Promising Antioxidant Activity of Erythrina Genus: An Alternative Treatment for Inflammatory Pain?

 , ,
, ,  , ,
, ,
Abstract
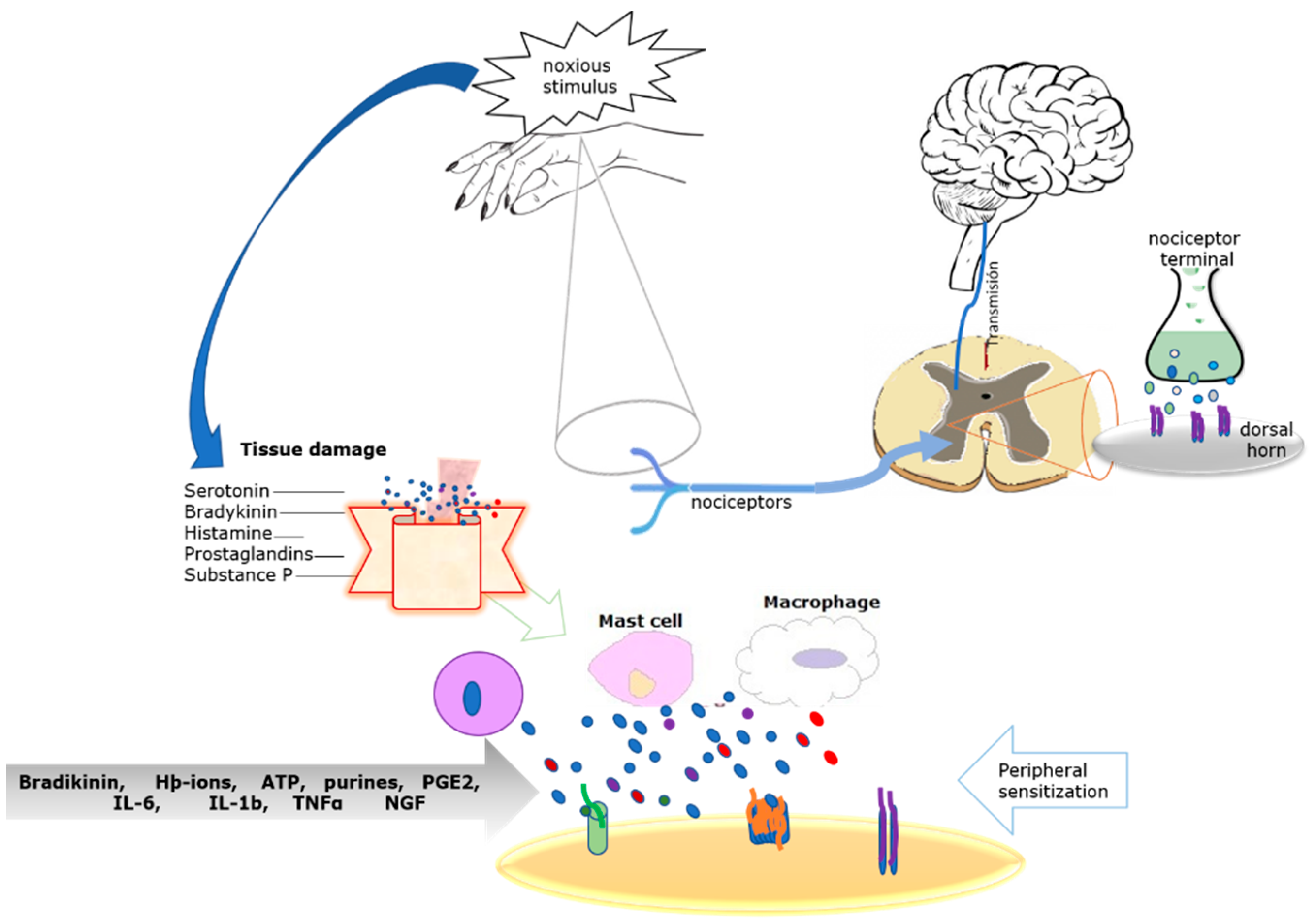
:1. Molecular Origin of Inflammatory Pain
2. Ethnomedicinal Use of the Genus Erythrina
3. Preclinical Studies of Pain and Inflammation from the Genus Erythrina
4. Radical Scavenging Activity in the Model In Vitro Systems
5. Molecular Mechanisms of Anti-Inflammatory Activity of the Genus Erythrina
6. Methods
6.1. Search Strategy
6.2. Inclusion Criteria
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| MAPK | Mitogen-Activated Protein Kinase |
| AP1 | Activator protein 1 |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| ERK | Extracellular signal-regulated kinases |
| JNK | Jun N-Terminal Kinase |
| JAK | Janus kinases |
| STAT | Signal Transducer and Activator of Transcription |
| IASP | International Association for the Study of pain |
| NSAIDs | Nonsteroidal anti-inflammatory drugs |
| CNS | Central Nervous System |
| NO | Nitric Oxide |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| BHA | Butylated hydroxyanisole |
| NAChRs | Nicotinic Acetylcholine Receptors |
| COX-2 | Cyclooxygenase 2 |
| iNOs | Nitric oxide synthase, inducible |
| LOX | Lipoxygenases |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| TNFα | Tumor Necrosis Factor alpha |
| IL-1β | Interleukin-1-beta |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| HO-1 | Heme Oxygenase-1 |
| GCLC | γ-glutamyl cysteine synthetase catalytic subunit |
| NAD(P)H | Nicotinamide adenine dinucleotide phosphate |
| GSH | Glutathione |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| ELSE | E. speciosa methanol extract |
| TRPV1 | Transient receptor potential cation channel subfamily V member 1 |
| RMV | Rostral ventrolateral medulla |
| XO | Xanthine oxidase |
| XOI | Xanthine oxidase inhibitor |
| BDE | Second bond dissociation enthalpy |
| EDG | Group Electrondonators |
References
- Hudspith, M.J. Anatomy, physiology and pharmacology of pain. Anaesth. Intensive Care Med. 2016, 17, 425–430. [Google Scholar] [CrossRef]
- Bentley, N.; Awad, A.J.; Patil, P.G. Physiology and Pathophysiology of Chronic Pain. Neuromodulation 2018, 565–573. [Google Scholar] [CrossRef]
- Ellison, D.L. Physiology of Pain. Crit. Care Nurs. Clin. N. Am. 2017, 29, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, D.A.; Hobelmann, J.G.; Compton, P. Providing chronic pain management in the “Fifth Vital Sign” Era: Historical and treatment perspectives on a modern-day medical dilemma. Drug Alcohol Depend. 2017, 173, S11–S21. [Google Scholar] [CrossRef] [PubMed]
- Kakavandi, N.; Rezaee, S.; Hosseini-Fard, S.R.; Ghasempour, G.; Khosravi, M.; Shabani, M.; Najafi, M. Prostaglandin E2 (PGE2) synthesis pathway is involved in coronary artery stenosis and restenosis. Gene 2021, 765, 145131. [Google Scholar] [CrossRef]
- Kanda, H.; Kobayashi, K.; Yamanaka, H.; Okubo, M.; Dai, Y.; Noguchi, K. Localization of prostaglandin E2 synthases and E-prostanoid receptors in the spinal cord in a rat model of neuropathic pain. Brain Res. 2021, 1750, 147153. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Chaudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative stress and antioxidants in disease and cancer: A review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, C.; Naziroglu, M.; Rodríguez, A.B.; Pariente, J.A. Neuropathic pain: Delving into the oxidative origin and the possible implication of transient receptor potential channels. Front. Physiol. 2018, 9, 95. [Google Scholar] [CrossRef]
- Dahlhamer, J.; Lucas, J.; Zelaya, C.; Nahin, R.; Mackey, S.; DeBar, L.; Kerns, R.; von Korff, M.; Porter, L.; Helmick, C. Prevalence of Chronic Pain and High-Impact Chronic Pain Among Adults—United States, 2016. Mmwr. Morb. Mortal. Wkly. Rep. 2018, 67, 1001–1006. [Google Scholar] [CrossRef]
- Muley, M.M.; Krustev, E.; Mcdougall, J.J. Preclinical Assessment of Inflammatory Pain. CNS Neurosci. Ther. 2016, 22, 88–101. [Google Scholar] [CrossRef]
- Vargas, J.R.N.; Pinzón, J.E.C. The labyrinth of pain and the need for fostering basic research: El laberinto del dolor y la necesidad de impulsar la investigación básica. Colomb. J. Anesthesiol. 2012, 41, 1–3. [Google Scholar] [CrossRef]
- Robinson, M.M.; Zhang, X. The World Medicines Situation 2011 Traditional Medicines: Global Situation, Issues and Challenges; World Health Organization: Geneva, Switzerland, 2011; pp. 1–14. [Google Scholar]
- Benvenuti, S.; Bortolotti, E.; Maggini, R. Antioxidant power, anthocyanin content and organoleptic performance of edible flowers. Sci. Hortic. 2016, 199, 170–177. [Google Scholar] [CrossRef]
- Li, S.H.; Li, L.; Yang, R.N.; Liang, S.D. Compounds of traditional Chinese medicine and neuropathic pain. Chin. J. Nat. Med. 2020, 18, 28–35. [Google Scholar] [CrossRef]
