1. Introduction
Cell-to-cell contact is essential for communication and development of multicellular organisms. A prerequisite is the passage through membranes. That way, molecular exchange and information flow is regulated via hormones, membrane proteins and pores. In plants, the rigid cell walls prevent large membrane contact areas between protoplasts. Only plasmodesmata, minute channels between adjacent cells, form direct connections. Often, molecular data of the proteins involved are manifold but there is a lack of knowledge on functional and structural information.
1.1. The Wall between Cells
In turgid cells, the cortical cytoplasm and the plasma membrane (PM) is pushed towards the cell wall by the turgor pressure of the central vacuole [
1]. Thus, the cortical membranes are closely aligned with the cell wall and a structural or visual separation is therefore difficult. Hence, plant cell biologists use plasmolysis to induce physical detachment of the living protoplast from the cell wall [
2]. Plasmolysis was first described by de Vries in 1887 [
3]. Exposure of plant tissue to hyperosmotic salt or sugar solutions of about 0.8–1 M cause water efflux from the vacuole by osmosis. This water loss results in the shrinking of the protoplast and eventually the detachment of the PM from the cell wall. A reticulate network of endoplasmatic reticulum (ER) and PM remains attached to the cell wall. It is known as Hechtian reticulum [
4]. Fine strands (Hechtian strands) maintain a connection of the Hechtian reticulum with the plasmolyzed protoplast [
5,
6]. The phenomenon of plasmolysis is frequently used in plant cell biology classes to demonstrate the PM, tonoplast and cytoplasm and the cell wall as separate structures [
1,
7]. Additionally, plasmolysis is a useful tool to test the viability of plant cells as only in living cells, the semipermeable membranes are intact and allow for plasmolysis. Dead cells do not plasmolyze.
Here, we used plasmolysis for the analysis of structural details at the cell wall between two neighboring cells. The focus was on the attachment/detachment of the PM (stained with FM4-64) and the ER (labeled by a green fluorescent protein (GFP)-tag). In a first step, we plasmolyzed protonema cells and defined various forms of detachment and their statistical frequency. Secondly, we looked at the two sides of the cell wall like at a door, viewed from inside the cell of neighbor one and neighbor two, respectively.
1.2. The Microscopic View
In the light microscope, samples are usually seen in top view. However, this microscopic view only shows the narrowest side of the walls between cell neighbors. A 90° tilt would be necessary to look at the whole disc of the wall. This is possible by confocal laser scanning microscopy (CLSM) and optical sectioning of the probe. The resulting image stacks can be tilted at arbitrary angles thereby allowing a view from inside the cell onto the wall. If labeling for specific structures has been applied, segmentation and 3D reconstructions can result in additional information, e.g., by the separation of individual structures. In our case, we imaged the labeled ER and PM in separate channels. Likewise, any other protein of interest could be imaged if it can be fluorescently marked. AMIRA® was used to generate 3D reconstructions, segmentations and “helicopter views“; here, with a focus on the cell wall between two neighbors to analyze structural details of membrane-wall contact sites.
1.3. Moss Protonema as Model Cells
In search of the optimal samples, we chose filamentous moss protonemata [
8]. These consist of a single line of cells; branching occurs only occasionally [
9]. Hence, two cells normally possess only one connecting cell wall. Chloronema cells show more chloroplasts, straight cell walls and a slightly bigger diameter than caulonema cells with oblique walls [
10]. In both cases, the diameter is ideal for optical stacks of the whole cell by confocal microscopy without further mechanical sectioning. Although mosses are evolutionary older than seed plants, the structural features of the cells are very similar. This also applies for plasmolysis. Additionally, in the moss model
Physcomitrium (Physcomitrella) patens, various cell lines with GFP-tagged proteins are available (
https://sites.dartmouth.edu/bezanillalab) to allow specific tracing of proteins in living samples.
In the present study, we report on the 3D reconstruction and segmentation of membranes at the adjacent wall between two neighboring cells. We give examples on the possibilities of this 3D technique to visualize structural details. Thereby, we want to inspire further studies in the field of plant cell imaging.
