Promoting Effect of Basic Fibroblast Growth Factor in Synovial Mesenchymal Stem Cell-Based Cartilage Regeneration

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
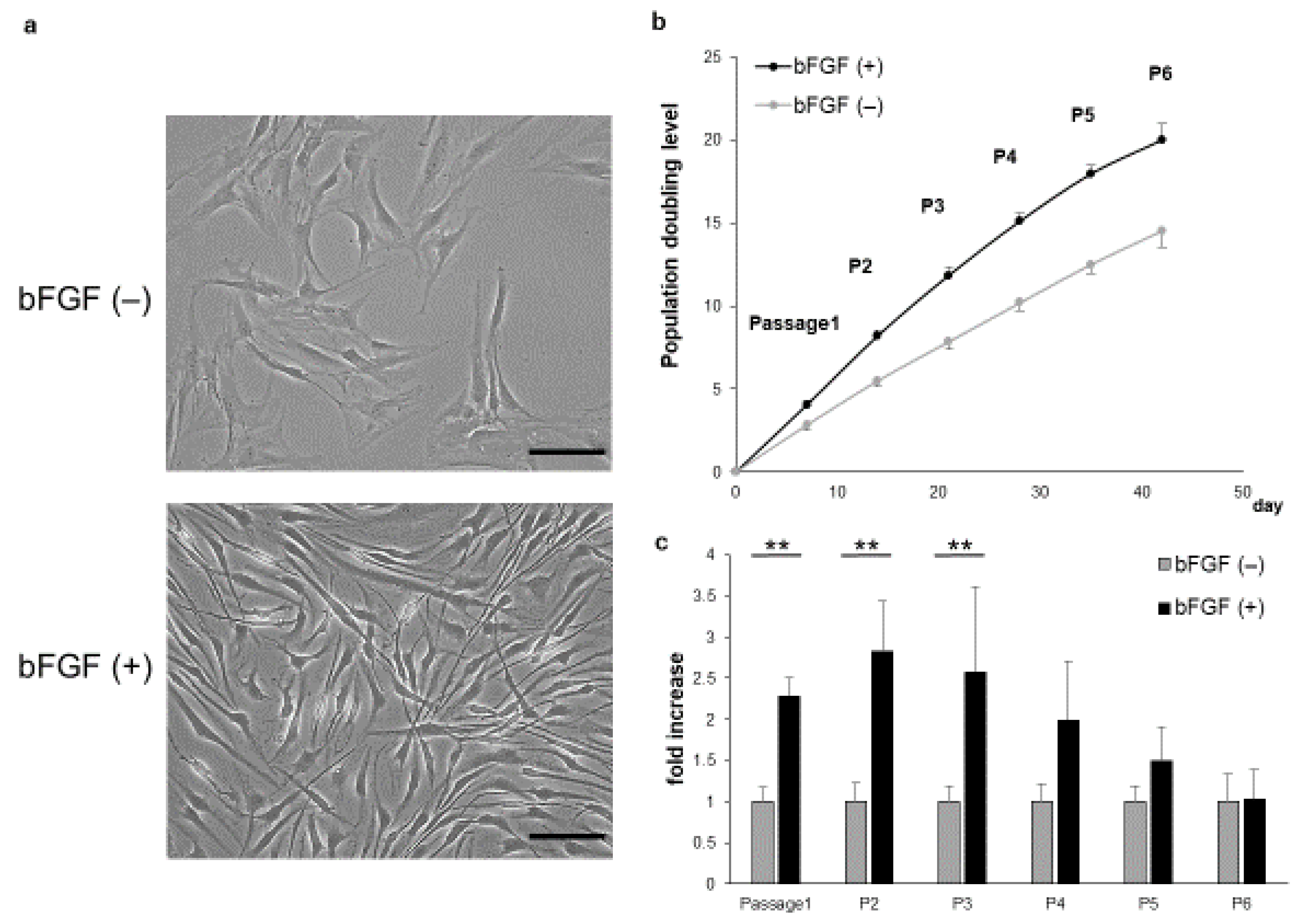
2.1. Proliferative Capacity and Cellular Morphology of SMSCs Cultured in the Presence or Absence of bFGF
2.2. Expression of FGFR3 Protein
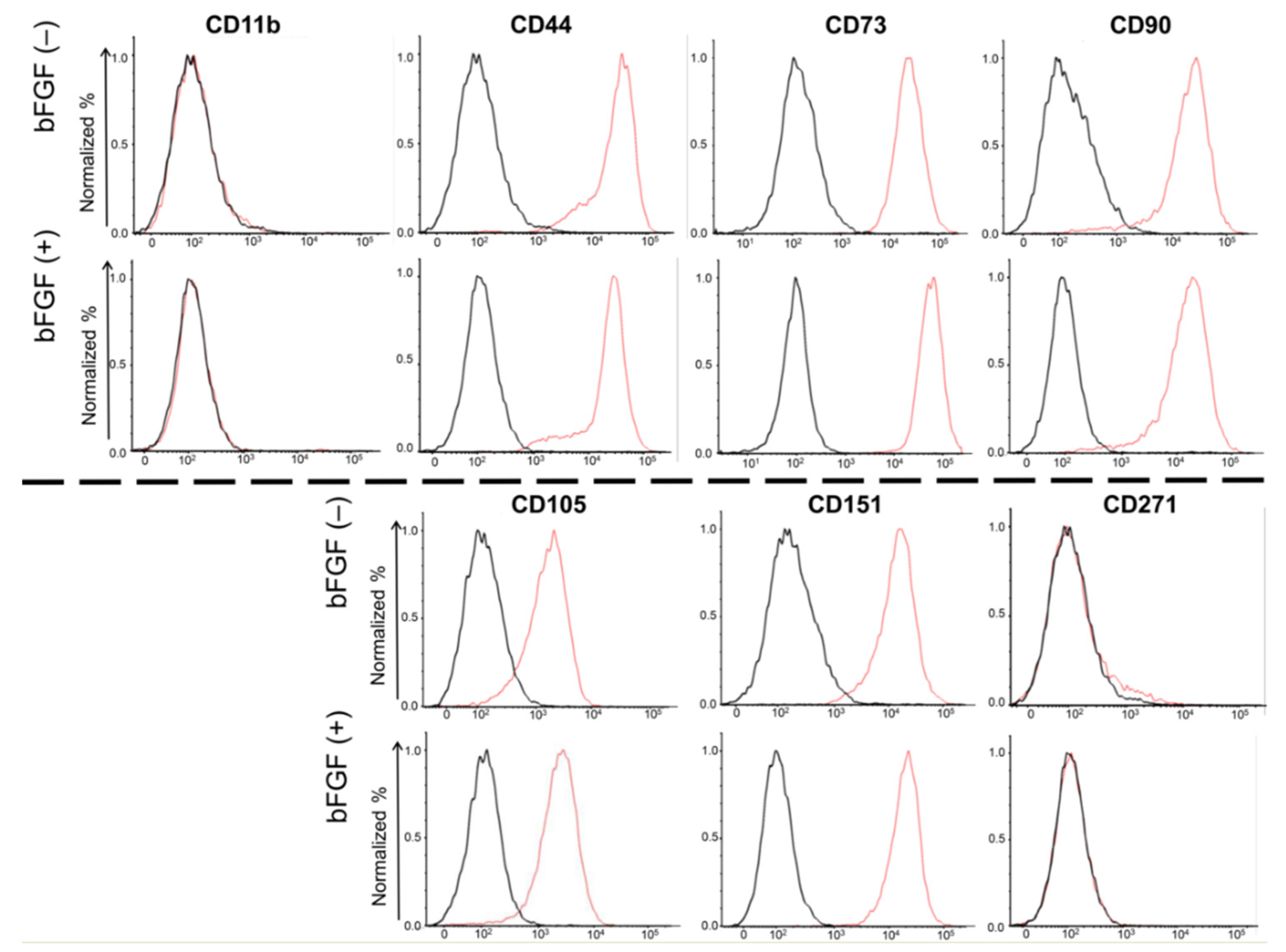
2.3. Expression of Cell Surface Markers
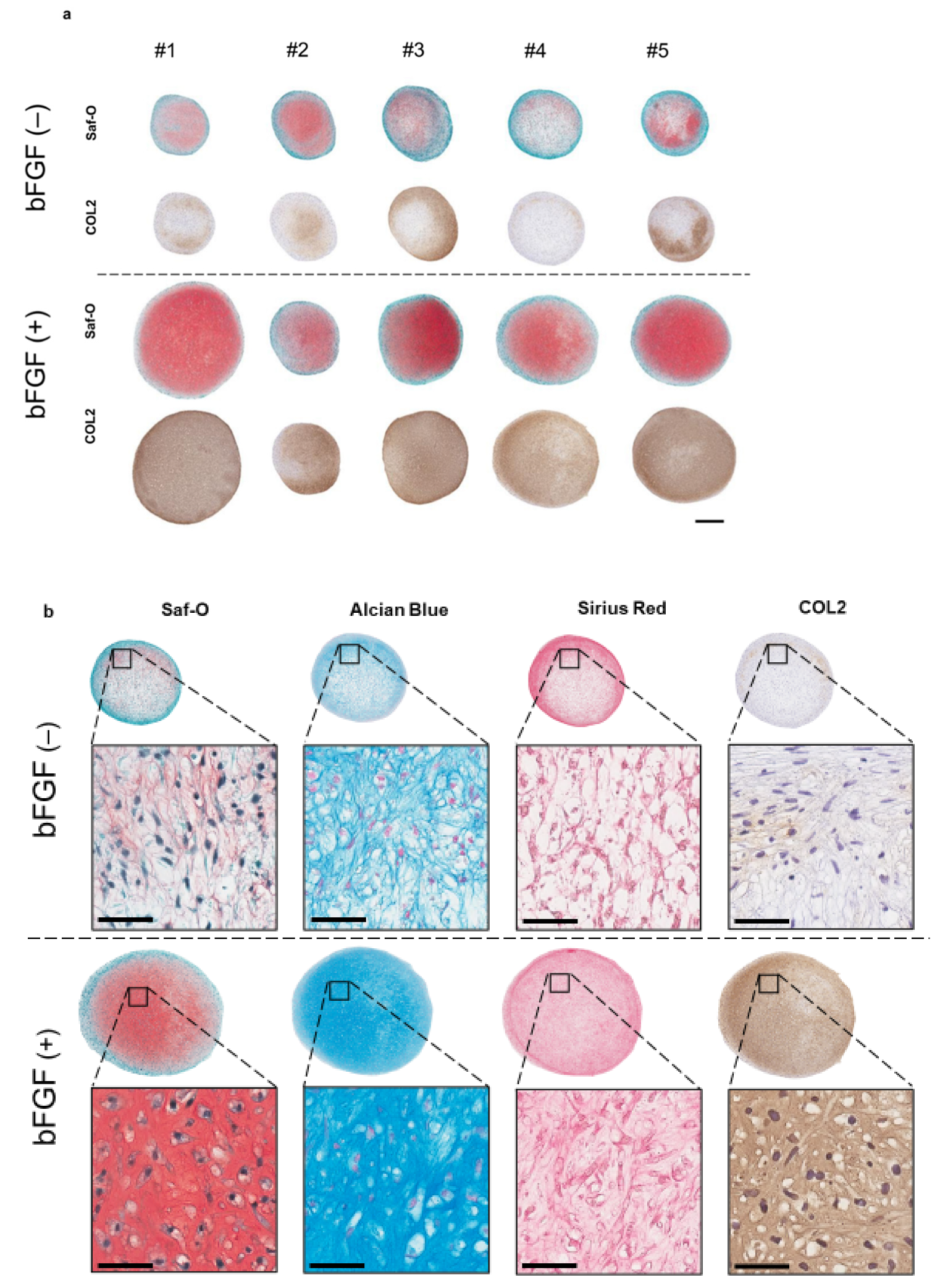
2.4. The Effect of bFGF on the Chondrogenesis of SMSCs in a Three-Dimensional (3D) Chondrogenic Culture
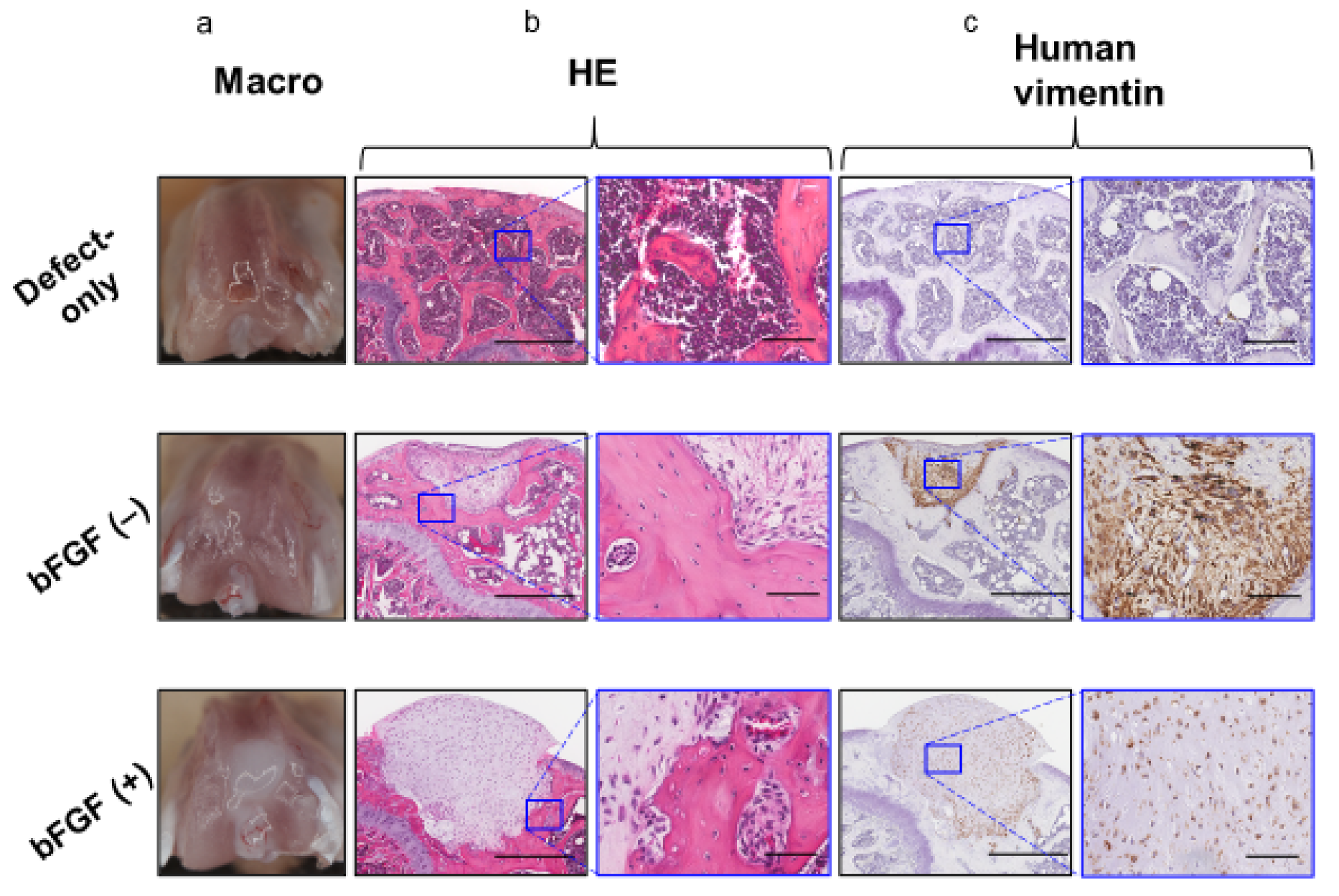
2.5. In vivo Osteochondral Repair in the Mouse Model Using Pellets Generated From the SMSCs From the bFGF (−) and bFGF (+) Groups
3. Discussion
4. Materials and Methods
4.1. Patient Demographics
4.2. Isolation and Culture of Human SMSCs
4.3. