Endoglin Targeting: Lessons Learned and Questions That Remain



Abstract
:1. Introduction
1.1. Introduction and Endoglin History
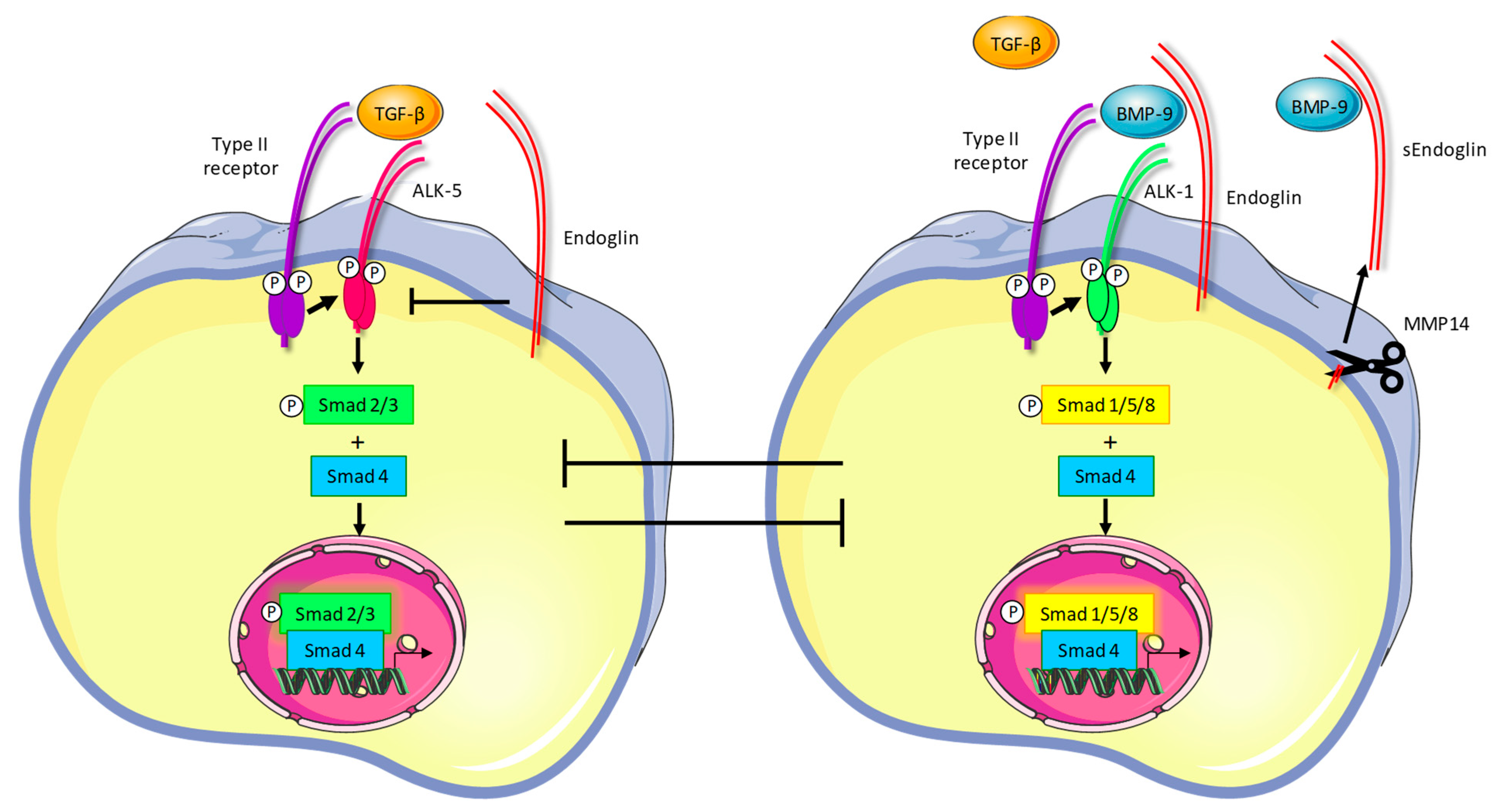
1.2. Endoglin Signaling
1.3. Targeting Endoglin for Imaging and Therapy
2. Preclinical Studies on Endoglin Targeting
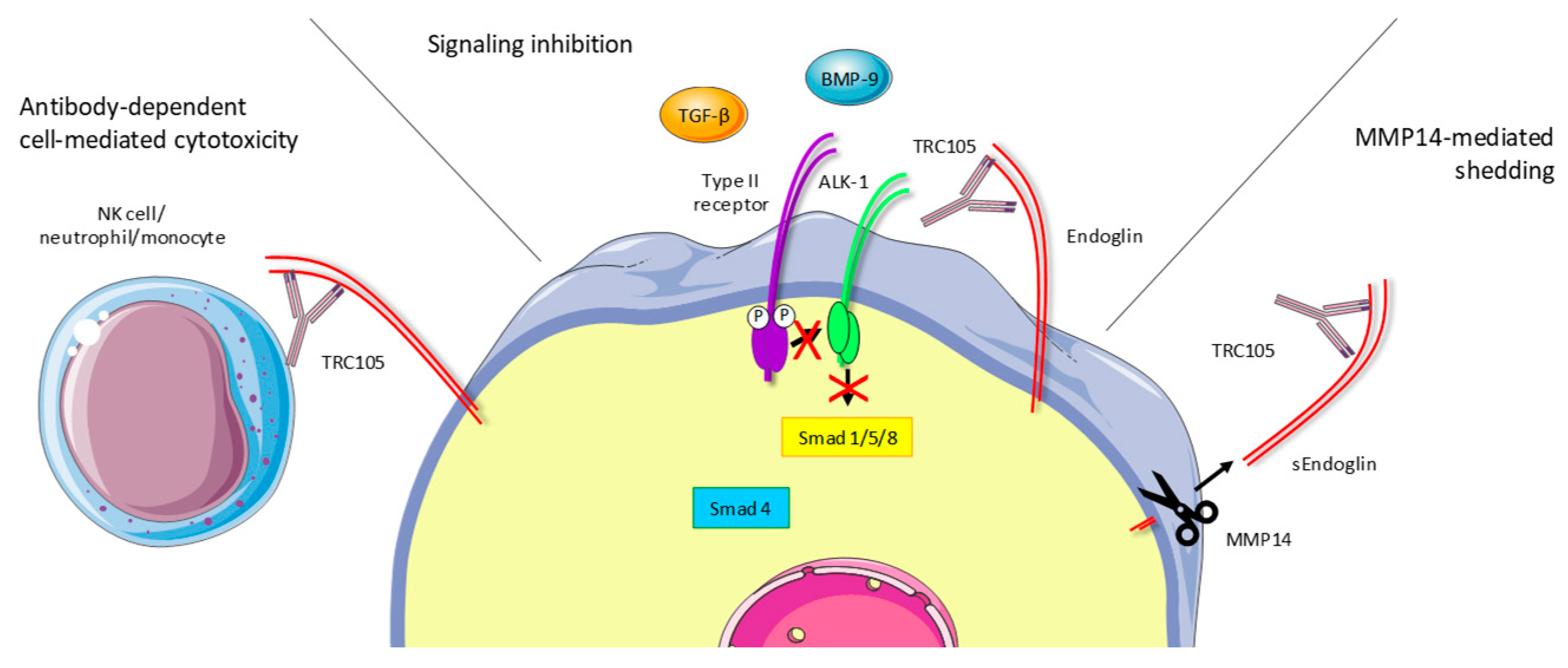
2.1. Development of the Anti-Endoglin Antibody Sn6j/TRC105
2.2. TRC105 Acts Through Immune-Dependent Mechanisms
2.3. TRC105 Inhibits BMP-9-Induced Signaling
2.4. TRC105 and Crosstalk with the Vascular Endothelial Growth Factor (VEGF) Pathway
2.5. TRC105 and Anti-Tumor Effects in Preclinical Cancer Models
3. TRC105 Clinical Trials
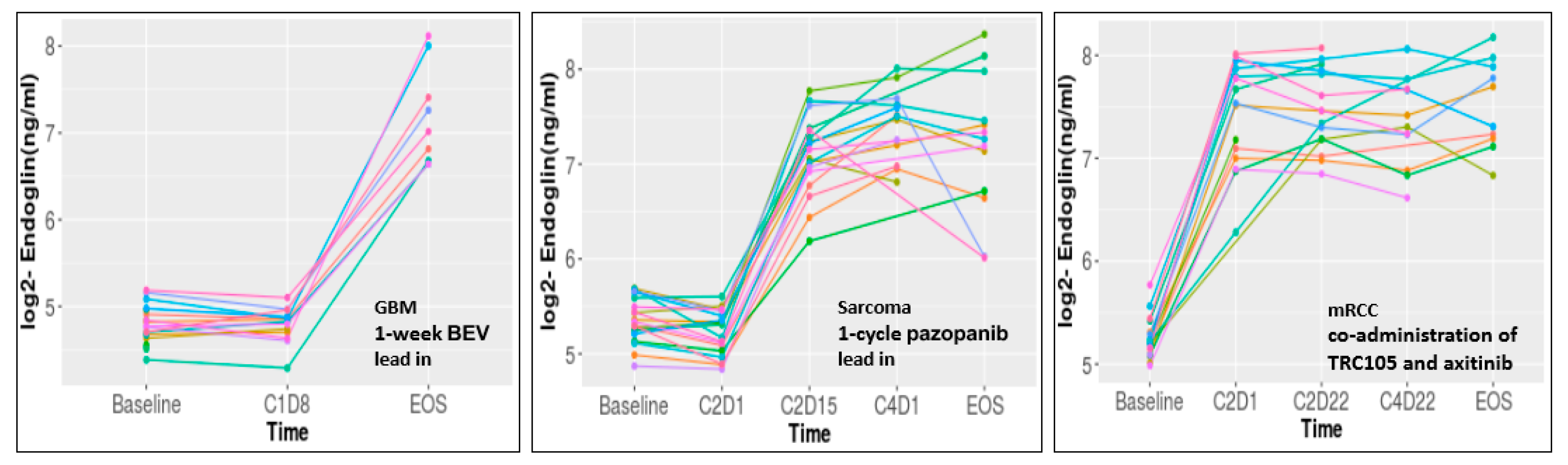
3.1. Biomarker Findings in Clinical Trials
3.2. TRC105 First-in-Human Clinical Trial
3.3. Crosstalk with the VEGF Pathway: TRC105 Plus VEGF Inhibitors in Phase Ib Trials
3.4. TRC105 Plus VEGF Inhibitors in a Series of Phase Ib Trials
3.4.1. Glioblastoma (NCT01648348)
3.4.2. Sarcoma (NCT01975519)
3.4.3. Renal Cell Carcinoma (NCT01806064)
3.4.4. TRC105 and Capecitabine in Breast Cancer Patients (NCT01326481)
3.5. TRC105 Biomarkers in Randomized Trials
3.6. Biomarker Conclusions
4. Discussion
4.1. Lessons Learned, Question to Be Answered
4.2. Looking Forward
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP | Bone morphogenetic protein |
| sEng | Soluble endoglin |
| ICAM-1 | Inter-cellular adhesion molecule 1 |
| OPN | Osteopontin |
| PDGF | Platelet-derived growth factor |
| PlGF | Placenta growth factor |
| TGFβ | Transforming growth factor-β |
| TSP2 | Thrombospondin 2 |
| VEGF | Vascular endothelial growth factor |
References
- Gougos, A.; Letarte, M. Primary structure of endoglin, an RGD-containing glycoprotein of human endothelial cells. J. Biol. Chem. 1990, 265, 8361–8364. [Google Scholar] [PubMed]
- Cheifetz, S.; Bellon, T.; Cales, C.; Vera, S.; Bernabeu, C.; Massague, J.; Letarte, M. Endoglin Is a Component of the Transforming Growth-Factor-Beta Receptor System in Human Endothelial-Cells. J. Biol. Chem. 1992, 267, 19027–19030. [Google Scholar] [PubMed]
- Bellon, T.; Corbi, A.; Lastres, P.; Cales, C.; Cebrian, M.; Vera, S.; Cheifetz, S.; Massague, J.; Letarte, M.; Bernabeu, C. Identification and expression of two forms of the human transforming growth factor-beta-binding protein endoglin with distinct cytoplasmic regions. Eur. J. Immunol. 1993, 23, 2340–2345. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Ichijo, H.; Grimsby, S.; Moren, A.; ten Dijke, P.; Miyazono, K. Endoglin forms a heteromeric complex with the signaling receptors for transforming growth factor-beta. J. Biol. Chem. 1994, 269, 1995–2001. [Google Scholar] [PubMed]
- McAllister, K.A.; Grogg, K.M.; Johnson, D.W.; Gallione, C.J.; Baldwin, M.A.; Jackson, C.E.; Helmbold, E.A.; Markel, D.S.; McKinnon, W.C.; Murrell, J.; et al. Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat. Genet. 1994, 8, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Goumans, M.J.; Mummery, C. Functional analysis of the TGFbeta receptor/Smad pathway through gene ablation in mice. Int. J. Dev. Biol. 2000, 44, 253–265. [Google Scholar]
- Arthur, H.M.; Ure, J.; Smith, A.J.; Renforth, G.; Wilson, D.I.; Torsney, E.; Charlton, R.; Parums, D.V.; Jowett, T.; Marchuk, D.A.; et al. Endoglin, an ancillary TGFbeta receptor, is required for extraembryonic angiogenesis and plays a key role in heart development. Dev. Biol. 2000, 217, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Ollauri-Ibanez, C.; Lopez-Novoa, J.M.; Pericacho, M. Endoglin-based biological therapy in the treatment of angiogenesis-dependent pathologies. Expert Opin. Biol. 2017, 17, 1053–1063. [Google Scholar] [CrossRef]
- Rosen, L.S.; Gordon, M.S.; Robert, F.; Matei, D.E. Endoglin for targeted cancer treatment. Curr. Oncol. Rep. 2014, 16, 365. [Google Scholar] [CrossRef]
- Paauwe, M.; ten Dijke, P.; Hawinkels, L.J. Endoglin for tumor imaging and targeted cancer therapy. Expert. Opin. Ther. Targets 2013, 17, 421–435. [Google Scholar] [CrossRef]
- O’Connell, P.J.; McKenzie, A.; Fisicaro, N.; Rockman, S.P.; Pearse, M.J.; d’Apice, A.J. Endoglin: A 180-kD endothelial cell and macrophage restricted differentiation molecule. Clin. Exp. Immunol. 1992, 90, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Gougos, A.; St Jacques, S.; Greaves, A.; O’Connell, P.J.; d’Apice, A.J.; Buhring, H.J.; Bernabeu, C.; van Mourik, J.A.; Letarte, M. Identification of distinct epitopes of endoglin, an RGD-containing glycoprotein of endothelial cells, leukemic cells, and syncytiotrophoblasts. Int. Immunol. 1992, 4, 83–92. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, S.; Cymerman, U.; Pece, N.; Letarte, M. Molecular characterization and in situ localization of murine endoglin reveal that it is a transforming growth factor-beta binding protein of endothelial and stromal cells. Endocrinology 1994, 134, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- Schoonderwoerd, M.J.A.; Goumans, M.T.H.; Hawinkels, L. Endoglin: Beyond the Endothelium. Biomolecules 2020, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, C.; Lopez-Novoa, J.M.; Quintanilla, M. The emerging role of TGF-beta superfamily coreceptors in cancer. Biochim. Biophys. Acta 2009, 1792, 954–973. [Google Scholar] [CrossRef]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A Marker of Tumor Vasculature and Potential Target for Therapy. Clin. Cancer Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef]
- Goumans, M.J.; Lebrin, F.; Valdimarsdottir, G. Controlling the angiogenic switch: A balance between two distinct TGF-b receptor signaling pathways. Trends Cardiovasc. Med. 2003, 13, 301–307. [Google Scholar] [CrossRef]
- Hawinkels, L.J.; Kuiper, P.; Wiercinska, E.; Verspaget, H.W.; Liu, Z.; Pardali, E.; Sier, C.F.; ten Dijke, P. Matrix Metalloproteinase-14 (MT1-MMP)-Mediated Endoglin Shedding Inhibits Tumor Angiogenesis. Cancer Res. 2010, 70, 4141–4150. [Google Scholar] [CrossRef] [Green Version]
- Aristorena, M.; Gallardo-Vara, E.; Vicen, M.; de Las Casas-Engel, M.; Ojeda-Fernandez, L.; Nieto, C.; Blanco, F.J.; Valbuena-Diez, A.C.; Botella, L.M.; Nachtigal, P.; et al. MMP-12, Secreted by Pro-Inflammatory Macrophages, Targets Endoglin in Human Macrophages and Endothelial Cells. Int. J. Mol. Sci. 2019, 20, 3107. [Google Scholar] [CrossRef] [Green Version]
- Armaly, Z.; Jadaon, J.E.; Jabbour, A.; Abassi, Z.A. Preeclampsia: Novel Mechanisms and Potential Therapeutic Approaches. Front. Physiol. 2018, 9, 973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicen, M.; Igreja Sa, I.C.; Tripska, K.; Vitverova, B.; Najmanova, I.; Eissazadeh, S.; Micuda, S.; Nachtigal, P. Membrane and soluble endoglin role in cardiovascular and metabolic disorders related to metabolic syndrome. Cell Mol. Life Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lawera, A.; Tong, Z.; Thorikay, M.; Redgrave, R.E.; Cai, J.; van Dinther, M.; Morrell, N.W.; Afink, G.B.; Charnock-Jones, D.S.; Arthur, H.M.; et al. Role of soluble endoglin in BMP9 signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 17800–17808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Bindokas, V.; Shen, J.; Fan, H.; Hoffman, R.M.; Xing, H.R. Time-course imaging of therapeutic functional tumor vascular normalization by antiangiogenic agents. Mol. Cancer 2011, 10, 1173–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, Y.; Hong, H.; Cai, W. Multimodality molecular imaging of CD105 (Endoglin) expression. Int. J. Clin. Exp. Med. 2011, 4, 32–42. [Google Scholar]
- Matsuno, F.; Haruta, Y.; Kondo, M.; Tsai, H.; Barcos, M.; Seon, B.K. Induction of lasting complete regression of preformed distinct solid tumors by targeting the tumor vasculature using two new anti-endoglin monoclonal antibodies. Clin. Cancer Res. 1999, 5, 371–382. [Google Scholar]
- Takahashi, N.; Haba, A.; Matsuno, F.; Seon, B.K. Antiangiogenic therapy of established tumors in human skin/severe combined immunodeficiency mouse chimeras by anti-endoglin (CD105) monoclonal antibodies, and synergy between anti-endoglin antibody and cyclophosphamide. Cancer Res. 2001, 61, 7846–7854. [Google Scholar]
- Shiozaki, K.; Harada, N.; Greco, W.R.; Haba, A.; Uneda, S.; Tsai, H.; Seon, B.K. Antiangiogenic chimeric anti-endoglin (CD105) antibody: Pharmacokinetics and immunogenicity in nonhuman primates and effects of doxorubicin. Cancer Immunol. Immunother. 2006, 55, 140–150. [Google Scholar] [CrossRef]
- Tsujie, M.; Uneda, S.; Tsai, H.; Seon, B.K. Effective anti-angiogenic therapy of established tumors in mice by naked anti-human endoglin (CD105) antibody: Differences in growth rate and therapeutic response between tumors growing at different sites. Int. J. Oncol. 2006, 29, 1087–1094. [Google Scholar] [CrossRef] [Green Version]
- Tsujie, M.; Tsujie, T.; Toi, H.; Uneda, S.; Shiozaki, K.; Tsai, H.; Seon, B.K. Anti-tumor activity of an anti-endoglin monoclonal antibody is enhanced in immunocompetent mice. Int. J. Cancer 2008, 122, 2266–2273. [Google Scholar] [CrossRef] [Green Version]
- Seon, B.K.; Haba, A.; Matsuno, F.; Takahashi, N.; Tsujie, M.; She, X.; Harada, N.; Uneda, S.; Tsujie, T.; Toi, H.; et al. Endoglin-targeted cancer therapy. Curr. Drug Deliv. 2011, 8, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Schoonderwoerd, M.J.A.; Koops, M.F.M.; Angela, R.A.; Koolmoes, B.; Toitou, M.; Paauwe, M.; Barnhoorn, M.C.; Liu, Y.; Sier, C.F.M.; Hardwick, J.C.H.; et al. Targeting Endoglin-Expressing Regulatory T Cells in the Tumor Microenvironment Enhances the Effect of PD1 Checkpoint Inhibitor Immunotherapy. Clin. Cancer Res. 2020, 26, 3831–3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan-Stevaux, O.; Zhong, W.; Culp, S.; Shaffer, K.; Hoover, J.; Wickramasinghe, D.; Ruefli-Brasse, A. Endoglin requirement for BMP9 signaling in endothelial cells reveals new mechanism of action for selective anti-endoglin antibodies. PLoS ONE 2012, 7, e50920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pan, C.C.; Bloodworth, J.C.; Nixon, A.B.; Theuer, C.; Hoyt, D.G.; Lee, N.Y. Antibody-directed coupling of endoglin and MMP-14 is a key mechanism for endoglin shedding and deregulation of TGF-beta signaling. Oncogene 2014, 33, 3970–3979. [Google Scholar] [CrossRef] [PubMed]
- Paauwe, M.; Heijkants, R.C.; Oudt, C.H.; van Pelt, G.W.; Cui, C.; Theuer, C.P.; Hardwick, J.C.; Sier, C.F.; Hawinkels, L.J. Endoglin targeting inhibits tumor angiogenesis and metastatic spread in breast cancer. Oncogene 2016, 35, 4069–4079. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Bokhove, M.; Croci, R.; Zamora-Caballero, S.; Han, L.; Letarte, M.; de Sanctis, D.; Jovine, L. Structural Basis of the Human Endoglin-BMP9 Interaction: Insights into BMP Signaling and HHT1. Cell Rep. 2017, 19, 1917–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergers, G.; Hanahan, D. Modes of resistance to anti-angiogenic therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.K. Antiangiogenesis strategies revisited: From starving tumors to alleviating hypoxia. Cancer Cell 2014, 26, 605–622. [Google Scholar] [CrossRef] [Green Version]
- Kieran, M.W.; Kalluri, R.; Cho, Y.J. The VEGF pathway in cancer and disease: Responses, resistance, and the path forward. Cold Spring Harb. Perspect. Med. 2012, 2, a006593. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tian, H.; Blobe, G.C.; Theuer, C.P.; Hurwitz, H.I.; Nixon, A.B. Effects of the combination of TRC105 and bevacizumab on endothelial cell biology. Invest. New Drugs 2014, 32, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Bockhorn, M.; Tsuzuki, Y.; Xu, L.; Frilling, A.; Broelsch, C.E.; Fukumura, D. Differential vascular and transcriptional responses to anti-vascular endothelial growth factor antibody in orthotopic human pancreatic cancer xenografts. Clin. Cancer Res. 2003, 9, 4221–4226. [Google Scholar]
- Liu, Z.; Lebrin, F.; Maring, J.A.; van den Driesche, S.; van der Brink, S.; van, D.M.; Thorikay, M.; Martin, S.; Kobayashi, K.; Hawinkels, L.J.; et al. ENDOGLIN is dispensable for vasculogenesis, but required for vascular endothelial growth factor-induced angiogenesis. PLoS ONE 2014, 9, e86273. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Huang, J.J.; Golzio, C.; Gao, X.; Hector-Greene, M.; Katsanis, N.; Blobe, G.C. Endoglin interacts with VEGFR2 to promote angiogenesis. FASEB J. 2018, 32, 2934–2949. [Google Scholar] [CrossRef] [Green Version]
- Brossa, A.; Buono, L.; Bussolati, B. Effect of the monoclonal antibody TRC105 in combination with Sunitinib on renal tumor derived endothelial cells. Oncotarget 2018, 9, 22680–22692. [Google Scholar] [CrossRef] [Green Version]
- Uneda, S.; Toi, H.; Tsujie, T.; Tsujie, M.; Harada, N.; Tsai, H.; Seon, B.K. Anti-endoglin monoclonal antibodies are effective for suppressing metastasis and the primary tumors by targeting tumor vasculature. Int. J. Cancer 2009, 125, 1446–1453. [Google Scholar] [CrossRef] [Green Version]
- Dourado, K.M.C.; Baik, J.; Oliveira, V.K.P.; Beltrame, M.; Yamamoto, A.; Theuer, C.P.; Figueiredo, C.A.V.; Verneris, M.R.; Perlingeiro, R.C.R. Endoglin: A novel target for therapeutic intervention in acute leukemias revealed in xenograft mouse models. Blood 2017, 129, 2526–2536. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Zhu, W.; Coffman, L.; Vlad, A.; Schwartz, L.E.; Elishaev, E.; Drapkin, R.; Buckanovich, R.J. CD105 Is Expressed in Ovarian Cancer Precursor Lesions and Is Required for Metastasis to the Ovary. Cancers 2019, 11, 1710. [Google Scholar] [CrossRef] [Green Version]
- Madhav, A.; Andres, A.; Duong, F.; Mishra, R.; Haldar, S.; Liu, Z.; Angara, B.; Gottlieb, R.; Zumsteg, Z.S.; Bhowmick, N.A. Antagonizing CD105 enhances radiation sensitivity in prostate cancer. Oncogene 2018, 37, 4385–4397. [Google Scholar] [CrossRef]
- Kato, M.; Placencio-Hickok, V.R.; Madhav, A.; Haldar, S.; Tripathi, M.; Billet, S.; Mishra, R.; Smith, B.; Rohena-Rivera, K.; Agarwal, P.; et al. Heterogeneous cancer-associated fibroblast population potentiates neuroendocrine differentiation and castrate resistance in a CD105-dependent manner. Oncogene 2019, 38, 716–730. [Google Scholar] [CrossRef]