2. Results
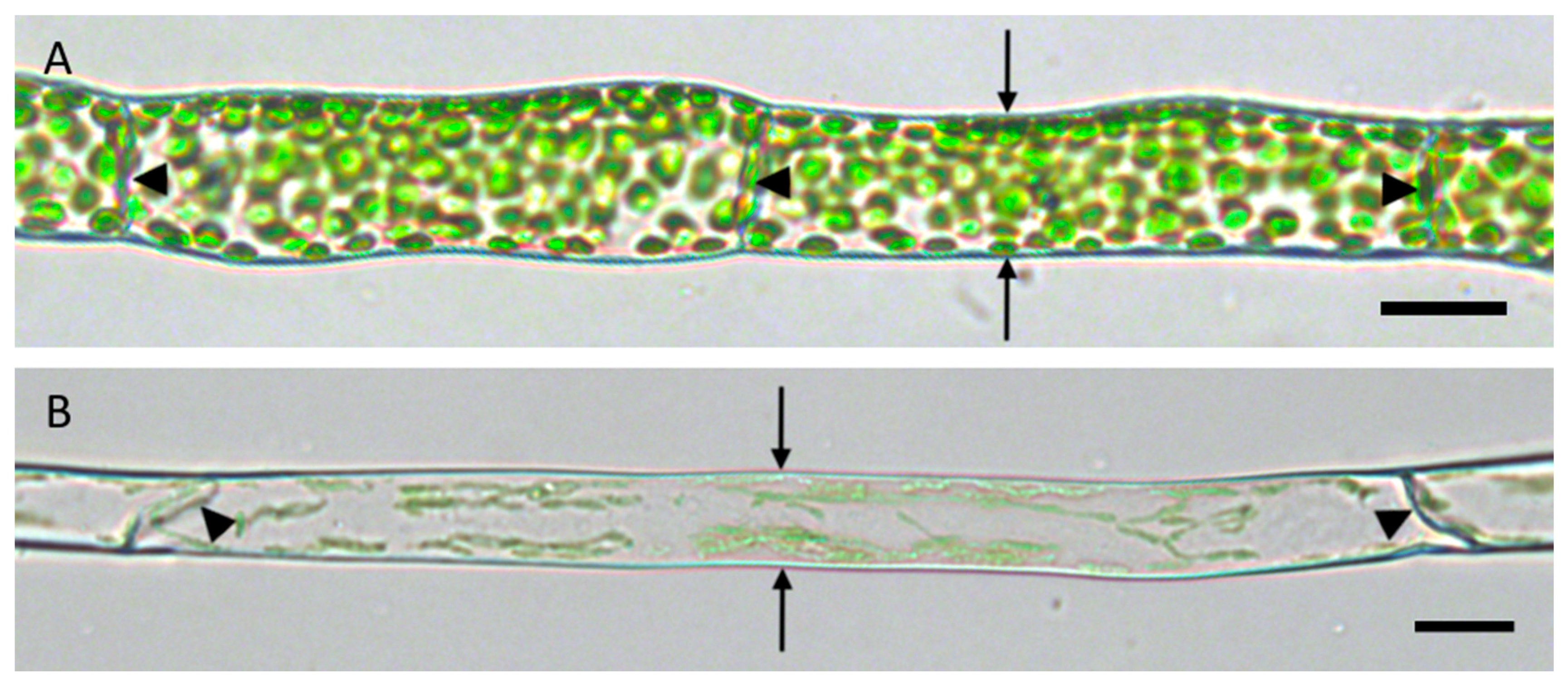
The filamentous protonemata of
P. patens consist of interconnected, cylindrical cells with occasional branching. Normally, two neighbors share a common middle lamella between their adjoining cell walls that is separating the individual cells. Protonemata can be distinguished into chloronema (
Figure 1A) and caulonema (
Figure 1B). Chloronema cells are shorter and thicker than caulonema cells. Additionally, there are differences in the abundance of chloroplasts and the inclination of the connecting cell walls. Chloronemata contain more chloroplasts and possess straight cell walls with orthogonal orientation to the growth direction. Caulonema cells are characterized by fewer chloroplasts and oblique cell walls (
Figure 1).
In turgid protonemata, the PM and the cortical cytoplasm are in close alignment with the cell wall at all sides. The cortical ER consists of a polygonal network of fine tubules and sheets [
11]. It has been shown to be in immediate contact with the PM [
12].
