Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
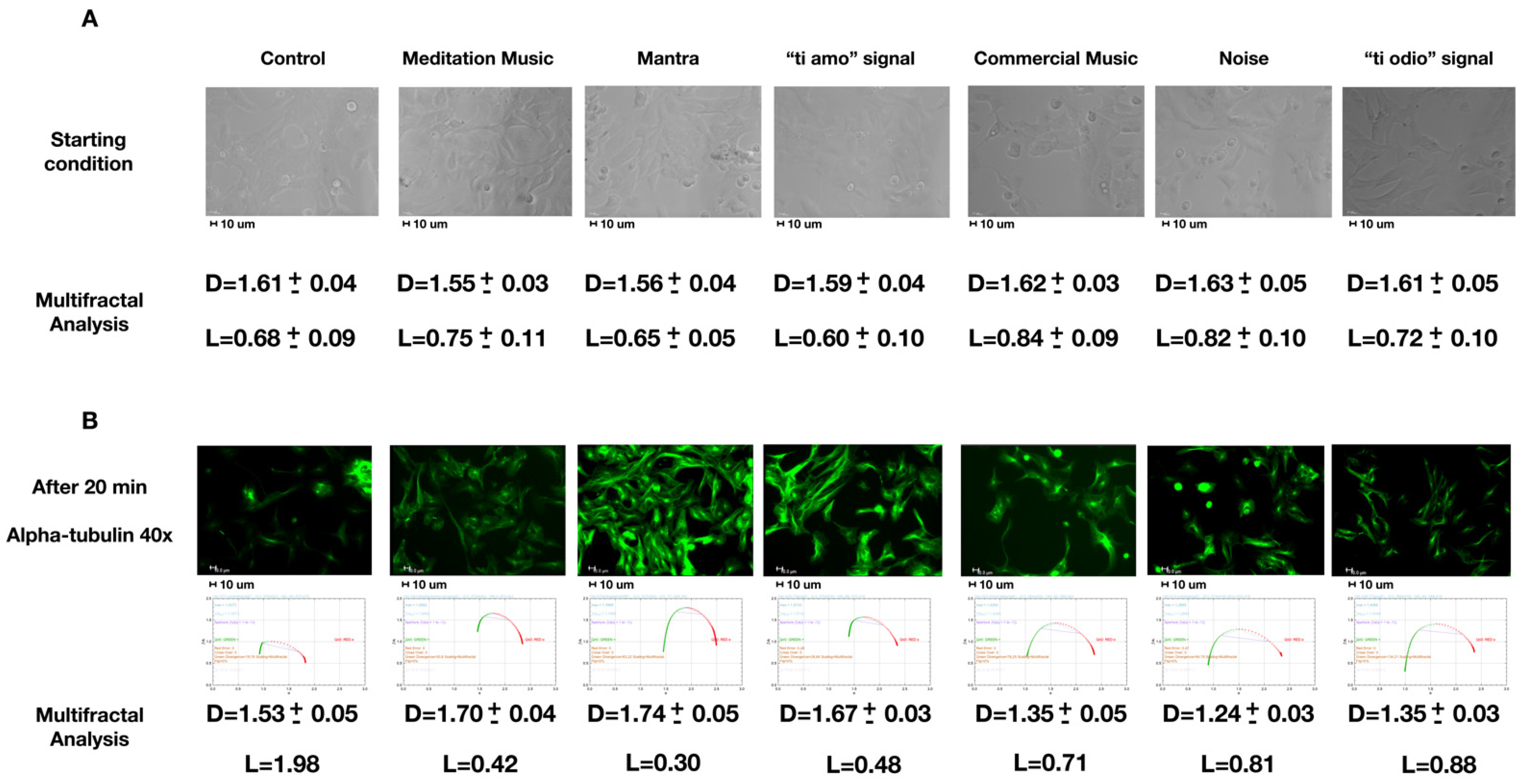
2.1. Spatial Organization: Starting Condition and Alpha-Tubulin Staining after Different Acoustic Stimulations
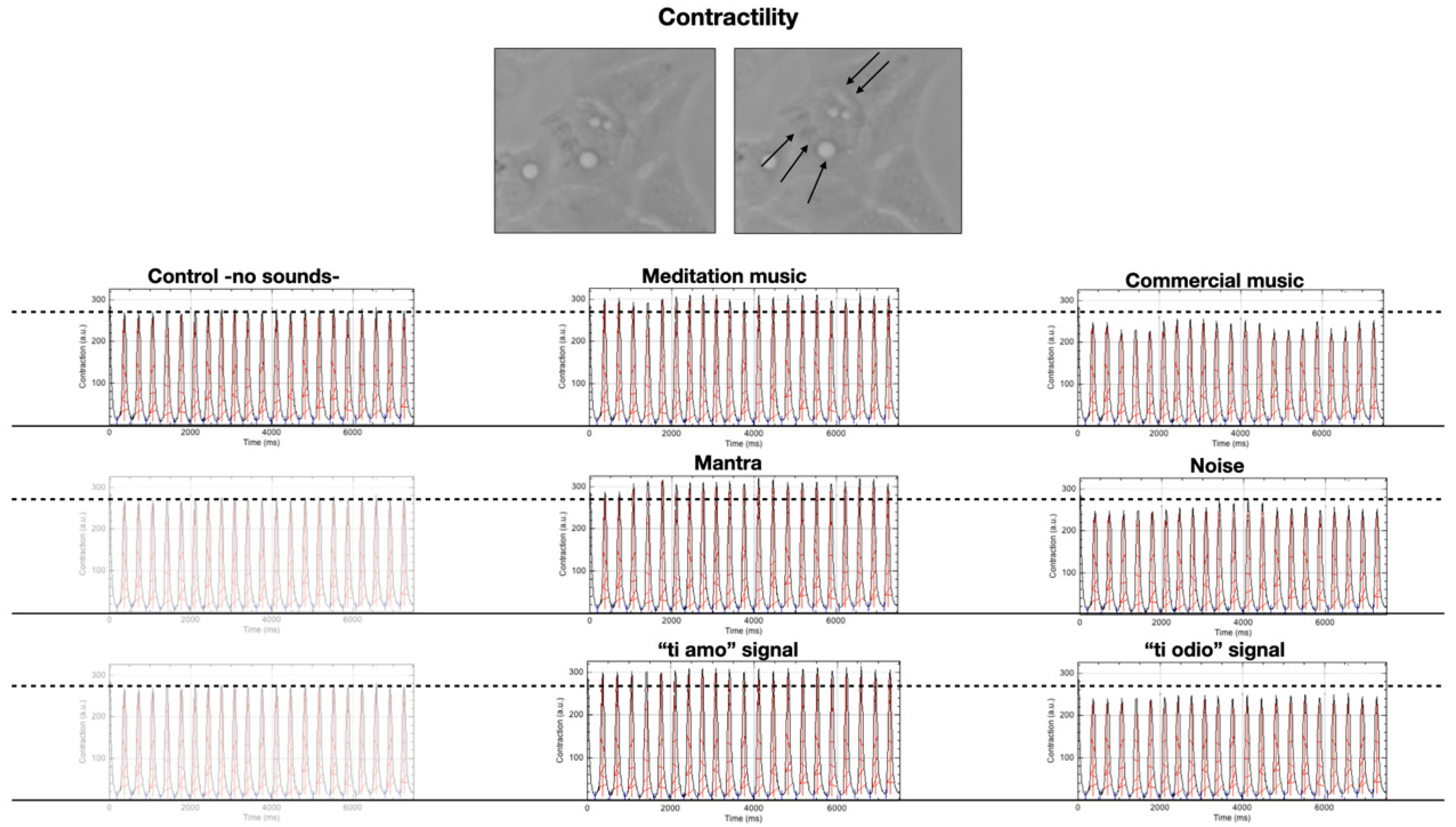
2.2. Contractility
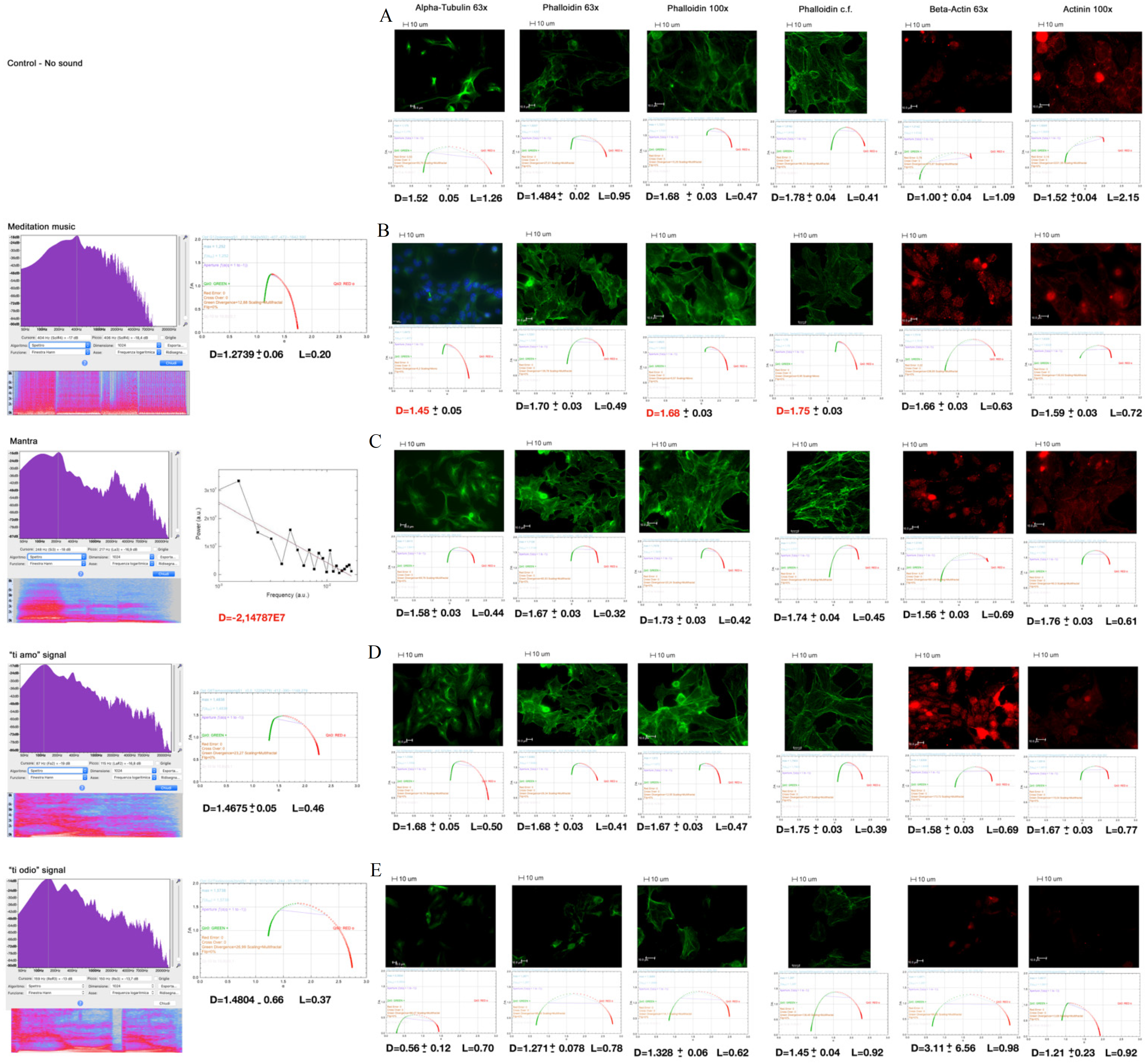
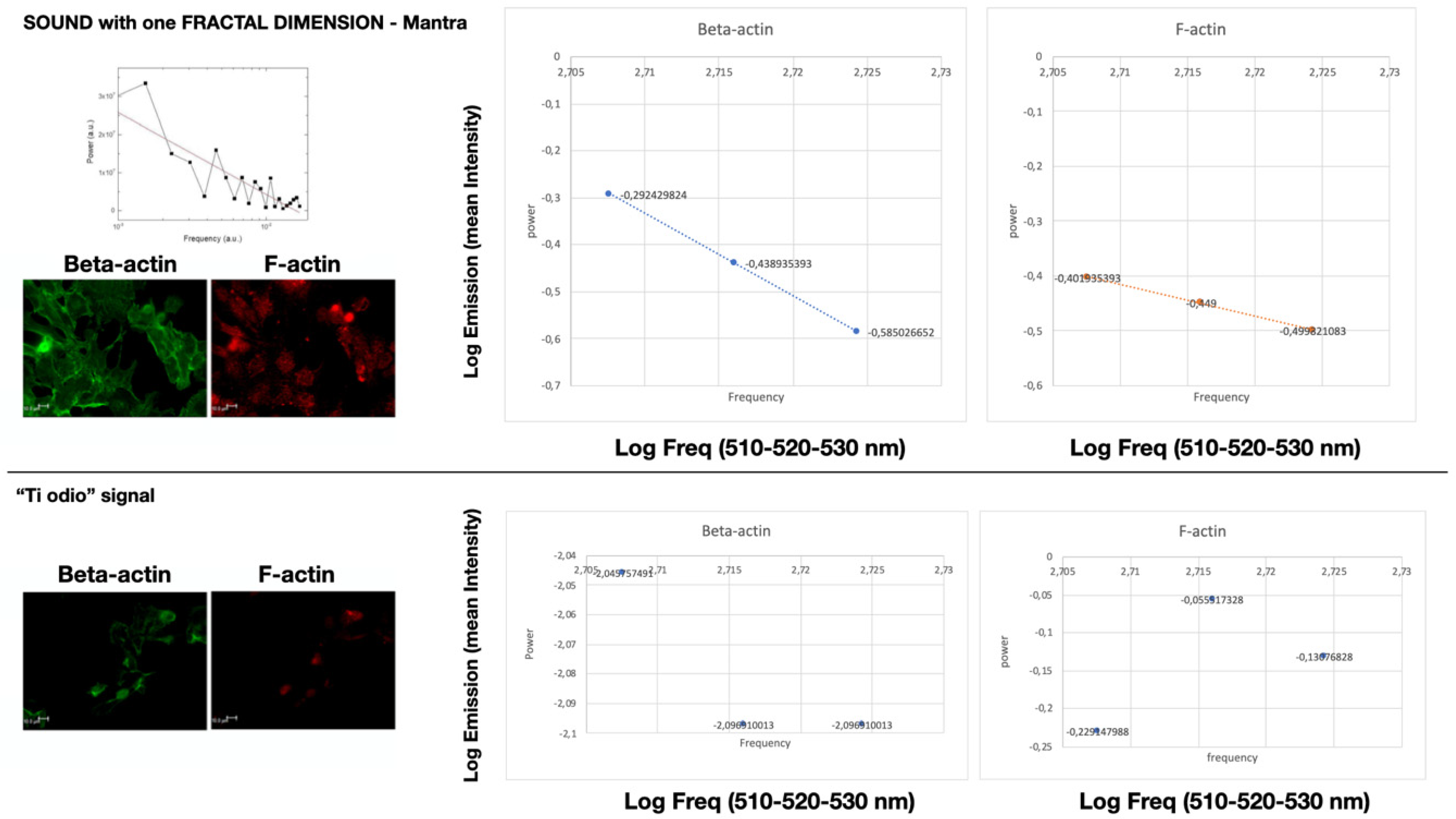
2.3. Specific Sound Analysis, Immunofluorescence, and Confocal Microscopy
2.4. Mean Fluorescence Intensity
3. Discussion
3.1. Theoretical Modelling
Modelling the Dynamical Origin of Positive and Negative Effects Due to Sounds