- Lee, F.H.; Raja, S.N. Complementary and alternative medicine in chronic pain. Pain 2011, 152, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Faggion, S.A.; Cunha, A.O.S.; Fachim, H.A.; Gavin, A.S.; dos Santos, W.F.; Pereira, A.M.S.; Beleboni, R.O. Anticonvulsant profile of the alkaloids (+)-erythravine and (+)-11-α-hydroxy-erythravine isolated from the flowers of Erythrina mulungu Mart ex Benth (Leguminosae-Papilionaceae). Epilepsy Behav. 2011, 20, 441–446. [Google Scholar] [CrossRef]
- Santos Rosa, D.; Faggion, S.A.; Gavin, A.S.; Anderson de Souza, M.; Fachim, H.A.; Ferreira dos Santos, W.; Soares Pereira, A.M.; Cunha, A.O.S.; Beleboni, R.O. Erysothrine, an alkaloid extracted from flowers of Erythrina mulungu Mart. ex Benth: Evaluating its anticonvulsant and anxiolytic potential. Epilepsy Behav. 2012, 23, 205–212. [Google Scholar] [CrossRef]
- Akter, K.; Barnes, E.C.; Loa-Kum-Cheung, W.L.; Yin, P.; Kichu, M.; Brophy, J.J.; Barrow, R.A.; Imchen, I.; Vemulpad, S.R.; Jamie, J.F. Antimicrobial and antioxidant activity and chemical characterisation of Erythrina stricta Roxb. (Fabaceae). J. Ethnopharmacol. 2016, 185, 171–181. [Google Scholar] [CrossRef]
- Amorim, J.; Borges, M.; de Carvalho Borges, M.; Fabro, A.T.; Contini, S.H.T.; Valdevite, M.; Pereira, A.M.S.; Carmona, F. The ethanolic extract from Erythrina mulungu Benth. flowers attenuates allergic airway inflammation and hyperresponsiveness in a murine model of asthma. J. Ethnopharmacol. 2019, 242. [Google Scholar] [CrossRef]
- Mukungu, N.; Abuga, K.; Okalebo, F.; Ingwela, R.; Mwangi, J. Medicinal plants used for management of malaria among the Luhya community of Kakamega East sub-County, Kenya. J. Ethnopharmacol. 2016, 194, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Bodofsky, S.; Merriman, T.R.; Thomas, T.J.; Schlesinger, N. Advances in our understanding of gout as an auto-inflammatory disease. Semin. Arthritis Rheum. 2020, 50, 1089–1100. [Google Scholar] [CrossRef]
- Togola, A.; Austarheim, I.; Theïs, A.; Diallo, D.; Paulsen, B. Ethnopharmacological uses of Erythrina senegalensis: A comparison of three areas in Mali, and a link between traditional knowledge and modern biological science. J. Ethnobiol. Ethnomedicine 2008, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luca, A.; Sibilio, G.; de Luca, P.; del Guacchio, E. DNA barcoding to confirm the morphological identification of the coral trees (Erythrina spp., fabaceae) in the ancient gardens of Naples (Campania, Italy). Plants 2018, 7, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Lingadurai, S.; Jain, A.; Barman, N. Erythrina variegata Linn: A review on morphology, phytochemistry, and pharmacological aspects. Pharmacogn. Rev. 2010, 4, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlage, C.; Mabula, C.; Mahunnah, R.L.A.; Heinrich, M. Medicinal plants of the Washambaa (Tanzania): Documentation and ethnopharmacological evaluation. Plant Biol. 2000, 2, 83–92. [Google Scholar] [CrossRef]
- Corrigan, B.M.; van Wyk, B.E.; Geldenhuys, C.J.; Jardine, J.M. Ethnobotanical plant uses in the KwaNibela Peninsula, St Lucia, South Africa. S. Afr. J. Bot. 2011, 77, 346–359. [Google Scholar] [CrossRef]
- Pompermaier, L.; Marzocco, S.; Adesso, S.; Monizi, M.; Schwaiger, S.; Neinhuis, C.; Stuppner, H.; Lautenschläger, T. Medicinal plants of northern Angola and their anti-inflammatory properties. J. Ethnopharmacol. 2018, 216, 26–36. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; McGaw, L.J.; Eloff, J.N. In vitro antibacterial, antioxidant and cytotoxic activity of acetone leaf extracts of nine under-investigated Fabaceae tree species leads to potentially useful extracts in animal health and productivity. BMC Complement. Altern. Med. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Mekky, H.; El-Masry, S. Metabolomics driven analysis of Erythrina lysistemon cell suspension culture in response to methyl jasmonate elicitation Erythrina lysistemon cell culture metabolomics. J. Adv. Res. 2016, 7, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Mhlongo, L.S.; van Wyk, B.E. Zulu medicinal ethnobotany: New records from the Amandawe area of KwaZulu-Natal, South Africa. S. Afr. J. Bot. 2019, 122, 266–290. [Google Scholar] [CrossRef]
- Wangchuk, P.; Yeshi, K.; Jamphel, K. Pharmacological, ethnopharmacological, and botanical evaluation of subtropical medicinal plants of Lower Kheng region in Bhutan. Integr. Med. Res. 2017, 6, 372–387. [Google Scholar] [CrossRef]
- Lee, S.; Xiao, C.; Pei, S. Ethnobotanical survey of medicinal plants at periodic markets of Honghe Prefecture in Yunnan Province, SW China. J. Ethnopharmacol. 2008, 117, 362–377. [Google Scholar] [CrossRef]
- Maroyi, A. An ethnobotanical survey of medicinal plants used by the people in Nhema communal area, Zimbabwe. J. Ethnopharmacol. 2011, 136, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Tene, V.; Malagón, O.; Finzi, P.V.; Vidari, G.; Armijos, C.; Zaragoza, T. An ethnobotanical survey of medicinal plants used in Loja and Zamora-Chinchipe, Ecuador. J. Ethnopharmacol. 2007, 111, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Kamble, M.Y.; Mane, S.S.; Murugan, C.; Jaisankar, I. Diversity of Ethno-Medicinal Plants of Tropical Islands-with Special Reference to Andaman and Nicobar Islands; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128130643. [Google Scholar]