Unless it is branching or a tip cell, each protonema cell has only two neighbors, one younger and one older cell, which are connected by a tilted (i.e., caulonema) or straight (i.e., chloronema) cell wall, respectively (
Figure 1). It is evident that these walls are essential for information flow and exchange within the thread. Disconnection would lead to cellular isolation and a lack of transport. Here, we used plasmolysis as a tool of osmotic water loss to investigate if and how the living protoplasts detached from the cell wall. A special focus was on the essential connection between two neighbored cells and their adjoining cell wall.
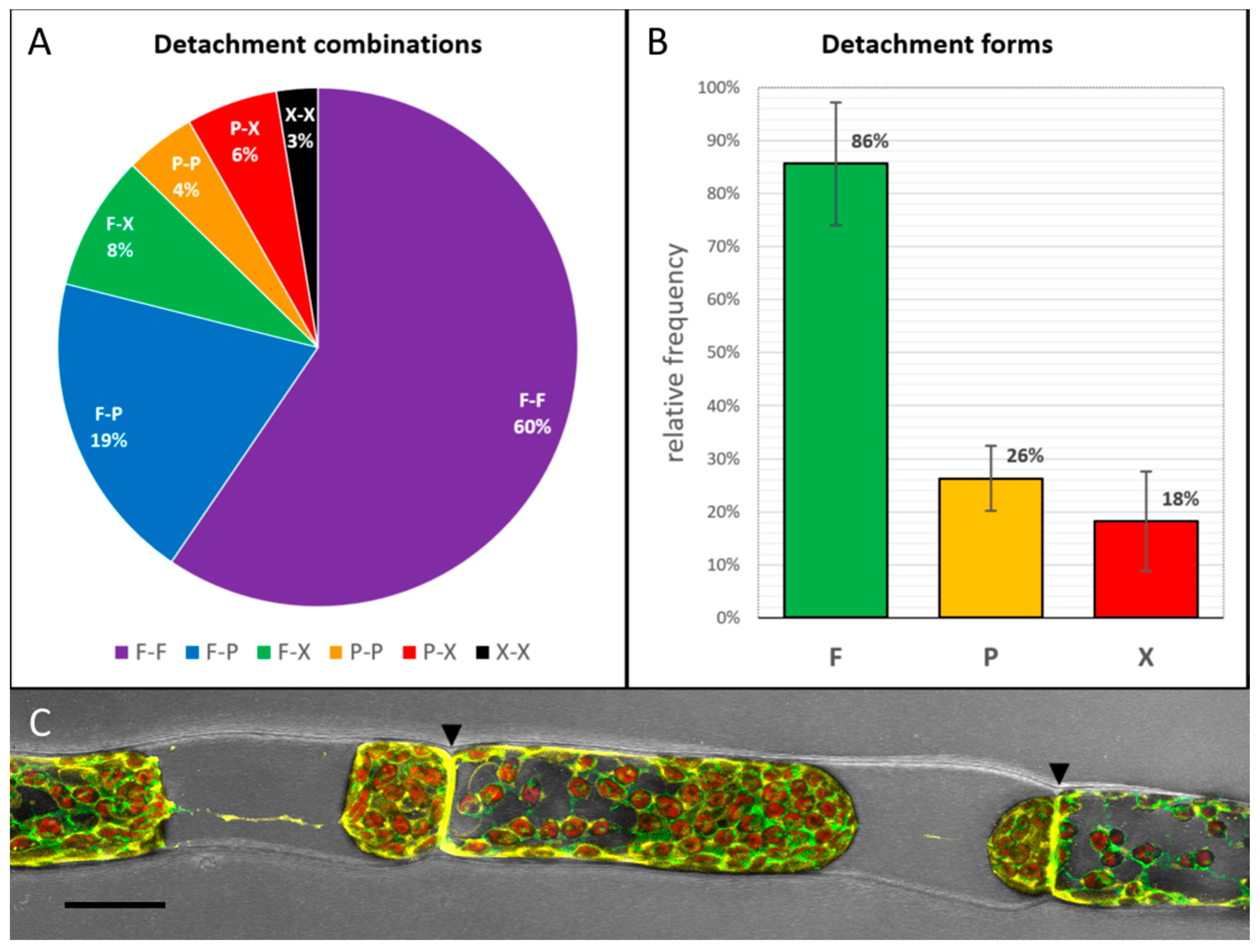
Preliminary plasmolysis experiments with 0.8 M mannitol resulted in three different detachment forms that were defined as “full attachment” (F), “partial detachment” (P) and “strong or full detachment” (X). Ergo, six different detachment combinations of protoplasts at each side of the connecting cell wall are possible: F-F, F-P, F-X, P-X and X-X. Consecutively, we examined 603 adjoining protonema cells after plasmolysis and their detachment forms (
Figure 2). Both, chloronema and caulonema cells plasmolyzed without visible differences.
