining NM contacts is longer and can reach up to a month of culture [89]. Furthermore, a very precise coordination for animal crossings is needed in order to have pups at postnatal day P3-P5 for myoblasts cultures and, around 12–14 days after culture, embryos at E12,5 for MNs or spinal cord slices. On the other side, by working with cells/tissues derived from the animal model of interest, the disease is recapitulated without the need for further manipulation.
- (iii)
- Recently, studies based on the co-culture of human iPSCs have been implemented. The major advantage of these techniques is to perform cultures with patient-derived cells (usually fibroblasts) that are specifically reprogrammed in order to generate iPSCs to be directly differentiated into muscle or MN cells (for a comprehensive review on the topic, see [90]). Two recent works have been published regarding 3D neuromuscular models based on iPSCs. The first model, by Bakooshli and colleagues consists in mixing human muscle progenitors with human pluripotent stem cell-derived MNs [91]. The co-cultures are able to self-organize to form functional NMJ connections. Authors validated the functional connectivity by calcium imaging and electrophysiological tests, and they applied the model to the study of myasthenia gravis. The second model, by Osaki and colleagues, is based on muscle and MN interaction through a microfluid device [92]. This model needs weeks to be implemented, but the advantage is that it permits to record in real time the NMJ formation and the synchronization of MN activity and muscle contraction.
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NMJs | Neuromuscular Junctions |
| MN | Motor Neuron |
| Ach | Acetylcholine |
| AChRs | Nicotinic Receptors |
| LRP4 | Lipoprotein Receptor-Related Protein 4 |
| Musk | Muscle-Specific Kinase |
| PNS | Peripheral Nervous System |
| CNS | Central Nervous System |
| NFs | Neurotrophic Factors |
| NGF | Nerve Growth Factor |
| BDNF | Brain-Derived Neurotrophic Factor |
| NT-3 | Neurotrophin-3 |
| NT-4 | Neurotrophin-4 |
| TGF-β | Transforming Growth Factor- Β |
| BMP | Bone Morphogenetic Protein |
| CNTF | Ciliary Neurotrophic Factor |
| LIF | Leukemia Inhibitory Factor |
| IL-6 | Interleukin-6 |
| IL-11 | Interleukin-11 |
| CTF1 | Cardio-Trophin-1 |
| FGF1 | Fibroblast Growth Factor 1 |
| FGF2 | Fibroblast Growth Factor 2 |
| EGF | Epidermal Growth Factor |
| IGF1 | Insulin-Like Growth Factor 1 |
| GDNF | Glial-Cell-Line-Derived Neurotrophic Factor |
| GFRα1 | GDNF family receptor alpha-1 |
| RET | REarranged during Transfection |
| NCAM | Neural Cell Adhesion Molecule |
| AD | Alzheimer’s Disease |
| ALS | Amyotrophic Lateral Sclerosis |
| SMA | Spinal Muscular Atrophy |
| APP | Amyloid Precursor Protein |
| APLP1 | Amyloid Precursor-Like Protein1 |
| APLP2 | Amyloid Precursor-Like Protein2 |
| PSs | Presenilins |
| iPSCs | Human Induced Pluripotent Stem |
| CSF | Cerebrospinal Fluid |
| FDA | Food and Drug Administration |
References
- Guarino, S.R.; Canciani, A.; Forneris, F. Dissecting the Extracellular Complexity of Neuromuscular Junction Organizers. Front. Mol. Biosci. 2020, 6, 156. [Google Scholar] [CrossRef] [PubMed]
- Bayati, A.; Berman, T. Localized vs. Systematic Neurodegeneration: A Paradigm Shift in Understanding Neurodegenerative Diseases. Front. Syst. Neurosci. 2017, 11, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, X.; Castro-Obregón, S.; Gutiérrez-Becker, B.; Gutiérrez-Ospina, G.; Karalis, N.; Khalil, A.A.; Lopez-Noguerola, J.S.; Rodríguez, L.L.; Martínez-Martínez, E.; Perez-Cruz, C.; et al. Re-thinking the Etiological Framework of Neurodegeneration. Front. Neurosci. 2019, 13, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, A.; Sethumadhavan, A.; Krishnan, U.M. Toward Building the Neuromuscular Junction: In Vitro Models to Study Synaptogenesis and Neurodegeneration. ACS Omega 2019, 4, 12969–12977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanni, C.; Stanga, S.; Racchi, M.; Govoni, S. The expanding universe of neurotrophic factors: Therapeutic potential in aging and age-associated disorders. Curr. Pharm. Des. 2010, 16, 698–717. [Google Scholar] [CrossRef] [Green Version]
- Bartus, R.T.; Johnson, E.M., Jr. Clinical tests of neurotrophic factors for human neurodegenerative diseases, part 1: Where have we been and what have we learned? Neurobiol. Dis. 2017, 97, 156–168. [Google Scholar] [CrossRef]
- Levy, Y.S.; Gilgun-Sherki, Y.; Melamed, E.; Offen, D. Therapeutic Potential of Neurotrophic Factors in Neurodegenerative Diseases. BioDrugs 2005, 19, 97–127. [Google Scholar] [CrossRef]
- Keller, G.B.; Barde, Y.-A. Signalling through the neurotrophin receptor p75NTR. Curr. Opin. Neurobiol. 1997, 7, 413–418. [Google Scholar] [CrossRef]
- Hamburger, V.; Levi-Montalcini, R. Proliferation, differentiation and degeneration in the spinal ganglia of the chick embryo under normal and experimental conditions. J. Exp. Zool. 1949, 111, 457–501. [Google Scholar] [CrossRef]
- Hellweg, R.; Von Richthofen, S.; Anders, D.; Baethge, C.; Röpke, S.; Hartung, H.-D.; Gericke, C.A. The time course of nerve growth factor content in different neuropsychiatric diseases—A unifying hypothesis. J. Neural Transm. 1998, 105, 871–903. [Google Scholar] [CrossRef]
- Hofer, M.; Pagliusi, S.R.; Hohn, A.; Leibrock, J.; Barde, Y.A. Regional distribution of brain-derived neurotrophic factor mRNA in the adult mouse brain. EMBO J. 1990, 9, 2459–2464. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Cattaneo, E. Brain-derived neurotrophic factor in neurodegenerative diseases. Nat. Rev. Neurol. 2009, 5, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, K.; Jasmin, B.J. BDNF Is Expressed in Skeletal Muscle Satellite Cells and Inhibits Myogenic Differentiation. J. Neurosci. 2006, 26, 5739–5749. [Google Scholar] [CrossRef] [Green Version]
- Li, M.O.; Flavell, R.A. TGF-β: A Master of All T Cell Trades. Cell 2008, 134, 392–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landreth, G.E. Growth factors. In Basic Neurochemistry, Molecular, Cellular and Medical Aspects, 6th ed.; Siegel, G.J., Agranoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1999; pp. 384–396. [Google Scholar]
- Horton, A.R.; Bartlett, P.F.; Pennica, D.; Davies, A.M. Cytokines promote the survival of mouse cranial sensory neurones at different developmental stages. Eur. J. Neurosci. 1998, 10, 673–679. [Google Scholar] [CrossRef]
- Curtis, R.; Adryan, K.M.; Zhu, Y.; Harkness, P.J.; Lindsay, R.M.; Distefano, P.S. Retrograde axonal transport of ciliary neurotrophic factor is increased by peripheral nerve injury. Nat. Cell Biol. 1993, 365, 253–255. [Google Scholar] [CrossRef]
- Quesada, A.; Romeo, H.E.; Micevych, P.E. Distribution and localization patterns of estrogen receptor-β and insulin-like growth factor-1 receptors in neurons and glial cells of the female rat substantia nigra: Localization of ERβ and IGF-1R in substantia nigra. J. Comp. Neurol. 2007, 503, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Runeberg-Roos, P.; Penn, R.D. Improving therapeutic potential of GDNF family ligands. Cell Tissue Res. 2020, 382, 173–183. [Google Scholar] [CrossRef]
- Rosenblad, C.; Kirik, D.; Devaux, B.; Moffat, B.; Phillips, H.S.; Björklund, A. Protection and regeneration of nigral dopaminergic neurons by neurturin or GDNF in a partial lesion model of Parkinson’s disease after administration into the striatum or the lateral ventricle. Eur. J. Neurosci. 1999, 11, 1554–1566. [Google Scholar] [CrossRef] [Green Version]
- Baloh, R.H.; Tansey, M.G.; Lampe, P.A.; Fahrner, T.J.; Enomoto, H.; Simburger, K.S.; Leitner, M.L.; Araki, T.; Johnson, E.M.; Milbrandt, J. Artemin, a Novel Member of the GDNF Ligand Family, Supports Peripheral and Central Neurons and Signals through the GFRα3–RET Receptor Complex. Neuron 1998, 21, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.F.; Doherty, D.H.; Lile, J.D.; Bektesh, S.; Collins, F. GDNF: A glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science 1993, 260, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Marks, W.J.; Bartus, R.T.; Siffert, J.; Davis, C.S.; Lozano, A.; Boulis, N.; Vitek, J.; Stacy, M.; Turner, D.; Verhagen, L.; et al. Gene delivery of AAV2-neurturin for Parkinson’s disease: A double-blind, randomised, controlled trial. Lancet Neurol. 2010, 9, 1164–1172. [Google Scholar] [CrossRef]
- Paratcha, G.; Ledda, F.; Baars, L.; Coulpier, M.; Besset, V.; Anders, J.; Scott, R.P.; Ibáñez, C.F. Released GFRα1 Potentiates Downstream Signaling, Neuronal Survival, and Differentiation via a Novel Mechanism of Recruitment of c-Ret to Lipid Rafts. Neuron 2001, 29, 171–184. [Google Scholar] [CrossRef]
- Baudet, C.; Ernfors, P. Soluble and bound forms of GFR?1 elicit different GDNF-independent neurite growth responses in primary sensory neurons. Dev. Dyn. 2003, 227, 27–34. [Google Scholar] [CrossRef]
- Wang, X. Structural studies of GDNF family ligands with their receptors—Insights into ligand recognition and activation of receptor tyrosine kinase RET. Biochim. Biophys. Acta (BBA) 2013, 1834, 2205–2212. [Google Scholar] [CrossRef] [PubMed]
- Paratcha, G.; Ledda, F.; Ibáñez, C.F. The Neural Cell Adhesion Molecule NCAM Is an Alternative Signaling Receptor for GDNF Family Ligands. Cell 2003, 113, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, M.M.; Sidorova, Y.A.; Tumova, S.; Ahonen-Bishopp, A.; Magalhães, A.C.; Kulesskiy, E.; Paveliev, M.; Rivera, C.; Rauvala, H.; Saarma, M. Heparan sulfate proteoglycan syndecan-3 is a novel receptor for GDNF, neurturin, and artemin. J. Cell Biol. 2011, 192, 153–169. [Google Scholar] [CrossRef] [Green Version]
- Springer, J.; Mu, X.; Bergmann, L.; Trojanowski, J. Expression of GDNF mRNA in Rat and Human Nervous Tissue. Exp. Neurol. 1994, 127, 167–170. [Google Scholar] [CrossRef]
- Cortés, D.; Robledo-Arratia, Y.; Hernández-Martínez, R.; Escobedo-Ávila, I.; Bargas, J.; Velasco, I. Transgenic GDNF Positively Influences Proliferation, Differentiation, Maturation and Survival of Motor Neurons Produced from Mouse Embryonic Stem Cells. Front. Cell. Neurosci. 2016, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Hellmich, H.L.; Kos, L.; Cho, E.S.; Mahon, K.A.; Zimmer, A. Embryonic expression of glial cell-line derived neurotrophic factor (GDNF) suggests multiple developmental roles in neural differentiation and epithelial-mesenchymal interactions. Mech. Dev. 1996, 54, 95–105. [Google Scholar] [CrossRef]
- Huang, L.; Guo, H.; Hellard, D.; Katz, D. Glial cell line-derived neurotrophic factor (GDNF) is required for differentiation of pontine noradrenergic neurons and patterning of central respiratory output. Neuroscience 2005, 130, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Oo, T.F.; Ries, V.; Cho, J.; Kholodilov, N.; Burke, R.E. Anatomical basis of glial cell line-derived neurotrophic factor expression in the striatum and related basal ganglia during postnatal development of the rat. J. Comp. Neurol. 2005, 484, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buss, R.R.; Gould, T.W.; Ma, J.; Vinsant, S.; Prevette, D.; Winseck, A.; Toops, K.A.; Hammarback, J.A.; Smith, T.L.; Oppenheim, R.W. Neuromuscular Development in the Absence of Programmed Cell Death: Phenotypic Alteration of Motoneurons and Muscle. J. Neurosci. 2006, 26, 13413–13427. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, A.; Baltazar, G.; Duarte, E.P. Driving GDNF expression: The green and the red traffic lights. Prog. Neurobiol. 2008, 86, 186–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotoyama, H.; Iwakura, Y.; Oda, K.; Sasaoka, T.; Takei, N.; Kakita, A.; Enomoto, H.; Nawa, H. Striatal hypodopamine phenotypes found in transgenic mice that overexpress glial cell line-derived neurotrophic factor. Neurosci. Lett. 2017, 654, 99–106. [Google Scholar] [CrossRef]
- Pozas, E.; Ibáñez, C.F. GDNF and GFRα1 Promote Differentiation and Tangential Migration of Cortical GABAergic Neurons. Neuron 2005, 45, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Ernsberger, U. The role of GDNF family ligand signalling in the differentiation of sympathetic and dorsal root ganglion neurons. Cell Tissue Res. 2008, 333, 353–371. [Google Scholar] [CrossRef] [Green Version]
- Bär, K.J.; Saldanha, G.J.F.; Kennedy, A.J.; Facer, P.; Birch, R.; Carlstedt, T.; Anand, P. GDNF and its receptor component Ret in injured human nerves and dorsal root ganglia. Neuroreport 1998, 9, 43–47. [Google Scholar] [CrossRef]
- Trupp, M.; Rydén, M.; Jörnvall, H.; Funakoshi, H.; Timmusk, T.; Arenas, E.; Ibáñez, C.F. Peripheral expression and biological activities of GDNF, a new neurotrophic factor for avian and mammalian peripheral neurons. J. Cell Biol. 1995, 130, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Hase, A.; Miyata, Y.; Arahata, K.; Akazawa, C. Prominent expression of glial cell line-derived neurotrophic factor in human skeletal muscle. J Comp. Neurol. 1998, 402, 303–312. [Google Scholar] [CrossRef]
- Nagano, M.; Suzuki, H. Quantitative analyses of expression of GDNF and neurotrophins during postnatal development in rat skeletal muscles. Neurosci. Res. 2003, 45, 391–399. [Google Scholar] [CrossRef]
- Zahavi, E.E.; Ionescu, A.; Gluska, S.; Gradus, T.; Ben-Yaakov, K.; Perlson, E. A compartmentalized microfluidic neuromuscular co-culture system reveals spatial aspects of GDNF functions. J. Cell Sci. 2015, 128, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, D.; Giudetti, G.; Micera, S.; Navarro, X.; Del Valle, J. Focal release of neurotrophic factors by biodegradable microspheres enhance motor and sensory axonal regeneration in vitro and in vivo. Brain Res. 2016, 1636, 93–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, M.; Zhuang, H.; Bu, S.; Hua, L.; Darabi, M.A.; Cao, X. Gelatin-methacrylamide gel loaded with microspheres to deliver GDNF in bilayer collagen conduit promoting sciatic nerve growth. Int. J. Nanomed. 2016, 11, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höke, A.; Gordon, T.; Zochodne, D.; Sulaiman, O. A Decline in Glial Cell-Line-Derived Neurotrophic Factor Expression Is Associated with Impaired Regeneration after Long-Term Schwann Cell Denervation. Exp. Neurol. 2002, 173, 77–85. [Google Scholar] [CrossRef]
- Walker, M.J.; Xu, X.-M. History of Glial Cell Line-Derived Neurotrophic Factor (GDNF) and Its Use for Spinal Cord Injury Repair. Brain Sci. 2018, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, M.D.; Sander, S.; Tenenbaum, L. GDNF, A Neuron-Derived Factor Upregulated in Glial Cells during Disease. J. Clin. Med. 2020, 9, 456. [Google Scholar] [CrossRef] [Green Version]
- Baudet, C.; Pozas, E.; Adameyko, I.; Andersson, E.; Ericson, J.; Ernfors, P. Retrograde Signaling onto Ret during Motor Nerve Terminal Maturation. J. Neurosci. 2008, 28, 963–975. [Google Scholar] [CrossRef] [Green Version]
- Oppenheim, R.W.; Houenou, L.J.; Johnson, J.E.; Lin, L.-F.H.; Li, L.; Lo, A.C.; Newsome, A.L.; Prevette, D.M.; Wang, S. Developing motor neurons rescued from programmed and axotomy-induced cell death by GDNF. Nat. Cell Biol. 1995, 373, 344–346. [Google Scholar] [CrossRef]
- Wang, Y.; Gupta, A.; Toledo-Rodriguez, M.; Wu, C.Z.; Markram, H. Anatomical, Physiological, Molecular and Circuit Properties of Nest Basket Cells in the Developing Somatosensory Cortex. Cereb. Cortex 2002, 12, 395–410. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Parsadanian, A.S.; Snider, W.D.; Lichtman, J.W. Hyperinnervation of Neuromuscular Junctions Caused by GDNF Overexpression in Muscle. Science 1998, 279, 1725–1729. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Yang, F.; He, X.-P.; Je, H.S.; Zhou, J.-Z.; Eckermann, K.; Kawamura, D.; Feng, L.; Shen, L.; Lu, B. Regulation of Neuromuscular Synapse Development by Glial Cell Line-derived Neurotrophic Factor and Neurturin. J. Biol. Chem. 2002, 277, 10614–10625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanga, S.; Zanou, N.; Audouard, E.; Tasiaux, B.; Contino, S.; Vandermeulen, G.; René, F.; Loeffler, J.; Clotman, F.; Gailly, P.; et al. APP-dependent glial cell line-derived neurotrophic factor gene expression drives neuromuscular junction formation. FASEB J. 2015, 30, 1696–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller-Peck, C.R.; Feng, G.; Sanes, J.R.; Yan, Q.; Lichtman, J.W.; Snider, W.D. Glial Cell Line-Derived Neurotrophic Factor Administration in Postnatal Life Results in Motor Unit Enlargement and Continuous Synaptic Remodeling at the Neuromuscular Junction. J. Neurosci. 2001, 21, 6136–6146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwick, M.; Teng, L.; Mu, X.; Springer, J.E.; Davis, B.M. Overexpression of GDNF Induces and Maintains Hyperinnervation of Muscle Fibers and Multiple End-Plate Formation. Exp. Neurol. 2001, 171, 342–350. [Google Scholar] [CrossRef]
- Wehrwein, E.A.; Roskelley, E.M.; Spitsbergen, J.M. GDNF is regulated in an activity-dependent manner in rat skeletal muscle. Muscle Nerve 2002, 26, 206–211. [Google Scholar] [CrossRef] [PubMed]
- McCullough, M.B.; Domire, Z.J.; Reed, A.M.; Amin, S.; Ytterberg, S.R.; Chen, Q.; An, K.-N. Evaluation of muscles affected by myositis using magnetic resonance elastography. Muscle Nerve 2011, 43, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Gyorkos, A.M.; Spitsbergen, J.M. GDNF content and NMJ morphology are altered in recruited muscles following high-speed and resistance wheel training. Physiol. Rep. 2014, 2, e00235. [Google Scholar] [CrossRef]
- Stanga, S.; Caretto, A.; Boido, M.; Vercelli, A. Mitochondrial Dysfunctions: A Red Thread across Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 3719. [Google Scholar] [CrossRef]
- Qing, J.; Liu, X.; Wu, Q.; Zhou, M.; Zhang, Y.; Mazhar, M.; Huang, X.; Wang, L.; He, F. Hippo/YAP Pathway Plays a Critical Role in Effect of GDNF Against Aβ-Induced Inflammation in Microglial Cells. DNA Cell Biol. 2020, 39, 1064–1071. [Google Scholar] [CrossRef]
- Pöyhönen, S.; Er, S.; Domanskyi, A.; Airavaara, M. Effects of Neurotrophic Factors in Glial Cells in the Central Nervous System: Expression and Properties in Neurodegeneration and Injury. Front. Physiol. 2019, 10, 486. [Google Scholar] [CrossRef]
- Choi, H.Y.; Liu, Y.; Tennert, C.; Sugiura, Y.; Karakatsani, A.; Kröger, S.; Johnson, E.B.; Hammer, R.E.; Lin, W.; Herz, J. APP interacts with LRP4 and agrin to coordinate the development of the neuromuscular junction in mice. eLife 2013, 2, e00220. [Google Scholar] [CrossRef]
- Lin, M.; Xiong, W.-C.; Mei, L. Neuromuscular Junction Formation, Aging, and Disorders. Annu. Rev. Physiol. 2018, 80, 159–188. [Google Scholar] [CrossRef]
- De Strooper, B.; Saftig, P.; Craessaerts, K.; Vanderstichele, H.; Guhde, G.; Annaert, W.; Von Figura, K.; Van Leuven, F. Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nat. Cell Biol. 1998, 391, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Hage, S.; Marinangeli, C.; Stanga, S.; Octave, J.-N.; Quetin-Leclercq, J.; Kienlen-Campard, P. Gamma-Secretase Inhibitor Activity of a Pterocarpus erinaceus Extract. Neurodegener. Dis. 2014, 14, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Hage, S.; Stanga, S.; Marinangeli, C.; Octave, J.-N.; Dewachter, I.; Quetin-Leclercq, J.; Kienlen-Campard, P. Characterization of Pterocarpus erinaceus kino extract and its gamma-secretase inhibitory properties. J. Ethnopharmacol. 2015, 163, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Stanga, S.; Vrancx, C.; Tasiaux, B.; Marinangeli, C.; Karlström, H.; Kienlen-Campard, P. Specificity of presenilin-1- and presenilin-2-dependent γ-secretases towards substrate processing. J. Cell. Mol. Med. 2017, 22, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Contino, S.; Porporato, P.E.; Bird, M.; Marinangeli, C.; Opsomer, R.; Sonveaux, P.; Bontemps, F.; Dewachter, I.; Octave, J.-N.; Bertrand, L.; et al. Presenilin 2-Dependent Maintenance of Mitochondrial Oxidative Capacity and Morphology. Front. Physiol. 2017, 8, 796. [Google Scholar] [CrossRef] [Green Version]
- Hardiman, O.; Berg, L.H.V.D.; Kiernan, M.C. Clinical diagnosis and management of amyotrophic lateral sclerosis. Nat. Rev. Neurol. 2011, 7, 639–649. [Google Scholar] [CrossRef]
- Kolb, S.J.; Kissel, J.T. Spinal Muscular Atrophy. Neurol. Clin. 2015, 33, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; McHugh, J.; Tork, C.; Shelley, B.; Hayes, A.; Bellantuono, I.; Aebischer, P.; Svendsen, C.N. Direct Muscle Delivery of GDNF With Human Mesenchymal Stem Cells Improves Motor Neuron Survival and Function in a Rat Model of Familial ALS. Mol. Ther. 2008, 16, 2002–2010. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, M.H.; Figlewicz, D.A.; Bohn, M.C. Intramuscular Grafts of Myoblasts Genetically Modified to Secrete Glial Cell Line-Derived Neurotrophic Factor Prevent Motoneuron Loss and Disease Progression in a Mouse Model of Familial Amyotrophic Lateral Sclerosis. Hum. Gene Ther. 1999, 10, 1853–1866. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, G.M.; Alkaslasi, M.; Vit, J.-P.; Lawless, G.; Godoy, M.; Gowing, G.; Shelest, O.; Svendsen, C.N. Systemic injection of AAV9-GDNF provides modest functional improvements in the SOD1G93A ALS rat but has adverse side effects. Gene Ther. 2017, 24, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koistinen, H.; Prinjha, R.; Soden, P.; Harper, A.; Banner, S.J.; Dingwall, C.; Pradat, P.-F.; Loeffler, J.-P. Elevated levels of amyloid precursor protein in muscle of patients with amyotrophic lateral sclerosis and a mouse model of the disease. Muscle Nerve 2006, 34, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Bryson, J.B.; Hobbs, C.; Parsons, M.J.; Bosch, K.D.; Pandraud, A.; Walsh, F.S.; Doherty, P.; Greensmith, L. Amyloid precursor protein (APP) contributes to pathology in the SOD1G93A mouse model of amyotrophic lateral sclerosis. Hum. Mol. Genet. 2012, 21, 3871–3882. [Google Scholar] [CrossRef] [Green Version]
- Stanga, S.; Brambilla, L.; Tasiaux, B.; Dang, A.H.; Ivanoiu, A.; Octave, J.-N.; Rossi, D.; Van Pesch, V.; Kienlen-Campard, P. A Role for GDNF and Soluble APP as Biomarkers of Amyotrophic Lateral Sclerosis Pathophysiology. Front. Neurol. 2018, 9, 384. [Google Scholar] [CrossRef]
- Menduti, G.; Rasà, D.M.; Stanga, S.; Boido, M. Drug Screening and Drug Repositioning as Promising Therapeutic Approaches for Spinal Muscular Atrophy Treatment. Front. Pharmacol. 2020, 11, 592234. [Google Scholar] [CrossRef]
- Boido, M.; Vercelli, A. Neuromuscular Junctions as Key Contributors and Therapeutic Targets in Spinal Muscular Atrophy. Front. Neuroanat. 2016, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Miller, N.; Shi, H.; Zelikovich, A.S.; Ma, Y.-C. Motor neuron mitochondrial dysfunction in spinal muscular atrophy. Hum. Mol. Genet. 2016, 25, 3395–3406. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Diaz, C.; Nicole, S.; Velasco, M.E.; Borra-Cebrian, C.; Panozzo, C.; Frugier, T.; Millet, G.; Roblot, N.; Joshi, V.; Melki, J. Neurofilament accumulation at the motor endplate and lack of axonal sprouting in a spinal muscular atrophy mouse model. Hum. Mol. Genet. 2002, 11, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Kariya, S.; Park, G.-H.; Maeno-Hikichi, Y.; Leykekhman, O.; Lutz, C.; Arkovitz, M.S.; Landmesser, L.T.; Monani, U.R. Reduced SMN protein impairs maturation of the neuromuscular junctions in mouse models of spinal muscular atrophy. Hum. Mol. Genet. 2008, 17, 2552–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valsecchi, V.; Boido, M.; De Amicis, E.; Piras, A.; Vercelli, A. Expression of Muscle-Specific MiRNA 206 in the Progression of Disease in a Murine SMA Model. PLoS ONE 2015, 10, e0128560. [Google Scholar] [CrossRef] [PubMed]
- McGovern, V.L.; Gavrilina, T.O.; Beattie, C.E.; Burghes, A.H. Embryonic motor axon development in the severe SMA mouse. Hum. Mol. Genet. 2008, 17, 2900–2909. [Google Scholar] [CrossRef] [Green Version]
- Murray, L.M.; Comley, L.H.; Thomson, D.; Parkinson, N.; Talbot, K.; Gillingwater, T.H. Selective vulnerability of motor neurons and dissociation of pre- and post-synaptic pathology at the neuromuscular junction in mouse models of spinal muscular atrophy. Hum. Mol. Genet. 2007, 17, 949–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Benito, L.; Ruiz, R.; Tabares, L. Synaptic defects in spinal muscular atrophy animal models. Dev. Neurobiol. 2012, 72, 126–133. [Google Scholar] [CrossRef]
- McGivern, J.V.; Patitucci, T.N.; Nord, J.A.; Barabas, M.-E.A.; Stucky, C.L.; Ebert, A.D. Spinal muscular atrophy astrocytes exhibit abnormal calcium regulation and reduced growth factor production. Glia 2013, 61, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Chiaretti, A.; Leoni, C.; Barone, G.; Genovese, O.; Brahe, C.; Mariotti, P.; Conti, G. Increased Levels of Glial Cell-Derived Neurotrophic Factor in CSF of Infants With SMA. Pediatr. Neurol. 2009, 41, 195–199. [Google Scholar] [CrossRef]
- Audouard, E.; Schakman, O.; René, F.; Huettl, R.-E.; Huber, A.B.; Loeffler, J.-P.; Gailly, P.; Clotman, F. The Onecut Transcription Factor HNF-6 Regulates in Motor Neurons the Formation of the Neuromuscular Junctions. PLoS ONE 2012, 7, e50509. [Google Scholar] [CrossRef] [Green Version]
- Patrón, L.A.; August, P.R. Modeling neuromuscular junctions in vitro: A review of the current progress employing human induced pluripotent stem cells. AIMS Cell Tissue Eng. 2017, 2, 91–118. [Google Scholar] [CrossRef]
- Bakooshli, M.A.; Lippmann, E.S.; Mulcahy, B.; Iyer, N.; Nguyen, C.T.; Tung, K.; Stewart, B.A.; van den Dorpel, H.; Fuehrmann, T.; Shoichet, M.; et al. A 3D culture model of innervated human skeletal muscle enables studies of the adult neuromuscular junction. eLife 2019, 8, e44530. [Google Scholar] [CrossRef]
- Osaki, T.; Uzel, S.G.M.; Kamm, R. On-chip 3D neuromuscular model for drug screening and precision medicine in neuromuscular disease. Nat. Protoc. 2020, 15, 421–449. [Google Scholar] [CrossRef] [PubMed]
- Lanni, C.; Racchi, M.; Stanga, S.; Mazzini, G.; Ranzenigo, A.; Polotti, R.; Memo, M.; Govoni, S.; Uberti, D. Unfolded p53 in Blood as a Predictive Signature Signature of the Transition from Mild Cognitive Impairment to Alzheimer’s Disease. J. Alzheimer’s Dis. 2010, 20, 97–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanga, S.; Lanni, C.; Govoni, S.; Uberti, D.; D’Orazi, G.; Racchi, M. Unfolded p53 in the pathogenesis of Alzheimer’s disease: Is HIPK2 the link? Aging 2010, 2, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanga, S.; Lanni, C.; Sinforiani, E.; Mazzini, G.; Racchi, M. Searching for Predictive Blood Biomarkers: Misfolded p53 In Mild Cognitive Impairment. Curr. Alzheimer Res. 2012, 9, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Summers, M.J.; Rainero, I.; Vercelli, A.E.; Aumayr, G.; De Rosario, H.; Mönter, M.; Kawashima, R.; Caglio, M.; Carbone, C.; Rubino, E.; et al. The My Active and Healthy Aging (My-AHA) ICT platform to detect and prevent frailty in older adults: Randomized control trial design and protocol. Alzheimer’s Dementia: Transl. Res. Clin. Interv. 2018, 4, 252–262. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Superfamilies | NFs Families | Submembers | Families’ Major Functions | Expression in Peripheral Tissues |
|---|---|---|---|---|
| NGF | NGF BDNF NTF3 NTF4 | NS development, Neuron survival or apoptosis and cell death, Neurite outgrowth | Skeletal muscle + | |
| TGF-β | GDNF | |||
| Neurturin | ||||
| Artemin | NS development, | Skeletal muscle ++ | ||
| GDNF family | Persephin | NMJ formation and maintenance | Schwann cells ++ | |
| TGFβ family | ||||
| BMP family | TGFβ1-3 | NS development | Immune system ++ | |
| Schwann cells + | ||||
| BMP 1-6, 71, 8a, 8b, 10, 15 | Bone formation | Bone ++ | ||
| Neurokine or Neuropoietin | CNTF LIF IL-6 IL-11 CTF1 Oncostatin-M Granulocyte colony-stimulating factor Cardiotrophin-1 | NS development, Regulation of neurotransmitter phenotype, Neurons’ rescue from axotomy-induced cell death | Skeletal muscle + | |
| Non-Neuronal Growth Factors | FGF1 FGF2 EGF IGF1 | NS survival | Liver ++ Bone + Cartilage + Skeletal muscle + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanga, S.; Boido, M.; Kienlen-Campard, P. How to Build and to Protect the Neuromuscular Junction: The Role of the Glial Cell Line-Derived Neurotrophic Factor. Int. J. Mol. Sci. 2021, 22, 136. https://doi.org/10.3390/ijms22010136
Stanga S, Boido M, Kienlen-Campard P. How to Build and to Protect the Neuromuscular Junction: The Role of the Glial Cell Line-Derived Neurotrophic Factor. International Journal of Molecular Sciences. 2021; 22(1):136. https://doi.org/10.3390/ijms22010136
Chicago/Turabian StyleStanga, Serena, Marina Boido, and Pascal Kienlen-Campard. 2021. "How to Build and to Protect the Neuromuscular Junction: The Role of the Glial Cell Line-Derived Neurotrophic Factor" International Journal of Molecular Sciences 22, no. 1: 136. https://doi.org/10.3390/ijms22010136
APA StyleStanga, S., Boido, M., & Kienlen-Campard, P. (2021). How to Build and to Protect the Neuromuscular Junction: The Role of the Glial Cell Line-Derived Neurotrophic Factor. International Journal of Molecular Sciences, 22(1), 136. https://doi.org/10.3390/ijms22010136