- Bhagyasri, Y.; Nagalatha, G.; Reddy, N.V.; Subramanian, N.S. Analgesic and anti-inflammatory activity of leaf extracts of Erythrina variegate. Indo Am. J. Pharm. Res. 2017, 7, 681–692. [Google Scholar]
- Fahmy, N.M.; Al-Sayed, E.; El-Shazly, M.; Singab, A.N. Comprehensive review on flavonoids biological activities of Erythrina plant species. Ind. Crop. Prod. 2018, 123, 500–538. [Google Scholar] [CrossRef]
- Haque, R.; Ali, M.S.; Saha, A.; Alimuzzaman, M. Analgesic Activity of Methanolic Extract of the Leaf of Erythrina variegata. Dhaka Univ. J. Pharm. Sci. 2006, 5, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Nasir Uddin, M.M.; Emran, T.B.; Mahib, M.; Dash, R. Molecular docking and analgesic studies of Erythrina variegata’s derived phytochemicals with COX enzymes. Bioinformation 2014, 10, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Krishna Raju Mantena, V.R.; Tejaswini, G. Anti inflammatory activity of Erythrina Variegata. Int. J. Pharm. Pharm. Sci. 2015, 7, 386–388. [Google Scholar]
- Vasconcelos, S.M.M.; Rebouças Oliveira, G.; Mohana De Carvalho, M.; Rodrigues, A.C.P.; Rocha Silveira, E.; Maria França Fonteles, M.; Florenço Sousa, F.C.; Barros Viana, G.S. Antinociceptive activities of the hydroalcoholic extracts from Erythrina velutina and Erythrina mulungu in mice. Biol. Pharm. Bull. 2003, 26, 946–949. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, S.M.M.; Macedo, D.S.; de Melo, C.T.v.; Monteiro, A.P.; Cunha, G.M.A.; Sousa, F.C.F.; Viana, G.S.B.; Rodrigues, A.C.P.; Silveira, E.R. Central activity of hydroalcoholic extracts from Erythrina velutina and Erythrina mulungu in mice. J. Pharm. Pharmacol. 2004, 56, 389–393. [Google Scholar] [CrossRef]
- Marchioro, M.; Blank, M.D.F.A.; Mourão, R.H.V.; Antoniolli, Â.R. Anti-nociceptive activity of the aqueous extract of Erythrina velutina leaves. Fitoterapia 2005, 76, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Thuong, P.T.; Lee, H.S.; Ndinteh, D.T.; Mbafor, J.T.; Fomum, Z.T.; Oh, W.K. Flavanones from the stem bark of Erythrina abyssinica. Bioorg. Med. Chem. 2008, 16, 10356–10362. [Google Scholar] [CrossRef]
- Sokeng, S.D.; Talla, E.; Jeweldai, V.; Yaya, A.J.G.; Koube, J.; Dongmo, F.; Goulimé, M.; Mbafor, J.T. Anti-inflammatory effect of Abyssinone V-4′-methyl ether on acute and chronic inflammation models. Hygeia J. Drugs Med. 2013, 5, 121–128. [Google Scholar]
- Talla, E.; Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Kamanyi, A.; Mbanya, J.C.; Giner, R.M.; Recio, M.C.; Máñez, S.; Ríos, J.L. Warangalone, the isoflavonoid anti-inflammatory principle of Erythrina addisoniae stem bark. J. Nat. Prod. 2003, 66, 891–893. [Google Scholar] [CrossRef] [PubMed]
- Branco, A.C.C.C.; Yoshikawa, F.S.Y.; Pietrobon, A.J.; Sato, M.N. Role of Histamine in Modulating the Immune Response and Inflammation. Mediat. Inflamm. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Kamanyi, A.; Mbanya, J.C.; Recio, M.C.; Giner, R.M.; Máñez, S.; Ríos, J.L. Anti-inflammatory activities of two flavanones, sigmoidin A and sigmoidin B, from Erythrina sigmoidea. Planta Med. 2004, 70, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Pillay, C.C.N.; Jäger, A.K.; Mulholland, D.A.; van Staden, J. Cyclooxygenase inhibiting and anti-bacterial activities of South African Erythrina species. J. Ethnopharmacol. 2001, 74, 231–237. [Google Scholar] [CrossRef]
- Yenesew, A.; Twinomuhwezi, H.; Kiremire, B.T.; Mbugua, M.N.; Gitu, P.M.; Heydenreich, M.; Peter, M.G. 8-Methoxyneorautenol and radical scavenging flavonoids from Erythrina abyssinica. Bull. Chem. Soc. Ethiop. 2009, 23, 205–210. [Google Scholar] [CrossRef]
- MacHumi, F.; Bojase-Moleta, G.; Mapitse, R.; Masesane, I.; Majinda, R.R.T. Radical scavenging-flavonoids from Erythrina abyssinica. Nat. Prod. Commun. 2006, 1, 287–292. [Google Scholar] [CrossRef]
- Mujahid, M.; Hussain, T.; Siddiqui, H.H.; Hussain, A. Evaluation of hepatoprotective potential of Erythrina indica leaves against antitubercular drugs induced hepatotoxicity in experimental rats. J. Ayurveda Integr. Med. 2017, 8, 7–12. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Joseph, J.M.; Rajendrakumaran, D. In vitro xanthine oxidase inhibitory activity of methanol extracts of Erythrina indica Lam. leaves and stem bark. Asian Pac. J. Trop. Biomed. 2012, 2, S1415–S1417. [Google Scholar] [CrossRef]
- Sre, P.R.R.; Sheila, T.; Murugesan, K. Phytochemical screening and “in-vitro” anti-oxidant activity of methanolic root extract of Erythrina indica. Asian Pac. J. Trop. Biomed. 2012, 2, 2–6. [Google Scholar] [CrossRef]
- Anouar, E.H. Antioxidant activity of mildbone and mildbenone secondary metabolites of Erythrina mildbraedii Harms: A theoretical approach. Comput. Theor. Chem. 2016, 1077, 106–112. [Google Scholar] [CrossRef]