In 60% of the observed cell pairs, the protoplasts of both cells stayed fully attached at the adjoining cell wall (F-F type;
Figure 2A,C), although the protoplasts detached from the lateral walls. These data confirm the existence of a strong membrane connection to the adjoining wall and the importance of the link between cell neighbors. The relative frequency of the three defined detachment forms showed that in 86% of the cell pairs at least one of the neighbors remained fully attached to the connecting cell wall (
Figure 2B). In contrast, a strong or full detachment of at least one of the protoplasts could only be observed in 18% of the investigated cell pairs.
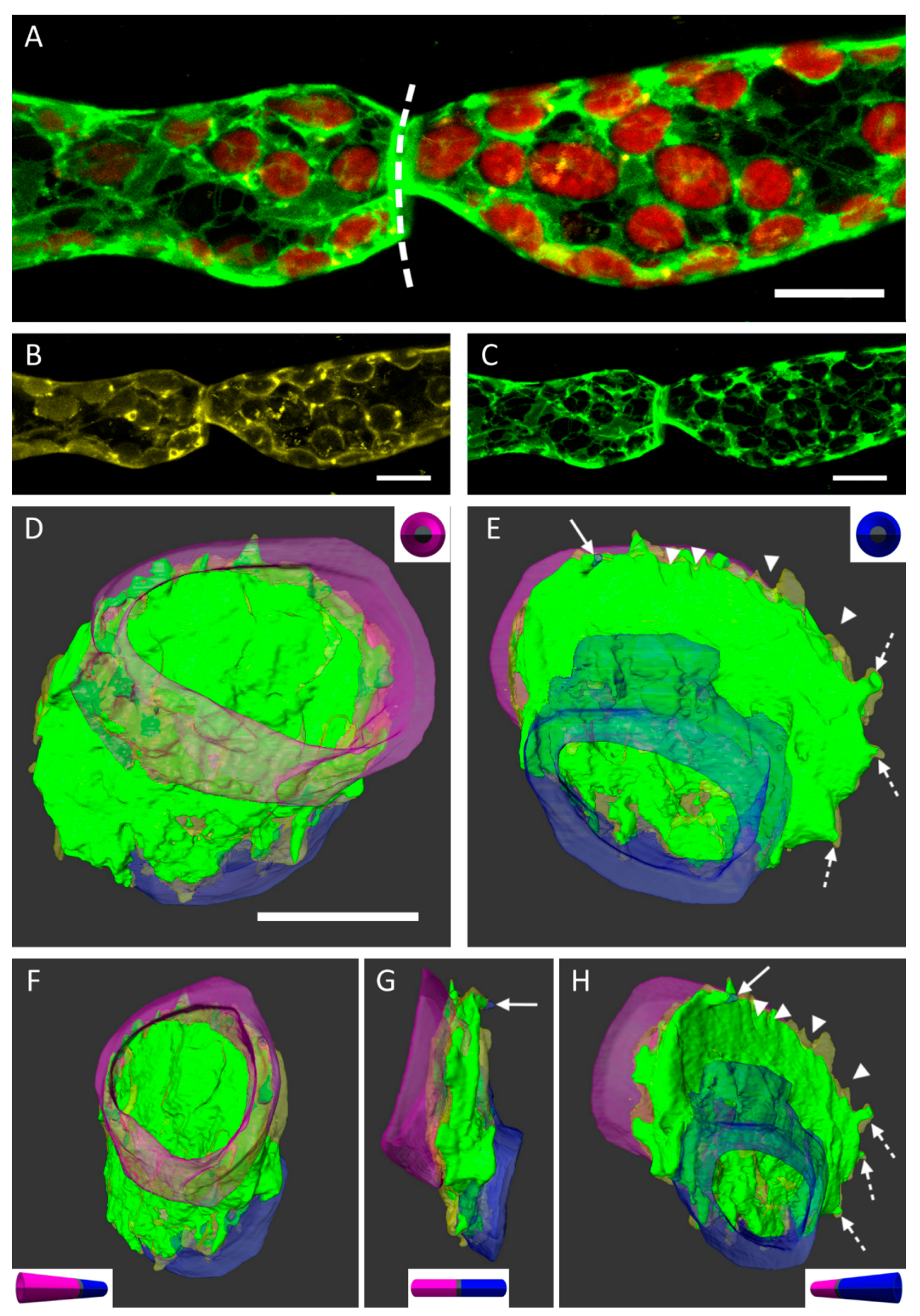
Further investigations of the situation at the cell walls between two cell neighbors and particular attachments of the PM and the ER were performed by high resolution confocal imaging (
Figure 3). We used a
P. patens cell line with a GFP-ER tag (eGFP-ER;
Figure 3B). The PM was stained with FM4-64, a commonly used styryl dye for cell membranes (
Figure 3C). The overlay of the confocal images in 3B and 3C, including the detection of chloroplasts by the autofluorescence of chlorophyll is shown in
Figure 3A. The cell wall between two adjacent protonema cells is depicted by the dashed line (
