Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations

 ,
,  , ,
, , 
Abstract
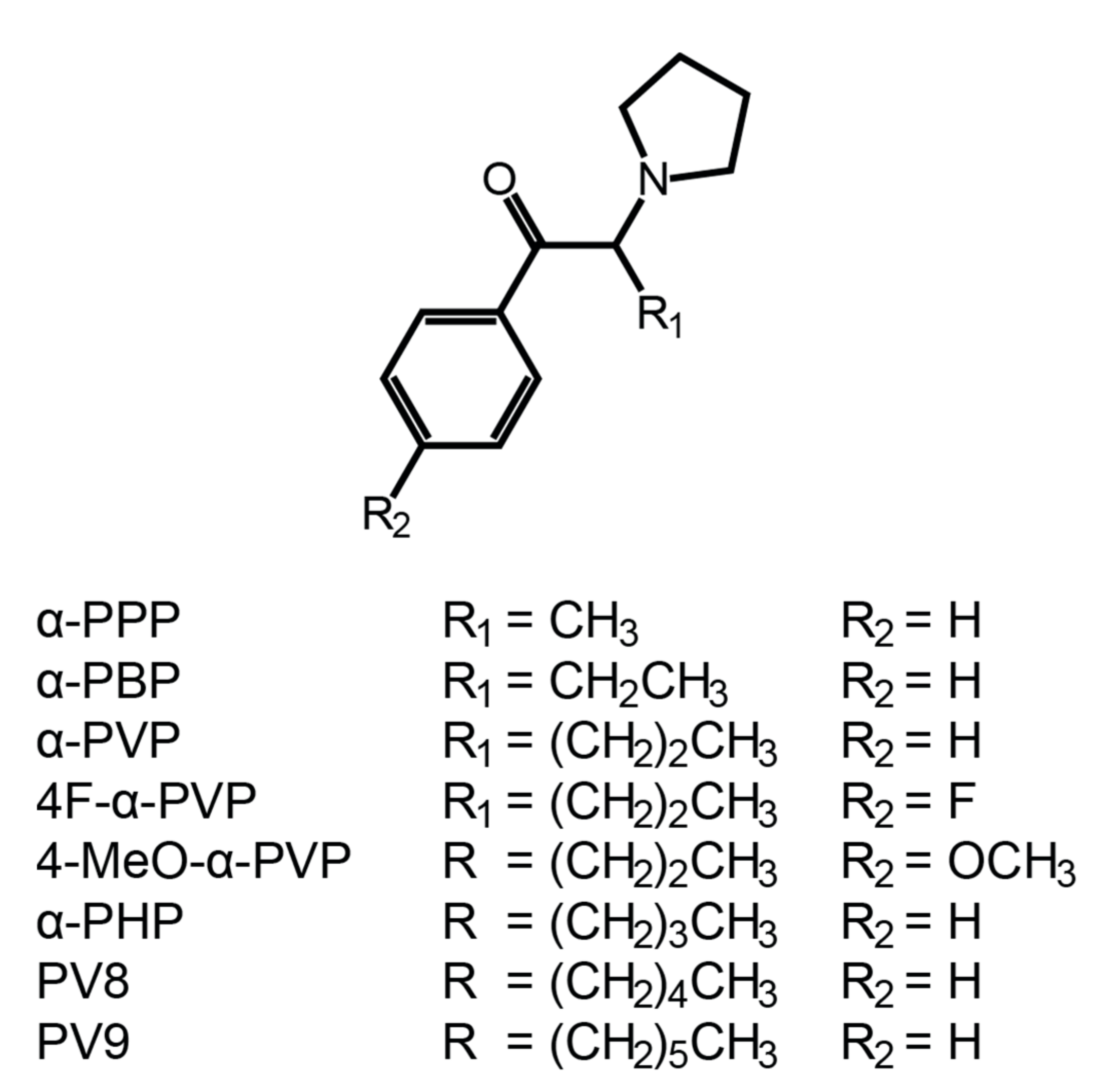
:1. Introduction
2. Results and Discussion
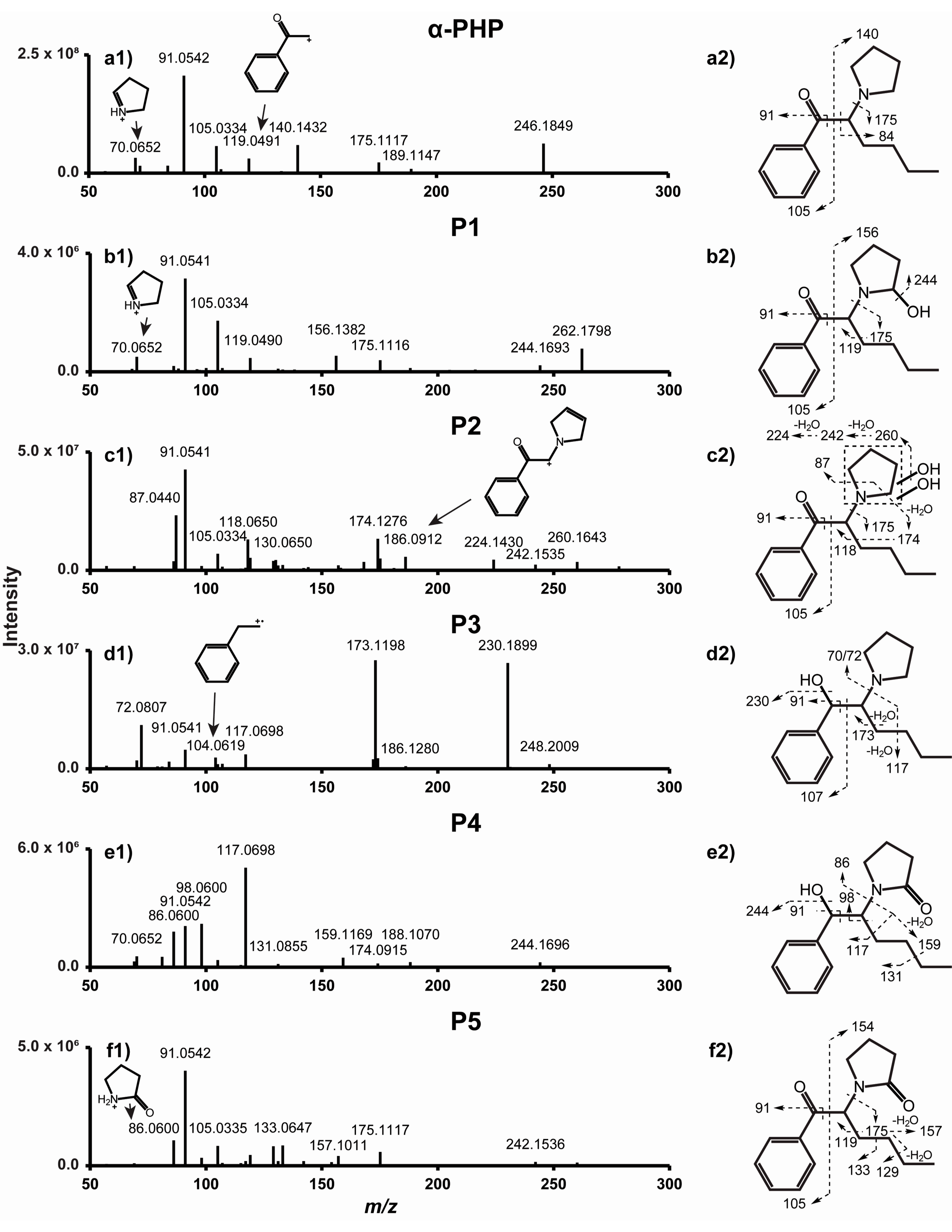
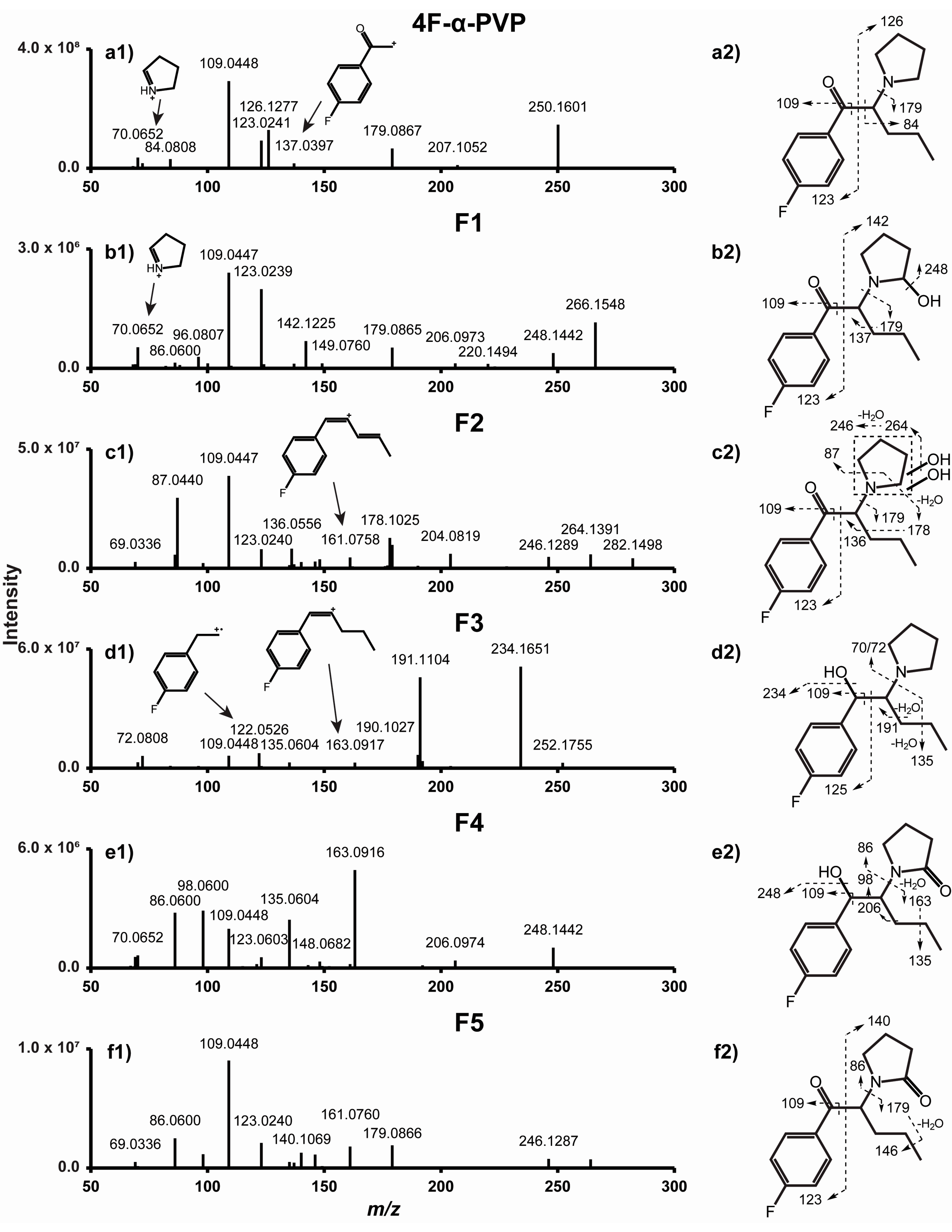
2.1. α-PHP and 4F-α-PVP MS/MS Fragmentation Patterns
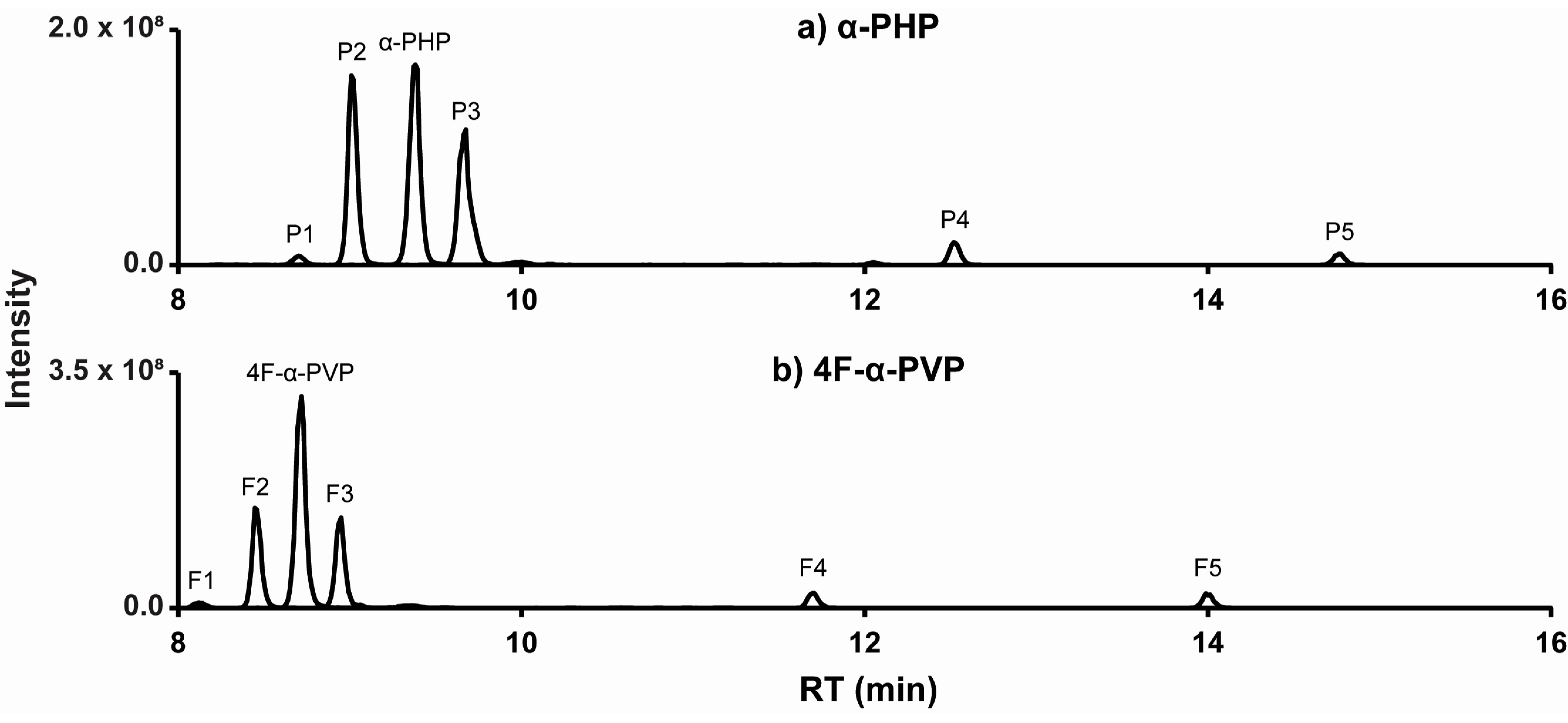
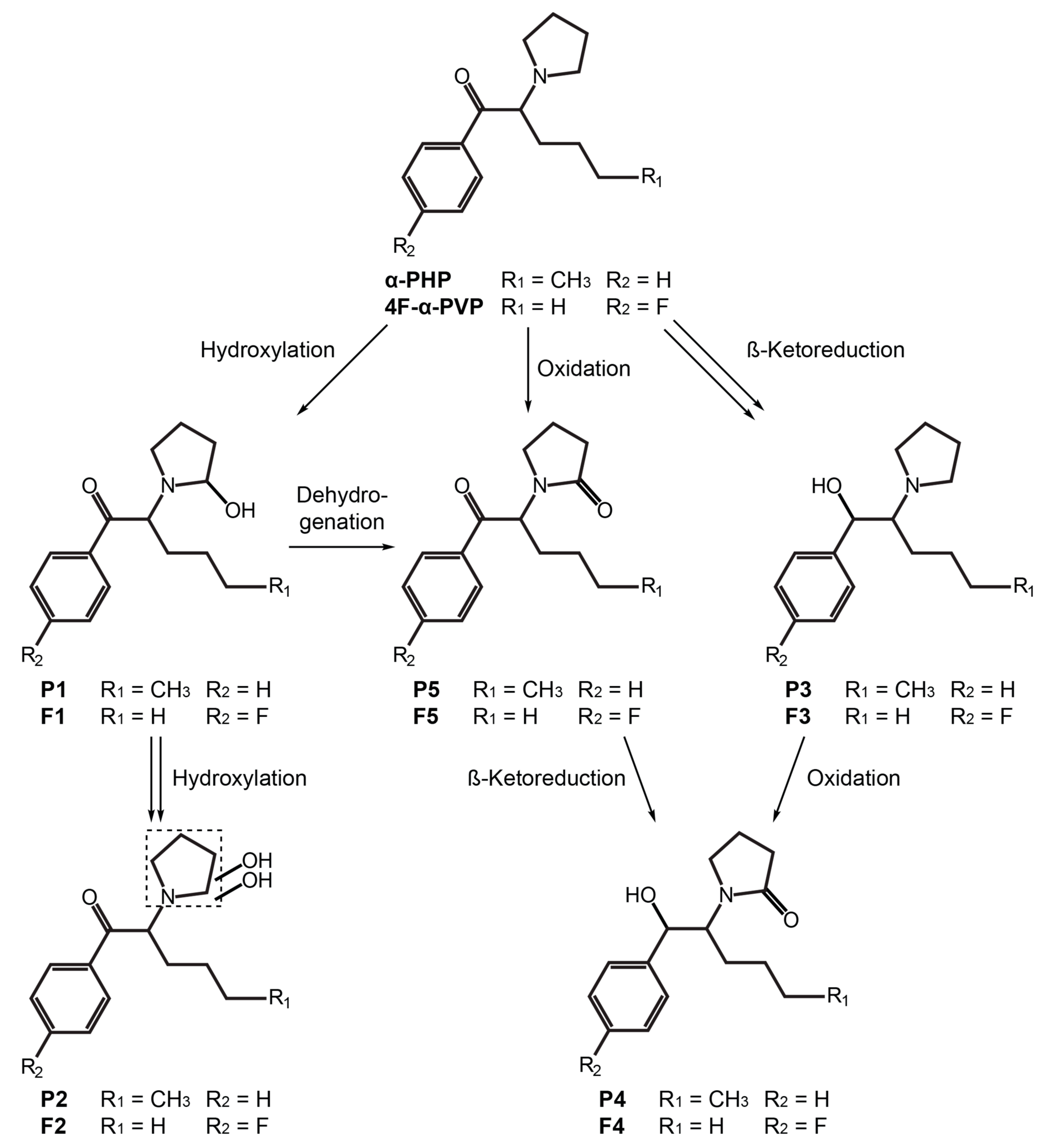
2.2. α-PHP Metabolism with Human Hepatocytes
2.2.1. β-Ketoreduction
2.2.2. Pyrrolidinyl Hydroxylation
2.2.3. Oxidation
2.3. 4F-α-PVP Metabolism with Human Hepatocytes
2.3.1. β-Ketoreduction
2.3.2. Pyrrolidinyl Hydroxylation
2.3.3. Oxidation
2.4. Optimal Targets for α-PHP and 4F-α-PVP
2.5. Analytical Considerations
3. Material and Methods
3.1. Chemicals and Reagents
3.2. Incubation with Pooled Human Hepatocytes
3.3. Sample Preparation
3.4. LC-HRMS/MS Parameters
3.4.1. LC Parameters
3.4.2. HRMS/MS Parameters
3.5. Metabolite Identification
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SC | Synthetic cathinone |
| NPS | Novel psychoactive substance |
| DAT | Dopamine transporter |
| NET | Norepinephrine transporter |
| SERT | Serotonin transporter |
| MDMA | 3″,4″-Methylenedioxymethamphetamine |
| MDPV | Methylenedioxypyrovalerone |
| α-PHP | α-pyrrolidinohexaphenone |
| 4F-α-PVP | 4″-Fluoro-α-pyrrolidinovalerophenone |
| α-PVP | α-Pyrrolidinovalerophenone |
| MDPH | 3″,4″-Methylenedioxy-α-pyrrolidinohexiophenone |
| EMCDDA | European Monitoring Centre for Drugs and Drug Addiction |
| UNODC | United Nations Office on Drugs and Crime |
| WHO | World Health Organization |
| PV8 | α-Pyrrolidinoheptaphenone |
| α-PVT | α-Pyrrolidinopentiothiophenone |
| LC | Liquid chromatography |
| MS | Mass spectrometry |
| MS/MS | Tandem mass spectrometry |
| HRMS | High resolution mass spectrometry |
| GC | Gas chromatography |
| FullMS | Full scan |
| ddMS2 | Data-dependent tandem mass spectrometry |
| AIF | All-ion fragmentation |
| AGC | Automatic gain control |
| IT | Injection time |
References
- Zaami, S.; Giorgetti, R.; Pichini, S.; Pantano, F.; Marinelli, E.; Busardò, F.P. Synthetic cathinones related fatalities: An update. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 268–274. [Google Scholar] [CrossRef] [PubMed]
- La Maida, N.; Di Trana, A.; Giorgetti, R.; Tagliabracci, A.; Busardò, F.P.; Huestis, M.A. A review of synthetic cathinone-related fatalities from 2017 to 2020. Ther. Drug Monit. 2020. [Google Scholar] [CrossRef] [PubMed]
- Luethi, D.; Liechti, M.E. Designer drugs: Mechanism of action and adverse effects. Arch. Toxicol. 2020, 94, 1085–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchiyama, N.; Shimokawa, Y.; Kawamura, M.; Kikura-Hanajiri, R.; Hakamatsuka, T. Chemical analysis of a benzofuran derivative, 2-(2-ethylaminopropyl)benzofuran (2-EAPB), eight synthetic cannabinoids, five cathinone derivatives, and five other designer drugs newly detected in illegal products. Forensic Toxicol. 2014, 32, 266–281. [Google Scholar] [CrossRef]
- Uchiyama, N.; Matsuda, S.; Kawamura, M.; Shimokawa, Y.; Kikura-Hanajiri, R.; Aritake, K.; Urade, Y.; Goda, Y. Characterization of four new designer drugs, 5-chloro-NNEI, NNEI indazole analog, α-PHPP and α-POP, with 11 newly distributed designer drugs in illegal products. Forensic Sci. Int. 2014, 243, 1–13. [Google Scholar] [CrossRef]
- Fels, H.; Lottner-Nau, S.; Sax, T.; Roider, G.; Graw, M.; Auwärter, V.; Musshoff, F. Postmortem concentrations of the synthetic opioid U-47700 in 26 fatalities associated with the drug. Forensic Sci. Int. 2019, 301, e20–e28. [Google Scholar] [CrossRef]
- Adamowicz, P.; Hydzik, P. Fetal death associated with the use of 3,4-MDPHP and α-PHP. Clin. Toxicol. 2019, 57, 112–116. [Google Scholar] [CrossRef]
- Freni, F.; Bianco, S.; Vignali, C.; Groppi, A.; Moretti, M.; Osculati, A.M.M.; Morini, L. A multi-analyte LC–MS/MS method for screening and quantification of 16 synthetic cathinones in hair: Application to postmortem cases. Forensic Sci. Int. 2019, 298, 115–120. [Google Scholar] [CrossRef]
- Vignali, C.; Moretti, M.; Groppi, A.; Osculati, A.M.M.; Tajana, L.; Morini, L. Distribution of the synthetic cathinone α-pyrrolidinohexiophenone in biological specimens. J. Anal. Toxicol. 2019, 43, e1–e6. [Google Scholar] [CrossRef]
- Carlier, J.; La Maida, N.; Di Trana, A.; Huestis, M.A.; Pichini, S.; Busardò, F.P. Testing unconventional matrices to monitor for prenatal exposure to heroin, cocaine, amphetamines, synthetic cathinones, and synthetic opioids. Ther. Drug Monit. 2020, 42, 205–221. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Critical Review Report: Alpha-PHP (α-Pyrrolidinohexanophenone) or PV-7; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Bonano, J.S.; Banks, M.L.; Kolanos, R.; Sakloth, F.; Barnier, M.L.; Glennon, R.A.; Cozzi, N.V.; Partilla, J.S.; Baumann, M.H.; Negus, S.S. Quantitative structure-activity relationship analysis of the pharmacology of para-substituted methcathinone analogues. Br. J. Pharmacol. 2015, 172, 2433–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suyama, J.A.; Sakloth, F.; Kolanos, R.; Glennon, R.A.; Lazenka, M.F.; Negus, S.S.; Banks, M.L. Abuse-related neurochemical effects of para-substituted methcathinone analogs in rats: Microdialysis studies of nucleus accumbens dopamine and serotonin. J. Pharmacol. Exp. Ther. 2016, 356, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshleman, A.J.; Nagarajan, S.; Wolfrum, K.M.; Reed, J.F.; Swanson, T.L.; Nilsen, A.; Janowsky, A. Structure-activity relationships of bath salt components: Substituted cathinones and benzofurans at biogenic amine transporters. Psychopharmacology 2019, 236, 939–952. [Google Scholar] [CrossRef]
- Luethi, D.; Walter, M.; Zhou, X.; Rudin, D.; Krähenbühl, S.; Liechti, M.E. Para-halogenation affects monoamine transporter inhibition properties and hepatocellular toxicity of amphetamines and methcathinones. Front. Pharmacol. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). World Health Organization Recommends 12 NPS for Scheduling; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- National Institute of Health Sciences (NIHS). Data Search System for New Psychoactive Substances. Available online: https://npsdb.nihs.go.jp (accessed on 11 November 2020).
- Paul, M.; Bleicher, S.; Guber, S.; Ippisch, J.; Polettini, A.; Schultis, W. Identification of phase i and II metabolites of the new designer drug α-pyrrolidinohexiophenone (α-PHP) in human urine by liquid chromatography quadrupole time-of-flight mass spectrometry (LC-QTOF-MS). J. Mass Spectrom. 2015, 50, 1305–1317. [Google Scholar] [CrossRef] [PubMed]
- Matsuta, S.; Shima, N.; Kamata, H.; Kakehashi, H.; Nakano, S.; Sasaki, K.; Kamata, T.; Nishioka, H.; Miki, A.; Katagi, M.; et al. Metabolism of the designer drug α-pyrrolidinobutiophenone (α-PBP) in humans: Identification and quantification of the phase I metabolites in urine. Forensic Sci. Int. 2015, 249, 181–188. [Google Scholar] [CrossRef]
- Shima, N.; Kakehashi, H.; Matsuta, S.; Kamata, H.; Nakano, S.; Sasaki, K.; Kamata, T.; Nishioka, H.; Zaitsu, K.; Sato, T.; et al. Urinary excretion and metabolism of the α-pyrrolidinophenone designer drug 1-phenyl-2-(pyrrolidin-1-yl)octan-1-one (PV9) in humans. Forensic Toxicol. 2015, 33, 279–294. [Google Scholar] [CrossRef]
- Swortwood, M.J.; Ellefsen, K.N.; Wohlfarth, A.; Diao, X.; Concheiro-Guisan, M.; Kronstrand, R.; Huestis, M.A. First metabolic profile of PV8, a novel synthetic cathinone, in human hepatocytes and urine by high-resolution mass spectrometry. Anal. Bioanal. Chem. 2016, 408, 4845–4856. [Google Scholar] [CrossRef]
- Swortwood, M.J.; Carlier, J.; Ellefsen, K.N.; Wohlfarth, A.; Diao, X.; Concheiro-Guisan, M.; Kronstrand, R.; Huestis, M.A. In vitro, in vivo and in silico metabolic profiling of α-pyrrolidinopentiothiophenone, a novel thiophene stimulant. Bioanalysis 2016, 8, 65–82. [Google Scholar] [CrossRef] [Green Version]
- Ellefsen, K.N.; Wohlfarth, A.; Swortwood, M.J.; Diao, X.; Concheiro, M.; Huestis, M.A. 4-Methoxy-α-PVP: In silico prediction, metabolic stability, and metabolite identification by human hepatocyte incubation and high-resolution mass spectrometry. Forensic Toxicol. 2016, 34, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Sauer, C.; Peters, F.T.; Haas, C.; Meyer, M.R.; Fritschi, G.; Maurera, H.H. New designer drug α-pyrrolidinovalerophenone (PVP): Studies on itsmetabolism and toxicological detection in rat urine using gas chromatographic/mass spectrometric techniques. J. Mass Spectrom. 2009, 44, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Vollmar, C.; Schwaninger, A.E.; Wolf, E.U.; Maurer, H.H. New cathinone-derived designer drugs 3-bromomethcathinone and 3-fluoromethcathinone: Studies on their metabolism in rat urine and human liver microsomes using GC-MS and LC-high-resolution MS and their detectability in urine. J. Mass Spectrom. 2012, 47, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Manier, S.K.; Richter, L.H.J.; Schäper, J.; Maurer, H.H.; Meyer, M.R. Different in vitro and in vivo tools for elucidating the human metabolism of alpha-cathinone-derived drugs of abuse. Drug Test. Anal. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wohlfarth, A.; Scheidweiler, K.B.; Pang, S.; Zhu, M.; Castaneto, M.; Kronstrand, R.; Huestis, M.A. Metabolic characterization of AH-7921, a synthetic opioid designer drug: In vitro metabolic stability assessment and metabolite identification, evaluation of in silico prediction, and in vivo confirmation. Drug Test. Anal. 2016, 8, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Diao, X.; Carlier, J.; Zhu, M.; Pang, S.; Kronstrand, R.; Scheidweiler, K.B.; Huestis, M.A. In vitro and in vivo human metabolism of a new synthetic cannabinoid NM-2201 (CBL-2201). Forensic Toxicol. 2017, 35, 20–32. [Google Scholar] [CrossRef]
- Diao, X.; Carlier, J.; Scheidweiler, K.B.; Huestis, M.A. In vitro metabolism of new synthetic cannabinoid SDB-006 in human hepatocytes by high-resolution mass spectrometry. Forensic Toxicol. 2017, 35, 252–262. [Google Scholar] [CrossRef]
- Carlier, J.; Diao, X.; Sempio, C.; Huestis, M.A. Identification of new synthetic cannabinoid ADB-CHMINACA (MAB-CHMINACA) metabolites in human hepatocytes. AAPS J. 2017, 19, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Wohlfarth, A.; Roman, M.; Andersson, M.; Kugelberg, F.C.; Diao, X.; Carlier, J.; Eriksson, C.; Wu, X.; Konradsson, P.; Josefsson, M.; et al. 25C-NBOMe and 25I-NBOMe metabolite studies in human hepatocytes, in vivo mouse and human urine with high-resolution mass spectrometry. Drug Test. Anal. 2017, 9, 680–698. [Google Scholar] [CrossRef]
- Diao, X.; Carlier, J.; Zhu, M.; Huestis, M.A. Human hepatocyte metabolism of novel synthetic cannabinoids MN-18 and its 5-fluoro analog 5F-MN-18. Clin. Chem. 2017, 63, 1753–1763. [Google Scholar] [CrossRef] [Green Version]
- Diao, X.; Carlier, J.; Zhu, M.; Huestis, M.A. Metabolism of the new synthetic cannabinoid EG-018 in human hepatocytes by high-resolution mass spectrometry. Forensic Toxicol. 2018, 36, 304–312. [Google Scholar] [CrossRef]
- Carlier, J.; Diao, X.; Huestis, M.A. Synthetic cannabinoid BB-22 (QUCHIC): Human hepatocytes metabolism with liquid chromatography-high resolution mass spectrometry detection. J. Pharm. Biomed. Anal. 2018, 157, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Matsuta, S.; Shima, N.; Kakehashi, H.; Kamata, H.; Nakano, S.; Sasaki, K.; Kamata, T.; Nishioka, H.; Miki, A.; Zaitsu, K.; et al. Metabolism of α-PHP and α-PHPP in humans and the effects of alkyl chain lengths on the metabolism of α-pyrrolidinophenone-type designer drugs. Forensic Toxicol. 2018, 36, 486–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vickers, S.; Polsky, S. The biotransformation of nitrogen containing xenobiotics to lactams. Curr. Drug Metab. 2000, 1, 357–389. [Google Scholar] [CrossRef] [PubMed]
- Shima, N.; Katagi, M.; Kamata, H.; Matsuta, S.; Sasaki, K.; Kamata, T.; Nishioka, H.; Miki, A.; Tatsuno, M.; Zaitsu, K.; et al. Metabolism of the newly encountered designer drug α- pyrrolidinovalerophenone in humans: Identification and quantitation of urinary metabolites. Forensic Toxicol. 2014, 32, 59–67. [Google Scholar] [CrossRef]
- Grapp, M.; Sauer, C.; Vidal, C.; Müller, D. GC-MS analysis of the designer drug α-pyrrolidinovalerophenone and its metabolites in urine and blood in an acute poisoning case. Forensic Sci. Int. 2016, 259, e14–e19. [Google Scholar] [CrossRef]
- Hukkanen, J.; Jacob, P.; Benowitz, N.L. Metabolism and disposition kinetics of nicotine. Pharmacol. Rev. 2005, 57, 79–115. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Jia, W.; Li, T.; Hua, Z.; Qian, Z. Identification and analytical characterization of nine synthetic cathinone derivatives N-ethylhexedrone, 4-Cl-pentedrone, 4-Cl-α-EAPP, propylone, N-ethylnorpentylone, 6-MeO-bk-MDMA, α-PiHP, 4-Cl-α-PHP, and 4-F-α-PHP. Drug Test. Anal. 2017, 9, 1162–1171. [Google Scholar] [CrossRef]
- Michaelis, W.; Russel, J.H.; Schindler, O. The metabolism of pyrovalerone hydrochloride. J. Med. Chem. 1970, 13, 497–503. [Google Scholar] [CrossRef]
- Kerrigan, S.; Savage, M.; Cavazos, C.; Bella, P. Thermal degradation of synthetic cathinones: Implications for forensic toxicology. J. Anal. Toxicol. 2016, 40, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tsujikawa, K.; Kuwayama, K.; Kanamori, T.; Iwata, Y.T.; Inoue, H. Thermal degradation of α-pyrrolidinopentiophenone during injection in gas chromatography/mass spectrometry. Forensic Sci. Int. 2013, 231, 296–299. [Google Scholar] [CrossRef]
- Fang, M.; Ivanisevic, J.; Benton, H.P.; Johnson, C.H.; Patti, G.J.; Hoang, L.T.; Uritboonthai, W.; Kurczy, M.E.; Siuzdak, G. Thermal degradation of small molecules: A global metabolomic investigation. Anal. Chem. 2015, 87, 10935–10941. [Google Scholar] [CrossRef] [PubMed]
- Carlier, J.; Diao, X.; Scheidweiler, K.B.; Huestis, M.A. Distinguishing intake of new synthetic cannabinoids ADB-PINACA and 5F-ADB-PINACA with human hepatocyte metabolites and high-resolution mass spectrometry. Clin. Chem. 2017, 63, 1008–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlier, J.; Diao, X.; Wohlfarth, A.; Scheidweiler, K.B.; Huestis, M.A. In vitro metabolite profiling of ADB-FUBINACA, a new synthetic cannabinoid. Curr. Neuropharmacol. 2017, 15, 682–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, X.; Huestis, M.A. Approaches, challenges, and advances in metabolism of new synthetic cannabinoids and identification of optimal urinary marker metabolites. Clin. Pharmacol. Ther. 2017, 101, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Huestis, M.A. New synthetic cannabinoids metabolism and strategies to best identify optimal marker metabolites. Front. Chem. 2019, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega, L.; Dinis-Oliveira, R.J. The synthetic cathinone α-pyrrolidinovalerophenone (α-PVP): Pharmacokinetic and pharmacodynamic clinical and forensic aspects. Drug Metab. Rev. 2018, 50, 125–139. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Biotransformation | [M + H]+ (m/z) | RT (min) | Mass Error (ppm) | Elemental Composition | Diagnostic Product Ions (m/z) | Peak Area at T3h | Metabolites Peak Area Fraction (%) |
|---|---|---|---|---|---|---|---|---|
| α-PHP | Parent drug | 246.1848 | 9.38 | 1.79 | C16H23NO | 70, 91, 105, 119, 140, 175 | 1.3 × 107 | |
| P1 | Hydroxylation (α-PHP 2′-hydroxypyrrolidinyl) | 262.1798 | 8.70 | 1.32 | C16H23NO2 | 70, 91, 105, 119, 156, 175 | 6.1 × 105 | 2.6% |
| P2 | Dihydroxylation (α-PHP dihydroxy-pyrrolidinyl) | 278.1746 | 9.02 | 1.72 | C16H23NO3 | 87, 91, 105, 118, 174, 186 | 1.0 × 107 | 44% |
| P3 | β-Ketoreduction (α-PHP hexanol) | 248.2005 | 9.66 | 1.63 | C16H25NO | 72, 91, 104, 117, 173, 230 | 9.7 × 106 | 40% |
| P4 | β-Ketoreduction + Ketone formation (α-PHP 2′-ketopyrrolidinyl-hexanol) | 262.1799 | 12.52 | 0.97 | C16H23NO2 | 70, 86, 91, 98, 117, 159 | 1.3 × 106 | 5.3% |
| P5 | Ketone formation (α-PHP 2′-ketopyrrolidinyl) | 260.1642 | 14.77 | 1.00 | C16H21NO2 | 86, 91, 105, 129, 133, 175 | 7.0 × 105 | 2.9% |
| 4F-α-PVP | Parent drug | 250.1598 | 8.71 | 1.33 | C15H20FNO | 70, 84, 109, 123, 126, 179 | 2.2 × 107 | |
| F1 | Hydroxylation (4F-α-PVP 2′-hydroxypyrrolidinyl) | 266.1547 | 8.12 | 1.35 | C15H20FNO2 | 70, 109, 123, 142, 179, 248 | 6.4 × 105 | 2.8% |
| F2 | Dihydroxylation (4F-α-PVP dihydroxy-pyrrolidinyl) | 282.1496 | 8.46 | 1.42 | C15H20FNO3 | 87, 109, 123, 136, 178, 179 | 9.4 × 106 | 41% |
| F3 | β-Ketoreduction (4F-α-PVP hexanol) | 252.1755 | 8.94 | 1.18 | C15H22FNO | 72, 109, 122, 190, 191, 234 | 9.1 × 106 | 40% |
| F4 | β-Ketoreduction + Ketone formation (4F-α-PVP 2′-ketopyrrolidinyl-hexanol) | 266.1548 | 11.70 | 1.01 | C15H20FNO2 | 86, 98, 109, 135, 163, 248 | 1.7 × 106 | 7.4% |
| F5 | Ketone formation (4F-α-PVP 2′-ketopyrrolidinyl) | 264.1391 | 14.00 | 1.15 | C15H18FNO2 | 98, 109, 123, 140, 161, 179 | 1.6 × 106 | 7.1% |
| α-PHP. | |||||
| FullMS/ddMS2 acquisition | |||||
| [M + H]+ (m/z) | Formula | [M + H]+ (m/z) | Formula | [M + H]+ (m/z) | Formula |
| 192.1383 | C12H17NO | 260.1645 | C16H21NO2 | 280.1907 | C16H25NO3 |
| 208.1332 | C12H17NO2 | 262.1802 | C16H23NO2 | 292.1543 | C16H21NO4 |
| 244.1696 | C16H21NO | 264.1958 | C16H25NO2 | 296.1856 | C16H25NO4 |
| 246.1852 | C16H23NO | 276.1594 | C16H21NO3 | 342.1370 | C16H23NSO5 |
| 248.2009 | C16H25NO | 278.1751 | C16H23NO3 | 438.2122 | C22H31NO8 |
| FullMS/AIF/ddMS2 acquisition | |||||
| Neutral loss (m/z) | Fragment loss | Neutral loss (m/z) | Fragment loss | Neutral loss (m/z) | Fragment loss |
| −57.0704 | −C4H9 | −127.1361 | −C8H17N | −174.1044 | −C12H14O |
| −71.0735 | −C4H9N | −141.1517 | −C9H19N | −176.1201 | −C12H16O |
| −79.9563 | −SO3 | −155.1310 | −C9H17NO | −176.0315 | −C6H8O6 |
| −106.0418 | −C7H6O | −162.1044 | −C11H14O | ||
| 4F-α-PVP | |||||
| FullMS/ddMS2 acquisition | |||||
| [M + H]+ (m/z) | Formula | [M + H]+ (m/z) | Formula | [M + H]+ (m/z) | Formula |
| 194.1176 | C11H15NO2 | 252.1758 | C15H22FNO | 282.1700 | C15H23NO4 |
| 196.1132 | C11H14FNO | 262.1438 | C15H19NO3 | 284.1657 | C15H22FNO3 |
| 212.1081 | C11H14FNO2 | 264.1394 | C15H18FNO2 | 296.1293 | C15H18FNO4 |
| 232.1696 | C15H21NO | 264.1594 | C15H21NO3 | 300.1606 | C15H22FNO4 |