- Ali, M.S.; Ali, M.I.; Ahmed, G.; Afza, N.; Lateef, M.; Iqbal, L.; Waffo, A.F.K.; Ahmed, Z. Potent antioxidant and lipoxygenase inhibitory flavanone and chalcone from Erythrina mildbraedii harms (Fabaceae) of cameroon. Z. Fur Nat.-Sect. B J. Chem. Sci. 2012, 67, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Njamen, D.; Talla, E.; Mbafor, J.T.; Fomum, Z.T.; Kamanyi, A.; Mbanya, J.C.; Cerdá-Nicolás, M.; Giner, R.M.; Recio, M.C.; Ríos, J.L. Anti-inflammatory activity of erycristagallin, a pterocarpene from Erythrina mildbraedii. Eur. J. Pharmacol. 2003, 468, 67–74. [Google Scholar] [CrossRef]
- Gabr, S.K.; Bakr, R.O.; Mostafa, E.S.; El-Fishawy, A.M.; El-Alfy, T.S. Antioxidant activity and molecular docking study of Erythrina × neillii polyphenolics. S. Afr. J. Bot. 2019, 121, 470–477. [Google Scholar] [CrossRef]
- Verma, S.M.; Prakash, J.; Sah, V.K. Phyto-Pharmacognostical Investigation and Evaluation of Anti-Inflammatory and Sedative Hypnotic Activity of the Leaves of Erythrina Indica Lam. Anc. Sci. Life 2005, 25, 79–83. [Google Scholar]
- Fischer, L.G.O.; Leitão, R.; Etcheverry, S.R.; de Campos-Buzzi, F.; Vãzquez, A.A.; Heinzen, H.A.; Filho, V.C. Analgesic properties of extracts and fractions from Erythrina crista-galli (Fabaceae) leaves. Nat. Prod. Res. 2007, 21, 759–766. [Google Scholar] [CrossRef]
- Vasconcelos, S.M.M.; Sales, G.T.M.; Lima, N.; Lobato, R.d.F.G.; Macêdo, D.S.; Barbosa-Filho, J.M.; Leal, L.K.A.M.; Fonteles, M.M.F.; Sousa, F.C.F.; Oliveira, J.L.; et al. Anti-inflammatory activities of the hydroalcoholic extracts from Erythrina velutina and E. Mulungu in mice. Braz. J. Pharmacogn. 2011, 21, 1155–1158. [Google Scholar] [CrossRef] [Green Version]
- Musa, A.; Nazifi, A.B.; Usman, A.I.; Kassim, A. Evaluation of analgesic and behavioural effects of ethanol root bark extract of Erythrina senegalensis DC ( Fabaceae ). UoN Protal 2016, 5, 81–86. [Google Scholar]
- Saidu, K.; Onah, J.; Orisadipe, A.; Olusola, A.; Wambebe, C.; Gamaniel, K. Antiplasmodial, analgesic, and anti-inflammatory activities of the aqueous extract of the stem bark of Erythrina senegalensis. J. Ethnopharmacol. 2000, 71, 275–280. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedane, K.G.; Kusari, S.; Masesane, I.B.; Spiteller, M.; Majinda, R.R.T. Flavanones of Erythrina livingstoniana with antioxidant properties. Fitoterapia 2016, 108, 48–54. [Google Scholar] [CrossRef]
- Bedane, K.G.; Kusari, S.; Eckelmann, D.; Masesane, I.B.; Spiteller, M.; Majinda, R.R.T. Erylivingstone A-C with antioxidant and antibacterial activities from Erythrina livingstoniana. Fitoterapia 2015, 105, 113–118. [Google Scholar] [CrossRef]
- Tauseef, S.; Ali, M.S.; Ahmed, A.; Ali, M.I.; Ahmed, Z.; Sherwani, S.K.; Ahmed, G.; Onocha, P.A.; Joseph, N.; Francois, A.; et al. In vitro Antioxidant activity analysis of five medicinally important plants. J. Pharmacogn. Phytochem. 2013, 2, 183–188. [Google Scholar]
- Zhang, C.; Zhou, Y.; Sun, Z.; Feng, J.; Wang, Y. Polysaccharides extraction from Erythirna variegata, chemical characterization and its antioxidant activity. Int. J. Biol. Macromol. 2014, 68, 267–273. [Google Scholar] [CrossRef]
- Janbaz, K.H.; Nisar, U.; Ashraf, M.; Qadir, M.I. Spasmolytic, bronchodilatory and antioxidant activities of Erythrina superosa Roxb. Acta Pol. Pharm.-Drug Res. 2012, 69, 1111–1117. [Google Scholar]
- Umamaheswari, M.; Asokkumar, K.; Sivashanmugam, A.T.; Remyaraju, A.; Subhadradevi, V.; Ravi, T.K. In vitro xanthine oxidase inhibitory activity of the fractions of Erythrina stricta Roxb. J. Ethnopharmacol. 2009, 124, 646–648. [Google Scholar] [CrossRef]
- Bakr, R.O.; Fayed, M.A.A.; Fayez, A.M.; Gabr, S.K.; El-Fishawy, A.M.; Taha, S.E.A. Hepatoprotective activity of Erythrina × neillii leaf extract and characterization of its phytoconstituents. Phytomedicine 2019, 53, 9–17. [Google Scholar] [CrossRef]
- Ozawa, M.; Kawamata, S.; Etoh, T.; Hayashi, M.; Komiyama, K.; Kishida, A.; Kuroda, C.; Ohsaki, A. Structures of new erythrinan alkaloids and nitric oxide production inhibitors from Erythrina crista-galli. Chem. Pharm. Bull. 2010, 58, 1119–1122. [Google Scholar] [CrossRef] [Green Version]
- Gurmessa, G.T.; Kusari, S.; Laatsch, H.; Bojase, G.; Tatolo, G.; Masesane, I.B.; Spiteller, M.; Majinda, R.R.T. Chemical constituents of root and stem bark of Erythrina brucei. Phytochem. Lett. 2018, 25, 37–42. [Google Scholar] [CrossRef]
- Yenesew, A.; Irungu, B.; Derese, S.; Midiwo, J.O.; Heydenreich, M.; Peter, M.G. Two prenylated flavonoids from the stem bark of Erythrina burttii. Phytochemistry 2003, 63, 445–448. [Google Scholar] [CrossRef]
- Yenesew, A.; Midiwo, J.O.; Heydenreich, M.; Schanzenbach, D.; Peter, M.G. Two isoflavanones from the stem bark of Erythrina sacleuxii. Phytochemistry 2000, 55, 457–459. [Google Scholar] [CrossRef]
- Koch, K.; Schulz, G.; Döring, W.; Büchter, C.; Havermann, S.; Mutiso, P.C.; Passreiter, C.; Wätjen, W. Abyssinone V, a prenylated flavonoid isolated from the stem bark of Erythrina melanacantha increases oxidative stress and decreases stress resistance in Caenorhabditis elegans. J. Pharm. Pharmacol. 2019, 71, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Yenesew, A.; Akala, H.M.; Twinomuhwezi, H.; Chepkirui, C.; Irungu, B.N.; Eyase, F.L.; Kamatenesi-Mugisha, M.; Kiremire, B.T.; Johnson, J.D.; Waters, N.C. The antiplasmodial and radical scavenging activities of flavonoids of Erythrina burttii. Acta Trop. 2012, 123, 123–127. [Google Scholar] [CrossRef]