Figure 3A). Optical sectioning of the sample resulted in image z-stacks, which could be implemented to generate 3D reconstructions of the observed area using AMIRA
® software (
Figure 3D–H,
Figure 4 and
Figure 5). Segmentation and false-coloring the membranes gave further, more detailed insights to the three-dimensional arrangement of partially detached protoplasts.
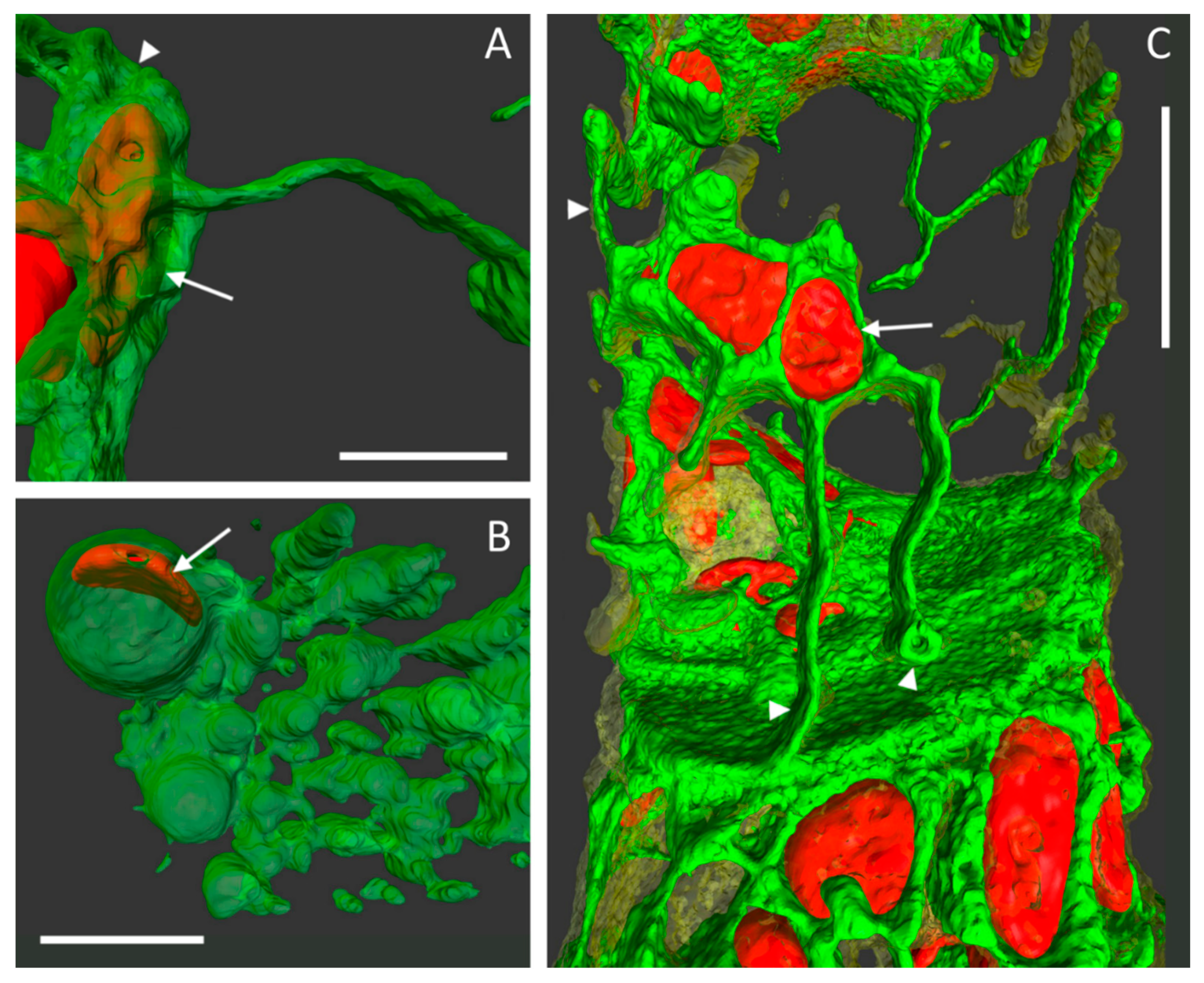
Figure 3D,E show the membrane attachments on each side of the common cell wall. Although the major part of the PM remained attached to the cell wall, the area of the two protoplasts was different at each side of the wall as seen in purple (
Figure 3D) and blue (
Figure 3E), respectively. Small notches (arrowheads) consisting of either ER or of PM appeared at the edges of the cell wall disc. Larger protrusions containing both membrane types were forming Hechtian strands (arrow) or were connected to the Hechtian reticulum (dotted arrows). The local shift of the two protoplasts at each side of the cell wall was only detected when comparing the situation imaged in front view (“purple” side;
Figure 3F) and then in side view (
Figure 3G). The number and location of emerging Hechtian strands (arrow) and Hechtian reticulum (dotted arrows) appeared by realizing tilted views from different angles (
Figure 3F,H). See also
supplemental Videos S1 and S2 to imaging the reconstructed cells in rotation.
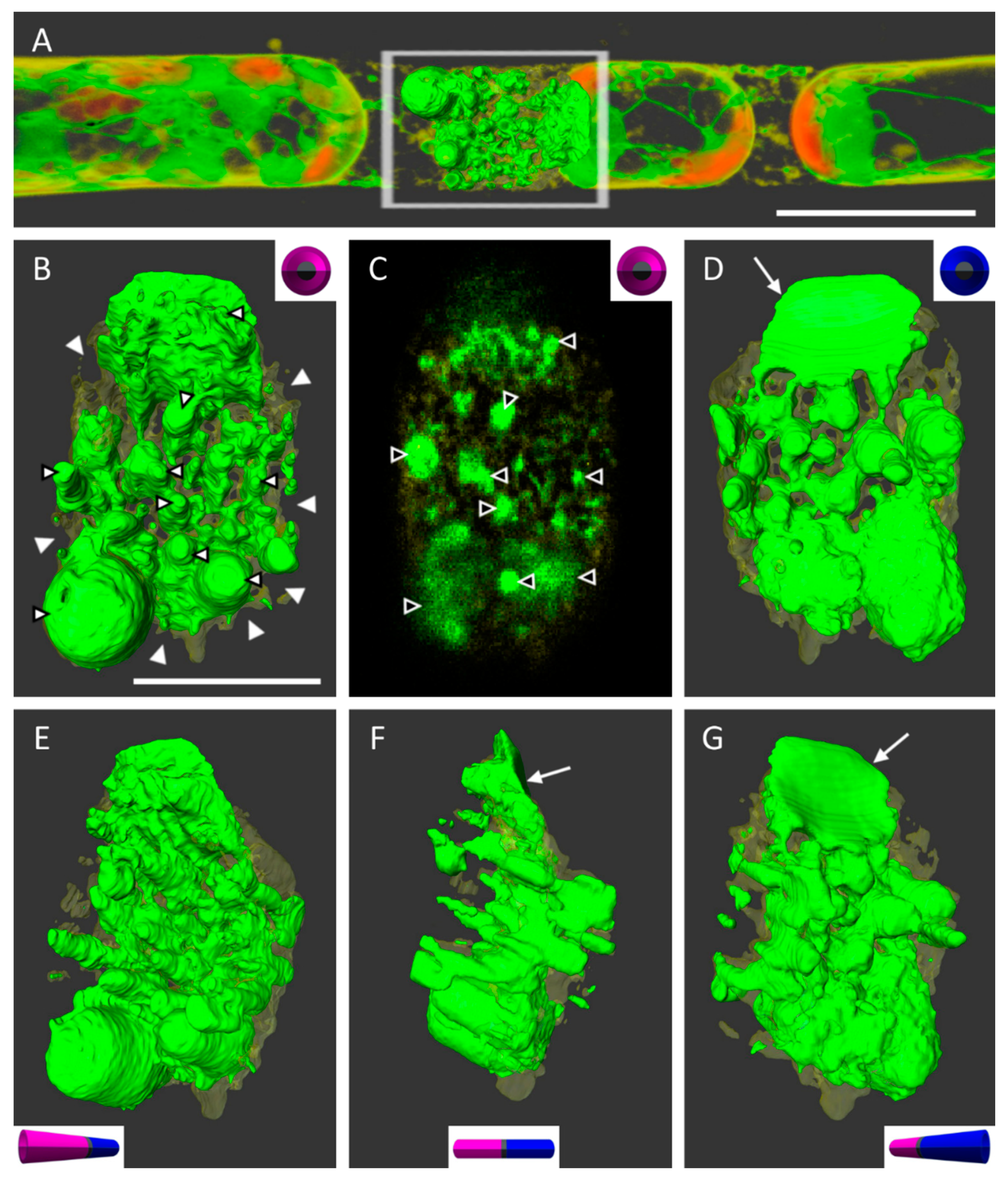
In conventional light microscopy, the cross walls of cells can only be observed in the top view, i.e., by looking at the narrow side of the wall. Only after generating a 3D surface, the area of interest can be turned and observed from various angles or from an inside-the-cell perspective. Due to technical restrictions and the multitude of chloroplasts in chloronemata, the resolution in the z-direction was relatively lower than in the xy direction. To increase the resolution and image quality, we therefore switched to caulonema cells with oblique walls that laid orthogonally to the scanning direction in the xy-plane (
Figure 4A). This way, any leftovers of ER and PM at the wall after plasmolysis could be analyzed more precisely (
Figure 4B–G). The detailed 3D reconstructions in
Figure 4 revealed the PM (in transparent yellow) surrounding the ER (shown in green) as a fine network at the adjoining wall between two cell neighbors. See also
supplemental Video S3.
After plasmolysis and the detachment of the main protoplast, ER membranes remained attached to the wall by forming big bulges. Virtually generated slices through the cell wall suggested the extension of some ER structures from one side to the other thereby connecting the cell neighbors (
Figure 4C).
3D reconstructions of confocal images also revealed great details of very delicate structures (
Figure 5). Networks and sheet-like components of the ER and PM and Hechtian strands could be clearly distinguished. As an example, we observed several times that ER tubules and membranes closely entangled a chloroplast (
Figure 5, arrows). Any Hechtian reticula and strands that remain in contact with the cell wall after plasmolysis could also be depicted in high resolution as tubules or little loops (
Figure 5, arrowheads).
3. Discussion
In biological systems, form is intrinsically connected to function. It is therefore essential to investigate the form of specific structures in order to understand their function. Modern, computer assisted imaging tools greatly support the visualization of structural details at all levels of complexity [
13,
14]. 3D presentations of biological structures inspire the imagination of the audience and are a fantastic asset, not only to solve scientific questions but also for teaching [
2].
In the present study, we focused on the structure of PM and ER membranes at the adjacent cell wall of two connected protonema cells. In interphase cells of
P. patens protonemata, the PM is usually closely aligned with the cell wall and the cortical ER forms are a dynamic network of tubules and sheets [
2,
15], as is also the case in cells of seed plants. A membrane-wall continuum between plant cell neighbors is formed by plasmodesmata [
16]. Additionally, various proteins like reticulons [
17,
18,
19], reticulon-like proteins and atlastins [
20] can shape the ER. Others provide ER–PM membrane contact sites [
6] and connect the ER and PM to the cell wall, also to the parts of the cell wall that do not possess adjacent cells. The nature of these linkers is not fully understood, but they could be the anchor points for Hechtian strands and reticulum at the cell wall. Additionally, Lang et al. proposed a physical anchor by cellulose fibers to link the plasma membrane to the cell wall [
5] and also the cytoskeleton is involved [
2,
6]. In all these cases, the visualization of the situation remained challenging in turgescent cells and therefore, we used plasmolysis for physical separation of the protoplast from the cell wall. This way, we could image the structural differences of cortical membranes and the cell wall. Additionally, segmentation of the z-stacks, image rotation and 3D reconstructions greatly help to distinguish structural differences. Particularly in the case of the adjacent walls in protonema cells, the connecting wall is normally observed from its narrow side as a line (
Figure 1) and not as a disc as is possible after 90° rotation of optical sections. To visualize the obscured parts, it is essential that the resolution in z direction is minimal and at least the same as in the xy direction in order to prevent image distortions.
Our data clearly show that cell–cell contact is essential in filamentous cells (
Figure 2). In 86% of the investigated cases, at least one of the neighbors remained fully attached to the cell wall after plasmolysis (
Figure 2B). A strong connection of the protoplast to one side of a cell has been described as “negativer Plasmolyseort” (location of negative plasmolysis; [
21,
22]). In
Avena sativa coleoptiles, the amount of plasmodesmata correlated positively with the adhesion of the plasmolyzed protoplast [
7]. Plasmodesmata have been proposed to provide connection sites for Hechtian strands and the Hechtian reticulum [
23]. Bryophytes also possess plasmodesmata with desmotubules [
24] and a membranous connection between cell neighbors in
P. patens is also confirmed here. Virtually generated slices of the cell wall in caulonema cells clearly show transversing membranes between the adjacent cells (
Figure 4). However, the reasons for the strong membrane–wall connection at the walls between cell neighbors in
P. patens protonemata are still not fully understood and it remains unclear why the contact is sometimes lost and how the disconnected/connected areas are defined.
As depicted in
Figure 3, segmentations of the protoplasts show differences in the connecting areas at each side of the cell wall; the purple protoplast was larger than the blue protoplast (
Figure 3E). There was a narrow part of the blue protoplast that maintained the contact in the center of the cell wall disc (
Figure 3F). Additionally, image tilts and reconstructions of the cell wall in
Figure 3 revealed strands of PM and ER and small notches at the edge of the cell wall disc. These were visible at the corner of protonema cells, distant from the plasmolyzed protoplasts. It remained to be tested if these structures colocalize with cytoskeletal elements like specific microtubules that bend at the cell corners [
25] or leftovers of zones that evolved from phragmoplast formation [
26].
High resolution z-stacks allow for 3D imaging of fine, structural details and the positioning of organelles within membranes (
Figure 5). Volume renderings in combination with varying transparency levels reveal that the chloroplasts are closely entangled by networks and sheets of ER membranes [
27], and that the membranes of ER and PM are distinct and separate from each other.
The images of the present study should serve as an example: depending on the scientific question, other proteins or structures of interest could be labeled and then analyzed as described here. Furthermore, segmentation and 3D imaging is not restricted to CLSM z-stacks. Segmentation software like AMIRA
® usually accept also data sets originating from other research fields or imaging techniques like micro computed tomography (µCT) [
28] transmission electron microscopy (TEM; [
29] or focused ion beam scanning electron microscopy (FIB-SEM), as shown for the green alga
Micrasterias detenticulata [
13] or
Klebsormidium sp. [
14].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