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RR | Relaxation Response |
| D | Fractal Dimension |
| L | Lacunarity |
Appendix A. Phonons and Spontaneous Breakdown of Symmetry in Quantum Field Theory
Appendix B. Non-Dissipative Energy Transfer on Molecular Chains, Microtubules, and Fractal Structures
References
- Davis, M.T.; Holmes, S.E.; Pietrzak, R.H.; Esterlis, I. Neurobiology of Chronic Stress-Related Psychiatric Disorders: Evidence from Molecular Imaging Studies. Chronic Stress 2017, 1, 247054701771091. [Google Scholar] [CrossRef] [PubMed]
- Dal Lin, C.; Tona, F.; Osto, E. The Heart as a Psychoneuroendocrine and Immunoregulatory Organ. Adv. Exp. Med. Biol. 2018, 1065, 225–239. [Google Scholar] [CrossRef] [PubMed]
- De Hert, M.; Detraux, J.; Vancampfort, D. The intriguing relationship between coronary heart disease and mental disorders. Dialogues Clin. Neurosci. 2018, 20, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Harvard Heart Letter. From Irritated to Enraged: Anger’s Toxic Effect on the Heart. Available online: https://www.health.harvard.edu/heart-health/from-irritated-to-enraged-angers-toxic-effect-on-the-heart (accessed on 1 December 2020).
- Posternak, M.A.; Zimmerman, M. Anger and Aggression in Psychiatric Outpatients. J. Clin. Psychiatry 2002, 63, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Whooley, M.A.; Wong, J. Hostility and Cardiovascular Disease. J. Am. Coll. Cardiol. 2011, 58, 1229–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- AHA Loving-Kindness Meditation for Compassion and Wellbeing. Available online: https://www.heart.org/en/healthy-living/healthy-lifestyle/mental-health-and-wellbeing/loving-kindness-meditation-for-compassion-and-wellbeing (accessed on 1 December 2020).
- Momennasab, M.; Moattari, M.; Abbaszade, A.; Shamshiri, B. Spirituality in survivors of myocardial infarction. Iran. J. Nurs. Midwifery Res. 2012, 17, 343–351. [Google Scholar]
- Dal Lin, C.; Poretto, A.; Scodro, M.; Perazzolo Marra, M.; Iliceto, S.; Tona, F. Coronary microvascular and endothelial function regulation: Crossroads of psychoneuroendocrine immunitary signals and quantum physics. J. Integr. Cardiol. 2015, 1, 132–209. [Google Scholar] [CrossRef] [Green Version]
- McGee, M. Meditation and psychiatry. Psychiatry 2008, 5, 28–41. [Google Scholar]
- Dal Lin, C.; Falanga, M.; De Lauro, E.; De Martino, S.; Vitiello, G. Biochemical and biophysical mechanisms underlying the heart and the brain dialog. AIMS Biophys. 2021, 8, 1–33. [Google Scholar] [CrossRef]
- Dal Lin, C.; Marinova, M.; Rubino, G.; Gola, E.; Brocca, A.; Pantano, G.; Brugnolo, L.; Sarais, C.; Cucchini, U.; Volpe, B.; et al. Thoughts modulate the expression of inflammatory genes and may improve the coronary blood flow in patients after a myocardial infarction. J. Tradit. Complement. Med. 2018, 8, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Dal Lin, C.; Gola, E.; Brocca, A.; Rubino, G.; Marinova, M.; Brugnolo, L.; Plebani, M.; Iliceto, S.; Tona, F. miRNAs may change rapidly with thoughts: The Relaxation Response after myocardial infarction. Eur. J. Integr. Med. 2018, 20, 63–72. [Google Scholar] [CrossRef]
- Pavanello, S.; Campisi, M.; Tona, F.; Dal Lin, C.; Iliceto, S. Exploring Epigenetic Age in Response to Intensive Relaxing Training: A Pilot Study to Slow Down Biological Age. Int. J. Environ. Res. Public Health 2019, 16, 3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Lin, C.; Tona, F.; Osto, E. The crosstalk between the cardiovascular and the immune system. Vasc. Biol. 2019, 1, H83–H88. [Google Scholar] [CrossRef] [Green Version]
- Dal Lin, C.; Tona, F.; Osto, E. Coronary Microvascular Function and Beyond: The Crosstalk between Hormones, Cytokines, and Neurotransmitters. Int. J. Endocrinol. 2015, 2015, 312848. [Google Scholar] [CrossRef]
- Dal Lin, C.; Marinova, M.; Brugnolo, L.; Rubino, G.; Plebani, M.; Tona, F. Rapid Senectome and Alternative Splicing miRNAs Changes With the Relaxation Response: A One Year Follow-Up Study. Preprints 2020. [Google Scholar] [CrossRef]
- Dal Lin, C.; Grasso, R.; Scordino, A.; Triglia, A.; Tona, F.; Iliceto, S.; Vitiello, G.; Elia, V.; Napoli, E.; Germano, R.; et al. Ph, Electric Conductivity and Delayed Luminescence Changes in Human Sera of Subjects Undergoing the Relaxation Response: A Pilot Study. Available online: https://www.preprints.org/manuscript/202004.0202/v1 (accessed on 16 November 2020).
- Dal Lin, C.; Brugnolo, L.; Marinova, M.; Plebani, M.; Iliceto, S.; Tona, F.; Vitiello, G. Toward a Unified View of Cognitive and Biochemical Activity: Meditation and Linguistic Self-Reconstructing May Lead to Inflammation and Oxidative Stress Improvement. Entropy 2020, 22, 818. [Google Scholar] [CrossRef]
- Freeman, W.J. Mass Action in the Nervous System; Academic Press: New York, NY, USA, 1975. [Google Scholar] [CrossRef]
- Freeman, W. Neurodynamics: An Exploration of Mesoscopic Brain Dynamics; Springer: Berlin, Germany, 2000. [Google Scholar] [CrossRef]
- Huth, A.G.; De Heer, W.A.; Griffiths, T.L.; Theunissen, F.E.; Gallant, J.L. Natural speech reveals the semantic maps that tile human cerebral cortex. Nature 2016, 532, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Harrison, N.A.; Gray, M.A.; Gianaros, P.J.; Critchley, H.D. The embodiment of emotional feelings in the brain. J. Neurosci. 2010, 30, 12878–12884. [Google Scholar] [CrossRef] [Green Version]
- Levine, G.N.; Lange, R.A.; Bairey-Merz, C.N.; Davidson, R.J.; Jamerson, K.; Mehta, P.K.; Michos, E.D.; Norris, K.; Ray, I.B.; Saban, K.L.; et al. Meditation and Cardiovascular Risk Reduction. J. Am. Heart Assoc. 2017, 6, e002218. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Karperien, A. FracLac for ImageJ. Available online: http://rsb.info.nih.gov/ij/plugins/fraclac/FLHelp/Introduction.htm (accessed on 1 December 2020).
- Sala, L.; van Meer, B.J.; Tertoolen, L.G.J.; Bakkers, J.; Bellin, M.; Davis, R.P.; Denning, C.; Dieben, M.A.E.; Eschenhagen, T.; Giacomelli, E.; et al. MUSCLEMOTION. Circ. Res. 2018, 122, e5–e16. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, R. Acustica. Fondamenti e Applicazioni, 1st ed.; ISBN 10 8860084466.