- Bedane, K.G.; Kusari, S.; Bullach, A.; Masesane, I.B.; Mihigo, S.O.; Spiteller, M.; Majinda, R.R.T. Chemical constituents of the root bark of Erythrina droogmansiana. Phytochem. Lett. 2017, 20, 84–88. [Google Scholar] [CrossRef]
- Yaya, A.J.G.; Feumba, R.D.; Emmanuel, T.; Tchinda, A.T.; Fredérich, M.; Oben, J.; Mbafor, J.T. Antioxidant activity of compounds isolated from the root woods of Erythrina droogmansiana. Int. J. Pharm. Sci. Drug Res. 2014, 6, 160–163. [Google Scholar]
- Intiquilla, A.; Jimenez, K.; Zavaleta, A.I. Erythrina edulis (Pajuro) Seed Protein: A New Source of Antioxidant Peptides. Natural Product Communications 2016, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bedane, K.G.; Masesane, I.B.; Majinda, R.R.T. New isoflavans from the root bark of Erythrina livingstoniana. Phytochem. Lett. 2016, 17, 55–58. [Google Scholar] [CrossRef]
- Alam, M.K.; Rana, Z.H.; Islam, S.N.; Akhtaruzzaman, M. Comparative assessment of nutritional composition, polyphenol profile, antidiabetic and antioxidative properties of selected edible wild plant species of Bangladesh. Food Chem. 2020, 320, 126646. [Google Scholar] [CrossRef]
- Jang, J.P.; Na, M.K.; Thuong, P.T.; Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Woo, E.R.; Oh, W.K. Prenylated flavonoids with PTP1B inhibitory activity from the root bark of Erythrina mildbraedii. Chem. Pharm. Bull. 2008, 56, 85–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donfack, J.H.; Nico, F.N.; Ngameni, B.; Tchana, A.; Chuisseu, P.D.; Finzi, P.V.; Ngadjui, B.T.; Moundipa, P.F. In vitro hepatoprotective and antioxidant activities of diprenylated isoflavonoids from Erythrina senegalensis (Fabaceae). Pharmacologyonline 2008, 1, 382–391. [Google Scholar]
- Yahaya, E.S.; Cordier, W.; Steenkamp, P.A.; Steenkamp, V. Protective effect of Erythrina senegalensis sequential extracts against oxidative stress in SC-1 fibroblasts and THP-1 macrophages. J. Pharm. Pharmacogn. Res. 2020, 8, 247–259. [Google Scholar]
- Leonti, M.; Stafford, G.I.; Cero, M.D.; Cabras, S.; Castellanos, M.E.; Casu, L.; Weckerle, C.S. Reverse ethnopharmacology and drug discovery. J. Ethnopharmacol. 2017, 198, 417–431. [Google Scholar] [CrossRef]
- Anywar, G.; Namukobe, J. Factors Affecting the Choice for Plant-Based Products in Drug Discoveries; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128178904. [Google Scholar]
- Fahmy, N.M.; Al-Sayed, E.; El-Shazly, M.; Nasser Singab, A. Alkaloids of genus Erythrina: An updated review. Nat. Prod. Res. 2020, 34, 1891–1912. [Google Scholar] [CrossRef] [PubMed]
- Feehan, K.T.; Gilroy, D.W. Is Resolution the End of Inflammation? Trends Mol. Med. 2019, 25, 198–214. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299. [Google Scholar] [CrossRef]
- Elgorashi, E.E.; McGaw, L.J. African plants with in vitro anti-inflammatory activities: A review. S. Afr. J. Bot. 2019, 126, 142–169. [Google Scholar] [CrossRef]
- Aswad, M.; Rayan, M.; Abu-Lafi, S.; Falah, M.; Raiyn, J.; Abdallah, Z.; Rayan, A. Nature is the best source of anti-inflammatory drugs: Indexing natural products for their anti-inflammatory bioactivity. Inflamm. Res. 2018, 67, 67–75. [Google Scholar] [CrossRef]
- Amaral, P.d.A.; Antunes, A.R.; Barlow, J.W. Isolation of erythrinan alkaloids from the leaves and flowers of Erythrina speciosa. Rev. Bras. De Farmacogn. 2019, 29, 488–490. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, B.J.; Bao, M.F.; Cai, X.H. A new erythrinan N-oxide alkaloid from Erythrina stricta. Nat. Prod. Res. 2019, 33, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Crestey, F.; Jensen, A.A.; Borch, M.; Andreasen, J.T.; Andersen, J.; Balle, T.; Kristensen, J.L. Design, synthesis, and biological evaluation of Erythrina alkaloid analogues as neuronal nicotinic acetylcholine receptor antagonists. J. Med. Chem. 2013, 56, 9673–9682. [Google Scholar] [CrossRef] [PubMed]
- Umana, I.C.; Daniele, C.A.; McGehee, D.S. Neuronal nicotinic receptors as analgesic targets: It’s a winding road. Biochem. Pharmacol. 2013, 86, 1208–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.X.; Hierl, M.; Clarkin, K.; Juan, T.; Nguyen, H.; Valk, M.v.d.; Deng, H.; Guo, W.; Lehto, S.G.; Matson, D.; et al. Pharmacological effects of nonselective and subtype-selective nicotinic acetylcholine receptor agonists in animal models of persistent pain. Pain 2010, 149, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Egea, J.; Buendia, I.; Parada, E.; Navarro, E.; León, R.; Lopez, M.G. Anti-inflammatory role of microglial alpha7 nAChRs and its role in neuroprotection. Biochem. Pharmacol. 2015, 97, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, Y.; Kagan, S.; Cohen Ben-Ami, H.; Rostami, J.; Mizrahi, T.; Kulkarni, A.R.; Thakur, G.A.; Vaknin-Dembinsky, A.; Healy, L.M.; Brenner, T.; et al. RIC3, the cholinergic anti-inflammatory pathway, and neuroinflammation. Int. Immunopharmacol. 2020, 83, 106381. [Google Scholar] [CrossRef]