| 246.1489 | C15H19NO2 | 266.1551 | C15H20FNO2 | 328.1213 | C15H21NSO5 |
| 248.1445 | C15H18FNO | 268.1707 | C15H22FNO2 | 346.1119 | C15H20FNSO5 |
| 248.1645 | C15H21NO2 | 278.1387 | C15H19NO4 | 424.1966 | C21H29NO8 |
| 250.1602 | C15H20FNO | 280.1344 | C15H18FNO3 | 442.1872 | C21H28FNO8 |
| 250.1802 | C15H23NO2 | 282.1500 | C15H20FNO3 | ||
| FullMS/AIF/ddMS2 acquisition | |||||
| Neutral loss (m/z) | Fragment loss | Neutral loss (m/z) | Fragment loss | Neutral loss (m/z) | Fragment loss |
| −43.0548 | −C3H7 | −124.0325 | −C7H5FO | −176.0315 | −C6H8O6 |
| −71.0735 | −C4H9N | −127.1361 | −C8H17N | −178.0794 | −C11H11FO |
| −79.9563 | −SO3 | −141.1154 | −C8H15NO | −180.0951 | −C11H13FO |
| −113.1205 | −C7H15N | −166.0794 | −C10H11FO | ||
| α-PHP | 4F-α-PVP | |
|---|---|---|
| Phase I expected transformations | Dehydrogenation (–2H), dihydrodiol formation (+2O +2H), oxidation (+O), oxidative deamination to alcohol (–N +O –H), oxidative deamination to ketone (–N +O –3H), reduction (+2H) | Dehydrogenation (–2H), dihydrodiol formation (+2O +2H), oxidation (+O), oxidative deamination to alcohol (–N +O –H), oxidative deamination to ketone (–N +O –3H), oxidative defluorination (–F +O +H), reduction (+2H), reductive defluorination (–F +H) |
| Phase II expected transformations | Acetylation (+2C +O +2H), glucuronidation (+6C +6O +10H), sulfation (+S +3O) | Acetylation (+2C +O +2H), glucuronidation (+6C +6O +10H), sulfation (+S +3O) |
| Maximum number of dealkylation steps | 2 | 2 |
| Maximum number of phase II reactions | 1 | 1 |
| Maximum number of reactions | 5 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlier, J.; Diao, X.; Giorgetti, R.; Busardò, F.P.; Huestis, M.A. Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations. Int. J. Mol. Sci. 2021, 22, 230. https://doi.org/10.3390/ijms22010230
Carlier J, Diao X, Giorgetti R, Busardò FP, Huestis MA. Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations. International Journal of Molecular Sciences. 2021; 22(1):230. https://doi.org/10.3390/ijms22010230
Chicago/Turabian StyleCarlier, Jeremy, Xingxing Diao, Raffaele Giorgetti, Francesco P. Busardò, and Marilyn A. Huestis. 2021. "Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations" International Journal of Molecular Sciences 22, no. 1: 230. https://doi.org/10.3390/ijms22010230
APA StyleCarlier, J., Diao, X., Giorgetti, R., Busardò, F. P., & Huestis, M. A. (2021). Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations. International Journal of Molecular Sciences, 22(1), 230. https://doi.org/10.3390/ijms22010230