- Paauwe, M.; Schoonderwoerd, M.J.A.; Helderman, R.; Harryvan, T.J.; Groenewoud, A.; van Pelt, G.W.; Bor, R.; Hemmer, D.M.; Versteeg, H.H.; Snaar-Jagalska, B.E.; et al. Endoglin Expression on Cancer-Associated Fibroblasts Regulates Invasion and Stimulates Colorectal Cancer Metastasis. Clin. Cancer Res. 2018, 24, 6331–6344. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.W.; Sheard, M.A.; Malvar, J.; Fernandez, G.E.; DeClerck, Y.A.; Blavier, L.; Shimada, H.; Theuer, C.P.; Sposto, R.; Seeger, R.C. Anti-CD105 Antibody Eliminates Tumor Microenvironment Cells and Enhances Anti-GD2 Antibody Immunotherapy of Neuroblastoma with Activated Natural Killer Cells. Clin. Cancer Res. 2019, 25, 4761–4774. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lyu, J.; Bell Burdett, K.; Sibley, A.B.; Hatch, A.J.; Starr, M.D.; Brady, J.C.; Hammond, K.; Marmorino, F.; Rossini, D.; et al. Prognostic and Predictive Biomarkers in Patients with Metastatic Colorectal Cancer Receiving Regorafenib. Mol. Cancer 2020, 19, 2146–2154. [Google Scholar] [CrossRef]
- Rosen, L.S.; Hurwitz, H.I.; Wong, M.K.; Goldman, J.; Mendelson, D.S.; Figg, W.D.; Spencer, S.; Adams, B.J.; Alvarez, D.; Seon, B.K.; et al. A phase I first-in-human study of TRC105 (Anti-Endoglin Antibody) in patients with advanced cancer. Clin. Cancer Res. 2012, 18, 4820–4829. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Starr, M.D.; Brady, J.C.; Dellinger, A.; Pang, H.; Adams, B.; Theuer, C.P.; Lee, N.Y.; Hurwitz, H.I.; Nixon, A.B. Modulation of circulating protein biomarkers following TRC105 (anti-endoglin antibody) treatment in patients with advanced cancer. Cancer Med. 2014, 3, 580–591. [Google Scholar] [CrossRef]
- Gordon, M.S.; Robert, F.; Matei, D.; Mendelson, D.S.; Goldman, J.W.; Chiorean, E.G.; Strother, R.M.; Seon, B.K.; Figg, W.D.; Peer, C.J.; et al. An open-label phase Ib dose-escalation study of TRC105 (anti-endoglin antibody) with bevacizumab in patients with advanced cancer. Clin. Cancer Res. 2014, 20, 5918–5926. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Charnsangavej, C.; Faria, S.C.; Macapinlac, H.A.; Burgess, M.A.; Patel, S.R.; Chen, L.L.; Podoloff, D.A.; Benjamin, R.S. Correlation of computed tomography and positron emission tomography in patients with metastatic gastrointestinal stromal tumor treated at a single institution with imatinib mesylate: Proposal of new computed tomography response criteria. J. Clin. Oncol 2007, 25, 1753–1759. [Google Scholar] [CrossRef]
- Liu, Y.; Starr, M.D.; Brady, J.C.; Rushing, C.; Pang, H.; Adams, B.; Alvarez, D.; Theuer, C.P.; Hurwitz, H.I.; Nixon, A.B. Modulation of Circulating Protein Biomarkers in Cancer Patients Receiving Bevacizumab and the Anti-Endoglin Antibody, TRC105. Mol. Cancer 2018, 17, 2248–2256. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Starr, M.D.; Bulusu, A.; Pang, H.; Wong, N.S.; Honeycutt, W.; Amara, A.; Hurwitz, H.I.; Nixon, A.B. Correlation of angiogenic biomarker signatures with clinical outcomes in metastatic colorectal cancer patients receiving capecitabine, oxaliplatin, and bevacizumab. Cancer Med. 2013, 2, 234–242. [Google Scholar] [CrossRef]
- Maxwell, M.; Naber, S.P.; Wolfe, H.J.; Hedley-Whyte, E.T.; Galanopoulos, T.; Neville-Golden, J.; Antoniades, H.N. Expression of angiogenic growth factor genes in primary human astrocytomas may contribute to their growth and progression. Cancer Res. 1991, 51, 1345–1351. [Google Scholar]
- Shen, C.; Kaelin, W.G., Jr. The VHL/HIF axis in clear cell renal carcinoma. Semin. Cancer Biol. 2013, 23, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Morse, M.A.; Sun, W.; Kim, R.; He, A.R.; Abada, P.B.; Mynderse, M.; Finn, R.S. The Role of Angiogenesis in Hepatocellular Carcinoma. Clin. Cancer Res. 2019, 25, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Chen, Y.S.; Yao, Y.D.; Chen, J.Q.; Chen, J.N.; Huang, S.Y.; Zeng, Y.J.; Yao, H.R.; Zeng, S.H.; Fu, Y.S.; et al. CCL18 from tumor-associated macrophages promotes angiogenesis in breast cancer. Oncotarget 2015, 6, 34758–34773. [Google Scholar] [CrossRef] [Green Version]
- Sandlund, J.; Hedberg, Y.; Bergh, A.; Grankvist, K.; Ljungberg, B.; Rasmuson, T. Endoglin (CD105) expression in human renal cell carcinoma. BJU Int. 2006, 97, 706–710. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Tolar, J.; Nauta, A.J.; Osborn, M.J.; Panoskaltsis Mortari, A.; McElmurry, R.T.; Bell, S.; Xia, L.; Zhou, N.; Riddle, M.; Schroeder, T.M.; et al. Sarcoma derived from cultured mesenchymal stem cells. Stem. Cells 2007, 25, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, M. A phase 2 trial of TRC105 with bevacizumab for bevacizumab refractory glioblastoma. J. Clin. Oncol 2016, 34. [Google Scholar] [CrossRef]
- Ravi, V. TAPPAS: An adaptive enrichment phase 3 trial of TRC105 and pazopanib versus pazopanib alone in patients with advanced angiosarcoma. J. Clin. Oncol. 2018, 36. [Google Scholar] [CrossRef]
- Attia, S. A phase 1B/phase 2A study of TRC105 (Endoglin Antibody) in combination with pazopanib (P) in patients (pts) with advanced soft tissue sarcoma (STS). J. Clin. Oncol. 2016, 34. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Michaelson, M.D.; Posadas, E.M.; Sonpavde, G.P.; McDermott, D.F.; Nixon, A.B.; Liu, Y.; Yuan, Z.; Seon, B.K.; Walsh, M.; et al. An Open Label Phase Ib Dose Escalation Study of TRC105 (Anti-Endoglin Antibody) with Axitinib in Patients with Metastatic Renal Cell Carcinoma. Oncologist 2019, 24, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Dorff, T.B.; Longmate, J.A.; Pal, S.K.; Stadler, W.M.; Fishman, M.N.; Vaishampayan, U.N.; Rao, A.; Pinksi, J.K.; Hu, J.S.; Quinn, D.I.; et al. Bevacizumab alone or in combination with TRC105 for patients with refractory metastatic renal cell cancer. Cancer 2017, 123, 4566–4573. [Google Scholar] [CrossRef]
- Levine, E. A phase Ib dose-escalation study of TRC105 (anti-endoglin antibody) in combination with capecitabine for advanced solid tumors (including patients with progressive or recurrent HER2-negative metastatic breast cancer). J. Clin. Oncol 2013, 31. [Google Scholar] [CrossRef]
- Karzai, F.H.; Apolo, A.B.; Cao, L.; Madan, R.A.; Adelberg, D.E.; Parnes, H.; McLeod, D.G.; Harold, N.; Peer, C.; Yu, Y.; et al. A phase I study of TRC105 anti-endoglin (CD105) antibody in metastatic castration-resistant prostate cancer. BJU Int. 2015, 116, 546–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, A.G.; Ma, C.; Ulahannan, S.V.; Rahma, O.E.; Makarova-Rusher, O.; Cao, L.; Yu, Y.; Kleiner, D.E.; Trepel, J.; Lee, M.J.; et al. Phase I and Preliminary Phase II Study of TRC105 in Combination with Sorafenib in Hepatocellular Carcinoma. Clin. Cancer Res. 2017, 23, 4633–4641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, A.G.; Ulahannan, S.V.; Cao, L.; Rahma, O.E.; Makarova-Rusher, O.V.; Kleiner, D.E.; Fioravanti, S.; Walker, M.; Carey, S.; Yu, Y.; et al. A phase II study of TRC105 in patients with hepatocellular carcinoma who have progressed on sorafenib. United Eur. Gastroenterol. J. 2015, 3, 453–461. [Google Scholar] [CrossRef]
- Apolo, A.B.; Karzai, F.H.; Trepel, J.B.; Alarcon, S.; Lee, S.; Lee, M.J.; Tomita, Y.; Cao, L.; Yu, Y.; Merino, M.J.; et al. A Phase II Clinical Trial of TRC105 (Anti-Endoglin Antibody) in Adults With Advanced/Metastatic Urothelial Carcinoma. Clin. Genitourin. Cancer 2017, 15, 77–85. [Google Scholar] [CrossRef]
- Chang, X.; Yao, J.; He, Q.; Liu, M.; Duan, T.; Wang, K. Exosomes From Women With Preeclampsia Induced Vascular Dysfunction by Delivering sFlt (Soluble Fms-Like Tyrosine Kinase)-1 and sEng (Soluble Endoglin) to Endothelial Cells. Hypertension 2018, 72, 1381–1390. [Google Scholar] [CrossRef]
- Ruiz-Remolina, L.; Ollauri-Ibanez, C.; Perez-Roque, L.; Nunez-Gomez, E.; Perez-Barriocanal, F.; Lopez-Novoa, J.M.; Pericacho, M.; Rodriguez-Barbero, A. Circulating soluble endoglin modifies the inflammatory response in mice. PLoS ONE 2017, 12, e0188204. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Starr, M.D.; Brady, J.C.; Rushing, C.; Bulusu, A.; Pang, H.; Honeycutt, W.; Amara, A.; Altomare, I.; Uronis, H.E.; et al. Biomarker signatures correlate with clinical outcome in refractory metastatic colorectal cancer patients receiving bevacizumab and everolimus. Mol. Cancer 2015, 14, 1048–1056. [Google Scholar] [CrossRef] [Green Version]
- Murukesh, N.; Dive, C.; Jayson, G.C. Biomarkers of angiogenesis and their role in the development of VEGF inhibitors. Br. J. Cancer 2010, 102, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Choueiri, T.K.; Kocsis, J.; Pachynski, R.K.; Poprach, A.; deShazo, M.; Zakharia, Y.; Lara, P.N.; Pal, S.K.; Geczi, L.; Ho, T.H.; et al. Results of the Phase 2 TRAXAR Study: A Randomized Phase 2 Trial of Axitinib and TRC105 (TRAX) versus AXitinib (AX) Alone in Patients with Advanced or Metastatic Renal Cell Carcinoma (mRCC). Ann. Oncol. 2019, 30 (Suppl. 5), v356–v402. [Google Scholar] [CrossRef]
- Mehta, C.R.; Liu, L.; Theuer, C. An adaptive population enrichment phase III trial of TRC105 and pazopanib versus pazopanib alone in patients with advanced angiosarcoma (TAPPAS trial). Ann. Oncol. 2019, 30, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Lenato, G.M.; Guanti, G. Hereditary Haemorrhagic Telangiectasia (HHT): Genetic and molecular aspects. Curr. Pharm. Des. 2006, 12, 1173–1193. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Mehta, J.L.; Li, D.; Joseph, L.; Joseph, J. Transforming growth factor beta receptor endoglin is expressed in cardiac fibroblasts and modulates profibrogenic actions of angiotensin II. Circ. Res. 2004, 95, 1167–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapur, N.K.; Wilson, S.; Yunis, A.A.; Qiao, X.; Mackey, E.; Paruchuri, V.; Baker, C.; Aronovitz, M.J.; Karumanchi, S.A.; Letarte, M.; et al. Reduced endoglin activity limits cardiac fibrosis and improves survival in heart failure. Circulation 2012, 125, 2728–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawinkels, L.J.; Garcia de Vinuesa, A.; ten Dijke, P. Activin receptor-like kinase 1 as a target for anti-angiogenesis therapy. Expert. Opin. Investig. Drugs 2013, 22, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- Cunha, S.I.; Pietras, K. ALK1 as an emerging target for antiangiogenic therapy of cancer. Blood 2011, 117, 6999–7006. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Trial Description | NCT Number | Trial Phase | Patients Enrolled | Primary Outcome |
|---|---|---|---|---|
| Open label phase 1 dose-finding study of TRC105 in patients with solid cancer | NCT00582985 | 1 | 51 | Safety |
| Preoperative combination of letrozole, everolimus and TRC105 in postmenopausal hormone receptor-positive and Her2-negative breast cancer | NCT02520063 | 1/2 | 14 | MTD |
| Open label dose-finding study of TRC105 plus capecitabine for metastatic breast cancer (TRC105) | NCT01326481 | 1/2 | 19 | MTD |
| A phase I/II study of TRC105 in metastatic castration-resistant prostate cancer (CRPC) | NCT01090765 | 1/2 | 21 | MTD |
| Sorafenib and TRC105 in hepatocellular cancer | NCT01306058 | 1/2 | 27 | MTD and TTP |
| Open label continuation study of TRC105 for patients who have completed a prior TRC105 trial | NCT02354612 | 1/2 | 50 | Long-term TRC105 response |
| Bevacizumab with or without anti-endoglin monoclonal antibody TRC105 in treating patients with recurrent glioblastoma multiforme | NCT01648348 | 1/2 | 116 | MTD, safety and PFS |
| Study of carotuximab (TRC105) plus nivolumab in patients with metastatic NSCLC | NCT03181308 | 1b | 11 | Safety |
| Study of TRC105 combined with standard-dose bevacizumab for advanced solid tumors for which bevacizumab is indicated | NCT01332721 | 1b | 38 | MTD |
| A study of TRC105 in combination with paclitaxel/carboplatin and bevacizumab in non-squamous cell lung cancer | NCT02429843 | 1b | 16 | Change in medical management |
| A phase 1B dose-escalation and phase 2a study of carotuximab (TRC105) in combination with pazopanib in patients with advanced soft tissue sarcoma | NCT01975519 | 1b, 2 | 30, 89 | Safety, PFS, ORR |
| Trial of TRC105 and sorafenib in patients with hepatocellular carcinoma (HCC) | NCT02560779 | 1b/2 | 27 | Safety and ORR |
| TRC105 for recurrent glioblastoma | NCT01778530 | 2 | 2 | Radiographic response rate |
| TRC105 combined with standard-dose bevacizumab for two patients with metastatic and refractory choriocarcinoma | NCT02396511 | 2 | 2 | PFS and ORR |
| Study of TRC105 and bevacizumab in patients with refractory gestational trophoblastic neoplasia (GTN) | NCT02664961 | 2 | 3 | ORR |
| TRC105 for liver cancer that has not responded to sorafenib | NCT01375569 | 2 | 11 | TTP |
| Study of TRC105 with abiraterone and with enzalutamide in prostate cancer patients progressing on therapy | NCT03418324 | 2 | 11 | Disease stabilization or improvement at 2 months |
| TRC105 in adults with advanced/metastatic urothelial carcinoma | NCT01328574 | 2 | 13 | PFS |
| Study of TRC105 + paclitaxel/carboplatin and bevacizumab in patients with NSCLC | NCT03780010 | 2 | 15 | Safety |
| A phase 2 evaluation of TRC105 in combination with bevacizumab in patients with glioblastoma (105GM201) | NCT01564914 | 2 | 22 | OS |
| Evaluation of TRC105 in the treatment of recurrent ovarian, fallopian tube or primary peritoneal carcinoma | NCT01381861 | 2 | 23 | PFS, ORR, safety |
| Bevacizumab with or without TRC105 in treating patients with metastatic kidney cancer | NCT01727089 | 2 | 59 | PFS |
| Randomized phase 2 trial of axitinib and TRC105 versus axitinib alone in patients renal cell carcinoma | NCT01806064 | 2 | 150 | Safety and PFS |
| Trial of TRC105 and pazopanib versus pazopanib alone in patients with advanced angiosarcoma (TAPPAS) | NCT02979899 | 3 | 128 | PFS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Paauwe, M.; Nixon, A.B.; Hawinkels, L.J.A.C. Endoglin Targeting: Lessons Learned and Questions That Remain. Int. J. Mol. Sci. 2021, 22, 147. https://doi.org/10.3390/ijms22010147
Liu Y, Paauwe M, Nixon AB, Hawinkels LJAC. Endoglin Targeting: Lessons Learned and Questions That Remain. International Journal of Molecular Sciences. 2021; 22(1):147. https://doi.org/10.3390/ijms22010147
Chicago/Turabian StyleLiu, Yingmiao, Madelon Paauwe, Andrew B. Nixon, and Lukas J.A.C. Hawinkels. 2021. "Endoglin Targeting: Lessons Learned and Questions That Remain" International Journal of Molecular Sciences 22, no. 1: 147. https://doi.org/10.3390/ijms22010147
APA StyleLiu, Y., Paauwe, M., Nixon, A. B., & Hawinkels, L. J. A. C. (2021). Endoglin Targeting: Lessons Learned and Questions That Remain. International Journal of Molecular Sciences, 22(1), 147. https://doi.org/10.3390/ijms22010147