- Pohanka, M. Alpha7 nicotinic acetylcholine receptor is a target in pharmacology and toxicology. Int. J. Mol. Sci. 2012, 13, 2219–2238. [Google Scholar] [CrossRef] [Green Version]
- Setti-Perdigão, P.; Serrano, M.A.R.; Flausino, O.A.; Bolzani, V.S.; Guimarães, M.Z.P.; Castro, N.G. Erythrina mulungu alkaloids are potent inhibitors of neuronal nicotinic receptor currents in mammalian cells. PLoS ONE 2013, 8, e82726. [Google Scholar] [CrossRef]
- Sun, H.; Cai, W.; Wang, X.; Liu, Y.; Hou, B.; Zhu, X.; Qiu, L. Vaccaria hypaphorine alleviates lipopolysaccharide-induced inflammation via inactivation of NFΚB and ERK pathways in Raw 264.7 cells. BMC Complement. Altern. Med. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- El-masry, S.; Hammoda, H.M.; Zaatout, H.; Abdel-kader, M. Constituents of Erythrina caffra Stem Bark Grown in Egypt. Nat. Prod. Sci. 2010, 16, 210–216. [Google Scholar]
- Ozawa, M.; Honda, K.; Nakai, I.; Kishida, A.; Ohsaki, A. Hypaphorine, an indole alkaloid from Erythrina velutina, induced sleep on normal mice. Bioorg. Med. Chem. Lett. 2008, 18, 3992–3994. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharmacother. 2020, 129. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Ansari, M.Y.; Bano, S.; Haqqi, T.M. Imperatorin suppresses IL-1β-induced iNOS expression via inhibiting ERK-MAPK/AP1 signaling in primary human OA chondrocytes. Int. Immunopharmacol. 2020, 85, 106612. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Sureda, A.; Belwal, T.; Çetinkaya, S.; Süntar, İ.; Tejada, S.; Devkota, H.P.; Ullah, H.; Aschner, M. Polyphenols in the treatment of autoimmune diseases. Autoimmun. Rev. 2019, 18, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.K.; Choi, J.; Jang, H.; Seol, J.W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef] [PubMed]
- Afroz, N.; Ahsanul Hoq, M.; Jahan, S.; Mainul Islam, M.; Ahmed, F.; Shahid-Ud-Daula, A.F.M.; Hasanuzzaman, M. Methanol soluble fraction of fruits of Annona muricata possesses significant antidiarrheal activities. Heliyon 2020, 6, e03112. [Google Scholar] [CrossRef] [Green Version]
- Fahmy, N.M.; Al-Sayed, E.; Michel, H.E.; El-Shazly, M.; Singab, A.N.B. Gastroprotective effects of Erythrina speciosa (Fabaceae) leaves cultivated in Egypt against ethanol-induced gastric ulcer in rats. J. Ethnopharmacol. 2020, 248, 112297. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.L.; Tran, P.T.; Tran, H.N.K.; Lee, S.; Kim, O.; Min, B.S.; Lee, J.H. A prenylated flavonoid, 10-oxomornigrol F, exhibits anti-inflammatory effects by activating the Nrf2/heme oxygenase-1 pathway in macrophage cells. Int. Immunopharmacol. 2018, 55, 165–173. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Nachtergael, A.; Nguyen, T.M.; Cornet, V.; Duez, P.; Muller, M.; Ly Huong, D.T.; Kestemont, P. Anti-inflammatory properties of the ethanol extract from Clerodendrum cyrtophyllum Turcz based on in vitro and in vivo studies. J. Ethnopharmacol. 2020, 254, 112739. [Google Scholar] [CrossRef]
- Li, X.; Qin, X.; Tian, J.; Gao, X.; Wu, X.; Du, G.; Zhou, Y. Liquiritin protects PC12 cells from corticosterone-induced neurotoxicity via regulation of metabolic disorders, attenuation ERK1/2-NF-κB pathway, activation Nrf2-Keap1 pathway, and inhibition mitochondrial apoptosis pathway. Food Chem. Toxicol. 2020, 146, 111801. [Google Scholar] [CrossRef]
- Gandhi, M.; Nair, S. New vistas in malignant mesothelioma: MicroRNA architecture and NRF2/MAPK signal transduction. Life Sci. 2020, 257, 118123. [Google Scholar] [CrossRef]
- Zhao, J.; Piao, X.; Wu, Y.; Liang, S.; Han, F.; Liang, Q.; Shao, S.; Zhao, D. Cepharanthine attenuates cerebral ischemia/reperfusion injury by reducing NLRP3 inflammasome-induced inflammation and oxidative stress via inhibiting 12/15-LOX signaling. Biomed. Pharmacother. 2020, 127, 110151. [Google Scholar] [CrossRef]
- He, L.; Han, G.; Wu, S.; Du, S.; Zhang, Y.; Liu, W.; Jiang, B.; Zhang, L.; Xia, S.; Jia, S.; et al. Toll-like receptor 7 contributes to neuropathic pain by activating NF-κB in primary sensory neurons. Brainbehaviorand Immun. 2020, 87, 840–851. [Google Scholar] [CrossRef]
- Nashed, M.G.; Balenko, M.D.; Singh, G. Cancer-induced oxidative stress and pain. Curr. Pain Headache Rep. 2014, 18. [Google Scholar] [CrossRef]
- Kuete, V.; Sandjo, L.P.; Kwamou, G.M.N.; Wiench, B.; Nkengfack, A.E.; Efferth, T. Activity of three cytotoxic isoflavonoids from Erythrina excelsa and Erythrina senegalensis (neobavaisoflavone, sigmoidin H and isoneorautenol) toward multi-factorial drug resistant cancer cells. Phytomedicine 2014, 21, 682–688. [Google Scholar] [CrossRef]
- Xu, J.; Li, M.; Yao, Z.; Zhang, Y.; Li, S.; Hu, L.; Qin, Z.; Gonzalez, F.J.; Yao, X. In vitrometabolic mapping of neobavaisoflavone in human cytochromes P450 and UDP-glucuronosyltransferase enzymes by ultra high-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. J. Pharm. Biomed. Anal. 2018, 158, 351–360. [Google Scholar] [CrossRef]