- Maragos, P.; Potamianos, A. Fractal dimensions of speech sounds: Computation and application to automatic speech recognition. J. Acoust. Soc. Am. 1999, 105, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Bigerelle, M.; Iost, A. Fractal dimension and classification of music. Chaos Solitons Fractals 2000, 11, 2179–2192. [Google Scholar] [CrossRef]
- Arbesman, S. Fractal Musical Rhythms. Available online: https://www.wired.com/2012/02/fractal-musical-rhythms/ (accessed on 1 December 2020).
- Peitgen, H.-O.; Jürgens, H.; Saupe, D. Chaos and Fractals; Springer: New York, NY, USA, 2004; ISBN 978-1-4684-9396-2. [Google Scholar]
- Su, Z.-Y.; Wu, T. Multifractal analyses of music sequences. Phys. D Nonlinear Phenom. 2006, 221, 188–194. [Google Scholar] [CrossRef]
- Xie, Y.; Shen, C.; Wang, W.; Li, J.; Suo, D.; Popa, B.-I.; Jing, Y.; Cummer, S.A. Acoustic Holographic Rendering with Two-Dimensional Metamaterial-Based Passive Phased Array. Sci. Rep. 2016, 6, 35437. [Google Scholar] [CrossRef]
- Albrecht-Buehler, G. Rudimentary form of cellular “vision”. Proc. Nadl. Acad. Sci. USA 1992, 89, 8288–8292. [Google Scholar] [CrossRef] [Green Version]
- Albrecht-Buehler, G. A long-range attraction between aggregating 3T3 cells mediated by near-infrared light scattering. Proc. Natl. Acad. Sci. USA 2005, 102, 5050–5055. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.; Ghosh, S.; Fujita, D.; Bandyopadhyay, A. Live visualizations of single isolated tubulin protein self-assembly via tunneling current: Effect of electromagnetic pumping during spontaneous growth of microtubule. Sci. Rep. 2014, 4, 7303. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.; Ghosh, S.; Hirata, K.; Fujita, D.; Bandyopadhyay, A. Multi-level memory-switching properties of a single brain microtubule. Appl. Phys. Lett. 2013, 102, 123701. [Google Scholar] [CrossRef]
- Havelka, D.; Cifra, M.; Kučera, O.; Pokorný, J.; Vrba, J. High-frequency electric field and radiation characteristics of cellular microtubule network. J. Theor. Biol. 2011, 286, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Pelling, A.E.; Sehati, S.; Gralla, E.B.; Valentine, J.S.; Gimzewski, J.K. Local nanomechanical motion of the cell wall of Saccharomyces cerevisiae. Science 2004, 305, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Haase, K.; Pelling, A.E. Investigating cell mechanics with atomic force microscopy. J. R. Soc. Interface 2015, 12, 20140970. [Google Scholar] [CrossRef] [PubMed]
- Uzer, G.; Thompson, W.R.; Sen, B.; Xie, Z.; Yen, S.S.; Miller, S.; Bas, G.; Styner, M.; Rubin, C.T.; Judex, S.; et al. Cell Mechanosensitivity to Extremely Low-Magnitude Signals Is Enabled by a LINCed Nucleus. Stem Cells 2015, 33, 2063–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, C.; Turner, A.S.; Bain, S.; Mallinckrodt, C.; McLeod, K. Low mechanical signals strengthen long bones. Nature 2001, 412, 603–604. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.T.; Capilla, E.; Luu, Y.K.; Busa, B.; Crawford, H.; Nolan, D.J.; Mittal, V.; Rosen, C.J.; Pessin, J.E.; Judex, S. Adipogenesis is inhibited by brief, daily exposure to high-frequency, extremely low-magnitude mechanical signals. Proc. Natl. Acad. Sci. USA 2007, 104, 17879–17884. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.A.; Thutupalli, S.; Fourrière, A.; Hallatschek, O. Chimera states in mechanical oscillator networks. Proc. Natl. Acad. Sci. USA 2013, 110, 10563–10567. [Google Scholar] [CrossRef] [Green Version]
- Matsuhashi, M.; Pankrushina, A.N.; Takeuchi, S.; Ohshima, H.; Miyoi, H.; Endoh, K.; Murayama, K.; Watanabe, H.; Endo, S.; Tobi, M.; et al. Production of sound waves by bacterial cells and the response of bacterial cells to sound. J. Gen. Appl. Microbiol. 2005, 44, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Babayi, T.; Riazi, G.H. The Effects of 528 Hz Sound Wave to Reduce Cell Death in Human Astrocyte Primary Cell Culture Treated with Ethanol. J. Addict. Res. Ther. 2017, 8, 1000335. [Google Scholar] [CrossRef]
- Lestard, N.R.; Capella, M.A.M. Exposure to Music Alters Cell Viability and Cell Motility of Human Nonauditory Cells in Culture. Evid.-Based Complement. Altern. Med. 2016, 2016, 6849473. [Google Scholar] [CrossRef] [Green Version]
- Lestard, N.D.R.; Valente, R.C.; Lopes, A.G.; Capella, M.A.M. Direct effects of music in non-auditory cells in culture. Noise Health 2013, 15, 307–314. [Google Scholar] [CrossRef]
- Lenzi, P.; Frenzili, G.; Gesi, M.; Ferrucci, M.; Lazzeri, G.; Fornai, F.; Nigro, M. DNA damage associated with ultrastructural alterations in rat myocardium after loud noise exposure. Environ. Health Perspect. 2003, 111, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, E.; Borrecho, G.; Oliveira, P.; de Matos, A.P.A.; Brito, J.; Águas, A.; Martins dos Santos, J. Effects of low-frequency noise on cardiac collagen and cardiomyocyte ultrastructure: An immunohistochemical and electron microscopy study. Int. J. Clin. Exp. Pathol. 2013, 6, 2333–2341. [Google Scholar] [PubMed]