- Ye, H.; He, X.; Feng, X. Developing neobavaisoflavone nanoemulsion suppresses lung cancer progression by regulating tumor microenvironment. Biomed. Pharmacother. 2020, 129. [Google Scholar] [CrossRef]
- Srdí-Rají, T.; Koní Ristí, A. Antioxidants: Role on Health and Prevention. Encycl. Food Health 2015, 227–233. [Google Scholar] [CrossRef]
- Wood, I.; Trostchansky, A.; Rubbo, H. Structural considerations on lipoxygenase function, inhibition and crosstalk with nitric oxide pathways. Biochimie 2020. [Google Scholar] [CrossRef]
- Kerkel, F.; Brock, D.; Touraud, D.; Kunz, W. Stabilisation of biofuels with hydrophilic, natural antioxidants solubilised by glycerol derivatives. Fuel 2021, 284, 119055. [Google Scholar] [CrossRef]
- Biela, M.; Rimarčík, J.; Senajová, E.; Kleinová, A.; Klein, E. Antioxidant action of deprotonated flavonoids: Thermodynamics of sequential proton-loss electron-transfer. Phytochemistry 2020, 180. [Google Scholar] [CrossRef]
- Wen, L.; Shi, D.; Zhou, T.; Tu, J.; He, M.; Jiang, Y.; Yang, B. Identification of two novel prenylated flavonoids in mulberry leaf and their bioactivities. Food Chem. 2020, 315. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, Y.; Yang, J.; He, J.; Sun, J.; Chen, F.; Zhang, M.; Yang, B. Prenylated flavonoids, promising nutraceuticals with impressive biological activities. Trends Food Sci. Technol. 2015, 44, 93–104. [Google Scholar] [CrossRef]
- Mvondo, M.A.; Njamen, D.; Kretzschmar, G.; Imma Bader, M.; Tanee Fomum, S.; Wandji, J.; Vollmer, G. Alpinumisoflavone and abyssinone v 4′-methylether derived from Erythrina lysistemon (Fabaceae) promote HDL-cholesterol synthesis and prevent cholesterol gallstone formation in ovariectomized rats. J. Pharm. Pharmacol. 2015, 67, 990–996. [Google Scholar] [CrossRef]
- Njamen, D.; Djiogue, S.; Zingue, S.; Mvondo, M.A.; Nkeh-Chungag, B.N. In vivo and in vitro estrogenic activity of extracts from Erythrina poeppigiana (Fabaceae). J. Complement. Integr. Med. 2013, 10, 63–73. [Google Scholar] [CrossRef]
- Kushwaha, P.P.; Prajapati, S.K.; Pothabathula, S.V.; Singh, A.K.; Shuaib, M.; Joshi, K.; Kumar, S. Prenylated Flavonoids as a Promising Drug Discovery Candidate; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 5, ISBN 9780128178911. [Google Scholar]
- Cui, L.; Lee, H.S.; Ndinteh, D.T.; Mbafor, J.T.; Kim, Y.H.; Le, T.V.T.; Nguyen, P.H.; Oh, W.K. New prenylated flavanones from erythrina abyssinica with protein tyrosine phosphatase 1B (PTP1B) inhibitory activity. Planta Med. 2010, 76, 713–718. [Google Scholar] [CrossRef]
- Khaomek, P.; Ichino, C.; Ishiyama, A.; Sekiguchi, H.; Namatame, M.; Ruangrungsi, N.; Saifah, E.; Kiyohara, H.; Otoguro, K.; Omura, S.; et al. In vitro antimalarial activity of prenylated flavonoids from Erythrina fusca. J. Nat. Med. 2008, 62, 217–220. [Google Scholar] [CrossRef]
- Ombito, J.O.; Majinda, R.R.T.; Masesane, I.B.; Bojase, G.; Schüffler, A.; Opatz, T. Prenylated isoflavones from the stem bark of Erythrina sacleuxii. Phytochem. Lett. 2018, 26, 110–114. [Google Scholar] [CrossRef]
- Passreiter, C.M.; Suckow-Schnitker, A.K.; Kulawik, A.; Addae-Kyereme, J.; Wright, C.W.; Wätjen, W. Prenylated flavanone derivatives isolated from Erythrina addisoniae are potent inducers of apoptotic cell death. Phytochemistry 2015, 117, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Togola, A.; Hedding, B.; Theis, A.; Wangensteen, H.; Rise, F.; Paulsen, B.S.; Diallo, D.; Malterud, K.E. 15-Lipoxygenase Inhibitory Effects of Prenylated Flavonoids From Erythrina Senegalensis. Planta Med. 2009, 75, 1168–1170. [Google Scholar] [CrossRef]
- Tuenter, E.; Zarev, Y.; Matheeussen, A.; Elgorashi, E.; Pieters, L.; Foubert, K. Antiplasmodial prenylated flavonoids from stem bark of Erythrina latissima. Phytochem. Lett. 2019, 30, 169–172. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Part Used | Folk Use | Administration | Reference |
|---|---|---|---|---|
| E. abyssinica | Bark | Inflammation, backache, pain and cramps lower belly | Decoction, external use, extract drunk and boiled in milk. | [25,27,33] |
| E. arborences | Branch, seed and leaves | Bone fracture and back pain | Paste/fomentation, decoction oral, juice of leaves | [31,32] |
| E. caffra | Bark, leaves and roots | Sprains, aches | Decoction oral, eardrops and plaster | [26,28,29] |
| E. caffra E. lysistemon | Stem bark and leaves | Toothache and earache | Oral infusion | [30] |
| E. edulis | Bark | Headache | Aqueous infusion drink | [34] |
| E. humeana | Bark | Spraians | Decoction, external use | [26] |
| E. senegalensis | Bark | Inflammation and Backache | Decoction, external use, massage with ointment | [22] |
| E. variegata | Leaves and bark | Fever, body ache, chronic bronchitis and otalgia | Decoction, oral | [35] |
| Species | Extract | Part | Model | Reference |
|---|---|---|---|---|
| E. addisoniae | EtOAc and MeOH | Stem bark | Inhibition of leukotriene B4 production from rat polymorphonuclear leukocytes. Cyclooxygenase-1 (COX-1) activity from human platelets. PLA2 induced paw oedema in mouse. TPA-induced mouse ear oedema. | [46] |