- Naseer, S.M.; Manbachi, A.; Samandari, M.; Walch, P.; Gao, Y.; Zhang, Y.S.; Davoudi, F.; Wang, W.; Abrinia, K.; Cooper, J.M.; et al. Surface acoustic waves induced micropatterning of cells in gelatin methacryloyl (GelMA) hydrogels. Biofabrication 2017, 9, 15020. [Google Scholar] [CrossRef]
- Guo, F.; Li, P.; French, J.B.; Mao, Z.; Zhao, H.; Li, S.; Nama, N.; Fick, J.R.; Benkovic, S.J.; Huang, T.J. Controlling cell–cell interactions using surface acoustic waves. Proc. Natl. Acad. Sci. USA 2015, 112, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Haupt, A.; Minc, N. How cells sense their own shape—Mechanisms to probe cell geometry and their implications in cellular organization and function. J. Cell Sci. 2018, 131, jcs214015. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Havelka, D.; Kučera, O.; Deriu, M.A.; Cifra, M. Electro-acoustic behavior of the mitotic spindle: A semi-classical coarse-grained model. PLoS ONE 2014, 9, e86501. [Google Scholar] [CrossRef]
- Kučera, O.; Havelka, D.; Cifra, M. Vibrations of microtubules: Physics that has not met biology yet. Wave Motion 2017, 72, 13–22. [Google Scholar] [CrossRef]
- Wang, N.; Tytell, J.D.; Ingber, D.E. Mechanotransduction at a distance: Mechanically coupling the extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 2009, 10, 75–82. [Google Scholar] [CrossRef]
- Guzun, R.; Karu-Varikmaa, M.; Gonzalez-Granillo, M.; Kuznetsov, A.V.; Michel, L.; Cottet-Rousselle, C.; Saaremäe, M.; Kaambre, T.; Metsis, M.; Grimm, M.; et al. Mitochondria-cytoskeleton interaction: Distribution of β-tubulins in cardiomyocytes and HL-1 cells. Biochim. Biophys. Acta-Bioenergy 2011, 1807, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.V.; Javadov, S.; Guzun, R.; Grimm, M.; Saks, V. Cytoskeleton and regulation of mitochondrial function: The role of beta-tubulin II. Front. Physiol. 2013, 4, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robison, P.; Prosser, B.L. Microtubule mechanics in the working myocyte. J. Physiol. 2017, 595, 3931–3937. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, V.; Nijenkamp, L.L.A.; Regan, J.A.; van der Velden, J. The physiological role of cardiac cytoskeleton and its alterations in heart failure. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 700–722. [Google Scholar] [CrossRef] [Green Version]
- Davani, E.Y.; Dorscheid, D.R.; Lee, C.-H.; van Breemen, C.; Walley, K.R. Novel regulatory mechanism of cardiomyocyte contractility involving ICAM-1 and the cytoskeleton. Am. J. Physiol. Circ. Physiol. 2004, 287, H1013–H1022. [Google Scholar] [CrossRef] [Green Version]
- Dal Lin, C.; Radu, C.M.; Vitiello, G.; Romano, P.; Polcari, A.; Iliceto, S.; Simioni, P.; Tona, F. In vitro effects on cellular shaping, contratility, cytoskeletal organization and mitochondrial activity in HL1 cells after different sounds stimulation. A qualitative pilot study and a theoretical physical model. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kamkin, A.; Kiseleva, I. (Eds.) Mechanosensitivity and Mechanotransduction; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-90-481-9880-1. [Google Scholar]
- Sum, C.S.; Nickischer, D.; Lei, M.; Weston, A.; Zhang, L.; Schweizer, L. Establishing a High-content Analysis Method for Tubulin Polymerization to Evaluate Both the Stabilizing and Destabilizing Activities of Compounds. Curr. Chem. Genom. Transl. Med. 2014, 8, 16–26. [Google Scholar] [CrossRef]
- Romani, P.; Valcarcel-Jimenez, L.; Frezza, C.; Dupont, S. Crosstalk between mechanotransduction and metabolism. Nat Rev Mol. Cell Biol. 2021, 22, 22–38. [Google Scholar] [CrossRef]
- Ventura, C.; Graves, M.; Bergonzoni, A.; Tassinari, R.; Cavallini, C. Cell melodies: When sound speaks to stem cells. CellR4 2017, 5, e2331. [Google Scholar]
- Chang, C.T.; Bostwick, J.B.; Steen, P.H.; Daniel, S. Substrate constraint modifies the Rayleigh spectrum of vibrating sessile drops. Phys. Rev. E-Stat. Nonlinear Soft Matter Phys. 2013, 88, 23015. [Google Scholar] [CrossRef] [Green Version]
- Kučera, O.; Havelka, D. Mechano-electrical vibrations of microtubules-Link to subcellular morphology. BioSystems 2012, 109, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Misawa, T.; Takahama, M.; Kozaki, T.; Lee, H.; Zou, J.; Saitoh, T.; Akira, S. Microtubule-driven spatial arrangement of mitochondria promotes activation of the NLRP3 inflammasome. Nat. Immunol. 2013, 14, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Peleg, B.; Disanza, A.; Scita, G.; Gov, N. Propagating cell-membrane waves driven by curved activators of actin polymerization. PLoS ONE 2011, 6, e18635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierkes, K.; Sumi, A.; Solon, J.; Salbreux, G. Spontaneous Oscillations of Elastic Contractile Materials with Turnover. Phys. Rev. Lett. 2014, 113, 148102. [Google Scholar] [CrossRef] [PubMed]
- Bányai, L.A. A Compendium of Solid State Theory; Springer: Cham, Switzerland, 2018; ISBN 978-3-319-78612-4. [Google Scholar]
- Maldovan, M. Sound and heat revolutions in phononics. Nature 2013, 503, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Meijer, D.K.F.; Geesink, H.J.H. Guided Folding of Life’s Proteins in Integrate Cells with Holographic Memory and GM-Biophysical Steering. Open J. Biophys. 2018, 8, 117–154. [Google Scholar] [CrossRef] [Green Version]
- Meijer, D.K.F.; Geesink, J.H. Phonon Guided Biology. Architecture of Life and Conscious Perception Are Mediated by Toroidal Coupling of Phonon, Photon and Electron Information Fluxes at Discrete Eigenfrequencies. NeuroQuantology 2016, 14, 718–755. [Google Scholar] [CrossRef] [Green Version]
- Acbas, G.; Niessen, K.A.; Snell, E.H.; Markelz, A.G. Optical measurements of long-range protein vibrations. Nat. Commun. 2014, 5, 3076. [Google Scholar] [CrossRef]
- Del Giudice, E.; Doglia, S.; Milani, M.; Vitiello, G. A quantum field theoretical approach to the collective behavior of biological systems. Nucl. Phys. B 1985, 251, 375–400. [Google Scholar] [CrossRef]
- Del Giudice, E.; Doglia, S.; Milani, M.; Vitiello, G. Electromagnetic field and spontaneous symmetry breakdown in biological matter. Nucl. Phys. B 1986, 275, 185–199. [Google Scholar] [CrossRef]
- Del Giudice, E.; Doglia, S.; Milani, M.; Vitiello, G. Structure, correlations and electromagnetic interactions in living matter: Theory and applications. In Biological Coherence and Response to External Stimuli; Fröhlich, H., Ed.; Springer: Berlin, Germany, 1988; pp. 49–64. [Google Scholar] [CrossRef]
- Gerlich, S.; Eibenberger, S.; Tomandl, M.; Nimmrichter, S.; Hornberger, K.; Fagan, P.J.; Tüxen, J.; Mayor, M.; Arndt, M. Quantum interference of large organic molecules. Nat. Commun. 2011, 2, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cheng, J.; Qin, Y.X. Mechanobiological modulation of cytoskeleton and calcium influx in osteoblastic cells by short-term focused acoustic radiation force. PLoS ONE 2012, 7, e38343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavromatos, N.E. Non-linear dynamics in biological microtubules: Solitons and dissipation-free energy transfer. J. Phys. 2017, 880, 12010. [Google Scholar] [CrossRef] [Green Version]
- Mańka, R.; Ogrodnik, B. A model of soliton transport along microtubules. J. Biol. Phys. 1991, 18, 185–189. [Google Scholar] [CrossRef]
- Abdalla, E.; Maroufi, B.; Melgar, B.C.; Sedra, M.B. Information transport by sine-Gordon solitons in microtubules. Phys. A Stat. Mech. Appl. 2001, 301, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Every, A.G.; Tománek, D. Long-wavelength deformations and vibrational modes in empty and liquid-filled microtubules and nanotubes: A theoretical study. Phys. Rev. B 2017, 95, 205407. [Google Scholar] [CrossRef] [Green Version]
- Prodan, E.; Prodan, C. Topological Phonon Modes and Their Role in Dynamic Instability of Microtubules. Phys. Rev. Lett. 2009, 103, 248101. [Google Scholar] [CrossRef]
- Umezawa, H. Advanced Field Theory: Micro, Macro, and Thermal Physics; AIP: New York, NY, USA, 1993; ISBN 978-1-56396-456-5. [Google Scholar]
- Blasone, M.; Jizba, P.; Vitiello, G. Quantum Field Theory and Its Macroscopic Manifestations; Imperial College Press: London, UK, 2011; ISBN 9781848162808. [Google Scholar]
- Prodan, E.; Dobiszewski, K.; Kanwal, A.; Palmieri, J.; Prodan, C. Dynamical Majorana edge modes in a broad class of topological mechanical systems. Nat. Commun. 2017, 8, 14587. [Google Scholar] [CrossRef]
- Goldstone, J.; Salam, A. Weinberg S Broken Symmetries. Phys. Rev 1962, 127, 965–970. [Google Scholar] [CrossRef]
- Del Giudice, E.; Vitiello, G. The role of the electromagnetic field in the formation of domains in the process of symmetry breaking phase transitions. Phys. Rev. A 2006, 74, 022105. [Google Scholar] [CrossRef] [Green Version]
- Celeghini, E.; Rasetti, M.; Vitiello, G. Quantum dissipation. Ann. Phys. 1992, 215, 156–170. [Google Scholar] [CrossRef]
- Vitiello, G. Dissipation and memory capacity in the quantum brain model. Int. J. Mod. Phys. B 1995, 9, 973–989. [Google Scholar] [CrossRef]
- Engel, G.S.; Calhoun, T.R.; Read, E.L.; Ahn, T.-K.; Mančal, T.; Cheng, Y.-C.; Blankenship, R.E.; Fleming, G.R. Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature 2007, 446, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Cogdell, R.J.; Coker, D.F.; Duan, H.-G.; Hauer, J.; Kleinekathöfer, U.; Jansen, T.L.C.; Mančal, T.; Miller, R.J.D.; Ogilvie, J.P.; et al. Quantum biology revisited. Sci. Adv. 2020, 6, eaaz4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shayeghi, A.; Rieser, P.; Richter, G.; Sezer, U.; Rodewald, J.H.; Geyer, P.; Martinez, T.J.; Arndt, M. Matter-wave interference of a native polypeptide. Nat. Commun. 2020, 11, 1447. [Google Scholar] [CrossRef] [Green Version]
- Davydov, A.S. Solitons in Molecular Systems; Mathematics and Its Applications; Springer: Dordrecht, The Netherlands, 1985; Volume 4, ISBN 978-94-017-3027-3. [Google Scholar]
- Brizhik, L.; Eremko, A.; Cruzeiro-Hansson, L.; Olkhovska, Y. Soliton dynamics and Peierls-Nabarro barrier in a discrete molecular chain. Phys. Rev. B 2000, 61, 1129–1141. [Google Scholar] [CrossRef]
- Brizhik, L. Influence of electromagnetic field on soliton-mediated charge transport in biological systems. Electromagn. Biol. Med. 2015, 34, 123–132. [Google Scholar] [CrossRef]
- Brizhik, L.; Chiappini, E.; Stefanini, P.; Vitiello, G. Modeling Meridians Within the Quantum Field Theory. J. Acupunct. Meridian Stud. 2019, 12, 29–36. [Google Scholar] [CrossRef]
- Heimburg, T.; Jackson, A.D. On soliton propagation in biomembranes and nerves. Proc. Natl. Acad. Sci. USA 2005, 102, 9790–9795. [Google Scholar] [CrossRef] [Green Version]
- McDermott, M.L.; Vanselous, H.; Corcelli, S.A.; Petersen, P.B. DNA’s Chiral Spine of Hydration. ACS Cent. Sci. 2017, 3, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Marburger, J.H. Self-focusing: Theory. Prog. Quantum Electron. 1975, 4, 35–110. [Google Scholar] [CrossRef]
- Chiao, R.; Gustafson, T.; Kelley, P. Self-Focusing of Optical Beams. In Self Focusing: Past and Present; Boyd, R.W., Lukishova, S.G., Shen, Y.R., Eds.; Springer: New York, NY, USA, 2009; pp. 129–143. ISBN 978-0-387-32147-9. [Google Scholar]
- Zakharov, V.; Shabat, A. Exact theory of two-dimensional self-focusing and automodulation of waves in nonlinear media. Sov. J. Exp. Theory Phys. 1971, 61, 118–134. [Google Scholar]
- Benias, P.C.; Wells, R.G.; Sackey-Aboagye, B.; Klavan, H.; Reidy, J.; Buonocore, D.; Miranda, M.; Kornacki, S.; Wayne, M.; Carr-Locke, D.L.; et al. Structure and Distribution of an Unrecognized Interstitium in Human Tissues. Sci. Rep. 2018, 8, 4947. [Google Scholar] [CrossRef] [PubMed]
- Askar’yan, G. The self-focusing effect. Sov. Phys. Uspekhi 1974, 16, 680. [Google Scholar] [CrossRef]
- Mandoli, D.F.; Briggs, W.R. Optical properties of etiolated plant tissues. Proc. Natl. Acad. Sci. USA 1982, 79, 2902–2906. [Google Scholar] [CrossRef] [Green Version]
- Vitiello, G. Fractals, coherent states and self-similarity induced noncommutative geometry. Phys. Lett. A 2012, 376, 2527–2532. [Google Scholar] [CrossRef] [Green Version]
- Vitiello, G. Coherent States, Fractals And Brain Waves. New Math. Nat. Comput. 2009, 5, 245–264. [Google Scholar] [CrossRef] [Green Version]
- Vitiello, G. On the Isomorphism between Dissipative Systems, Fractal Self-Similarity and Electrodynamics. Toward an Integrated Vision of Nature. Systems 2014, 2, 203–216. [Google Scholar] [CrossRef]
- ASTM F2149-16. Standard Test Method for Automated Analyses of Cells—The Electrical Sensing Zone Method of Enumerating and Sizing Single Cell Suspensions; ASTM International: West Conshohocken, PA, USA, 2016; Available online: www.astm.org (accessed on 15 November 2020).
- Pokorný, J.; Pokorný, J.; Kobilková, J. Postulates on electromagnetic activity in biological systems and cancer. Integr. Biol. 2013, 5, 1439–1446. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Meditation Music | Mantra | “Ti Amo” Signal | Commercial Music | Noise | “Ti Odio” Signal | |
|---|---|---|---|---|---|---|---|
| Contractiona.u. | 268 (265–270) | 301 (300–304) | 306 (304–312) | 301 (299–303) | 244 (223–247) | 250 (245–255) | 241 (240–246) |
| p | - | <0.01 | <0.001 | <0.01 | <0.001 | <0.01 | <0.001 |
| A | B | C | |
|---|---|---|---|
| Phalloidin | 17.8 ± 4.3 #,∗ | 30.3 ± 5.4 #,• | 6.1 ± 2.9 ∗,• |
| Beta-actin | 1.5 ± 0.3 #,∗ | 7.4 ± 1.9 #,• | 2.7 ± 0.6 ∗,• |
| Alpha-actinin-1 | 1.5 ± 0.2 #,∗ | 3.1 ± 0.7 #,• | 1.0 ± 0.2 ∗,• |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dal Lin, C.; Radu, C.M.; Vitiello, G.; Romano, P.; Polcari, A.; Iliceto, S.; Simioni, P.; Tona, F. Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model. Int. J. Mol. Sci. 2021, 22, 156. https://doi.org/10.3390/ijms22010156
Dal Lin C, Radu CM, Vitiello G, Romano P, Polcari A, Iliceto S, Simioni P, Tona F. Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model. International Journal of Molecular Sciences. 2021; 22(1):156. https://doi.org/10.3390/ijms22010156
Chicago/Turabian StyleDal Lin, Carlo, Claudia Maria Radu, Giuseppe Vitiello, Paola Romano, Albino Polcari, Sabino Iliceto, Paolo Simioni, and Francesco Tona. 2021. "Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model" International Journal of Molecular Sciences 22, no. 1: 156. https://doi.org/10.3390/ijms22010156
APA StyleDal Lin, C., Radu, C. M., Vitiello, G., Romano, P., Polcari, A., Iliceto, S., Simioni, P., & Tona, F. (2021). Sounds Stimulation on In Vitro HL1 Cells: A Pilot Study and a Theoretical Physical Model. International Journal of Molecular Sciences, 22(1), 156. https://doi.org/10.3390/ijms22010156