| E. lysistemon E. latissima E. humeana E. zeyheri | Ethanol and ethyl acetate | Leaves and bark | Cyclooxygenase-1 inhibition | [48] |
| E. indica | MeOH | Leaves | Carrageenan-induced hind paw edema | [59] |
| E. droogmansina | Ethyl acetate and MeOH | Root bark | Carrageenan-induced hind paw edema Ear edema induced by xylene Cotton pellet-induced granuloma | [45] |
| E. crista-galli | EtOH (70%) Dichlorometane MeOH | Aerial parts | Writhing test, Formalin test, Hot-plate | [60] |
| E. mildbraedii | Ethyl acetate | Root bark | Carrageenan-induced hind paw edema PLA2 induced paw oedema in mouse TPA-induced mouse ear oedema | [57] |
| E. mulungu | EtOH and EtOH 30% | Flowers and stem bark | Ovalbumin (OVA)-induced asthma in mice Dextran induced paw edema | [19,41,61] |
| E. senegalensis | Aqueous and EtOH (70%) | Bark and roots | Writhing test Egg albumin induced paw edema in rats. Hot-plate | [62,63] |
| E. sigmoidea | Chloroform | Bark | Inhibition of leukotriene B4 production from rat polymorphonuclear leukocytes. Cyclooxygenase-1 (COX-1) activity from human platelets. PLA2 induced paw oedema in mouse. TPA-induced mouse ear oedema. | [49] |
| E. variegata | MeOH, EtOH (95%), EtOH and Aqueous. | Leaves and bark | Writhing test, Tail-flik Carrageenan-induced hind paw edema Cotton pellet induced granuloma Hot plate HRBC membrane stabilization | [36,39,40] |
| E. velutina | EtOH (30%) and Aqueous | Stem bark and leaves | Writhing test, Formalin test, Hot-plate Carrageenan-induced hind paw edema | [41,62] |
| Species | Part | Identified Extract or Compounds | Model | Reference |
|---|---|---|---|---|
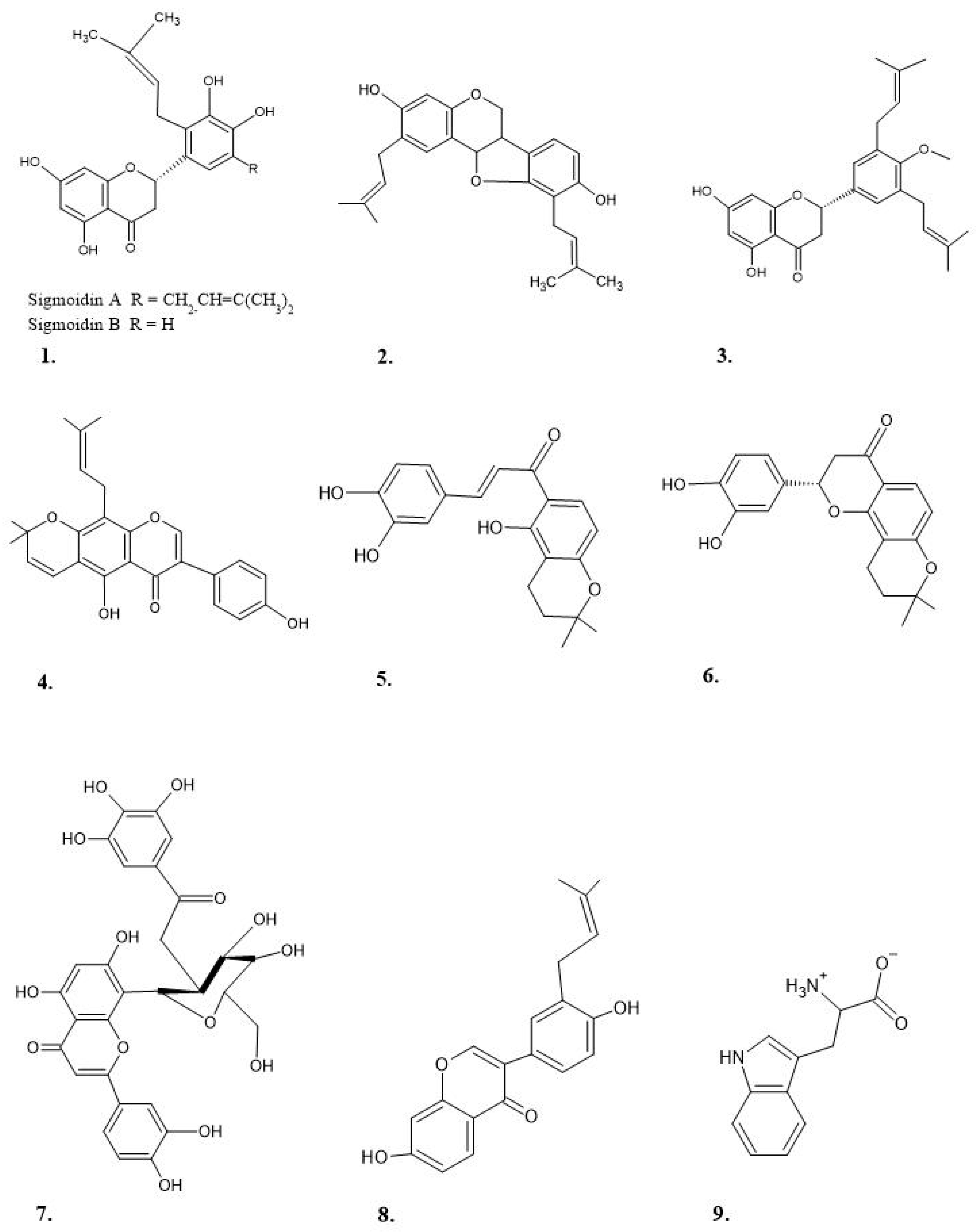
| E. abyssinica | Stem bark and root | Erycristagallin (4), 3-hydroxy-9-methoxy-10-(3,3-dimethylallyl) pterocarpene and 7,3′,4′-trihydroxy-5′-prenylflavanone (Abyssinone VII) | DPPH | [50,51] |
| E. burttii | Root bark | Burttinol-A and burttinol-C, and the 2-arylbenzofuran derivative burttinol-D | DPPH | [77] |
| E. crista-galli | Bark | Alkaloids, erythraline, erythrinine and hypaphorine | Inhibitory activity on LPS-induced nitric oxide (NO) | [72] |
| E. droogmansiana | Root bark | Genistein, 3-(3′,4′-methelenedioxyphenyl)-2,3-epoxypropanol, asperphenamate, Erydroogmansin B, vogelin C, Isolupalbigenin and erypostyrene | DPPH and FRAP | [78,79] |
| E. edulis | Seeds | Protein concentrate from the seed flour | ABTS, DPPH and ORAC | [80] |
| E. indica | Leaves and stem bark | Methanol extract | DPPH) and Nitric oxide scavenging assay | [52,53] |
| E. livingstoniana | Stem bark and twing | 7,3′-dihydroxy-4′-methoxy-5′-(3-methylbut-2-enyl) flavanone, 7, 3′,4′-trihydroxyflavanone and trans-3,4,2′,4′-tetrahydroxychalcone | DPPH | [65,81] |
| E. variegata | Leaves and bark | Methanolic extract and crude polysaccharides | DPPH, FRAP and TEAC | [67,82] |
| E. mildbraedi | Roots and bark | Flavanone (mildbone), chalcone (mildbenone) and Pterocarpene (Erycristagallin) | DPPH | [55,56,83] |
| E. senegalensis | Stem bark and leaves | Hydroalcoholic extract fraction 3 (polyphenols and flavonoids) and Methanol extract | DPPH, ABTS and FRAP | [84,85] |
| E. sigmoidea | Stem bark | Methanol extract; Flavanones, Sigmoidin A and Sigmoidin B | DPPH | [49,67] |
| E. stricta | Leaves | Hydromethanolic extract was | In vitro xanthine oxidase inhibitory activity All | [70] |
| E. suberosa | Flowers | Methanol extract | DPPH and Nitric oxide scavenging assay | [69] |
| E. vogelii | Leaves | Ethanol extract | DPPH | [67] |
| E. neillii | Leaves | Methanol total extract and its fractions | ORAC | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Cabrera, T.; Bautista, M.; Velázquez-González, C.; Jaramillo-Morales, O.A.; Guerrero-Solano, J.A.; Urrutia-Hernández, T.A.; De la O-Arciniega, M. Promising Antioxidant Activity of Erythrina Genus: An Alternative Treatment for Inflammatory Pain? Int. J. Mol. Sci. 2021, 22, 248. https://doi.org/10.3390/ijms22010248
Jiménez-Cabrera T, Bautista M, Velázquez-González C, Jaramillo-Morales OA, Guerrero-Solano JA, Urrutia-Hernández TA, De la O-Arciniega M. Promising Antioxidant Activity of Erythrina Genus: An Alternative Treatment for Inflammatory Pain? International Journal of Molecular Sciences. 2021; 22(1):248. https://doi.org/10.3390/ijms22010248
Chicago/Turabian StyleJiménez-Cabrera, Tania, Mirandeli Bautista, Claudia Velázquez-González, Osmar Antonio Jaramillo-Morales, José Antonio Guerrero-Solano, Thania Alejandra Urrutia-Hernández, and Minarda De la O-Arciniega. 2021. "Promising Antioxidant Activity of Erythrina Genus: An Alternative Treatment for Inflammatory Pain?" International Journal of Molecular Sciences 22, no. 1: 248. https://doi.org/10.3390/ijms22010248
APA StyleJiménez-Cabrera, T., Bautista, M., Velázquez-González, C., Jaramillo-Morales, O. A., Guerrero-Solano, J. A., Urrutia-Hernández, T. A., & De la O-Arciniega, M. (2021). Promising Antioxidant Activity of Erythrina Genus: An Alternative Treatment for Inflammatory Pain? International Journal of Molecular Sciences, 22(1), 248. https://doi.org/10.3390/ijms22010248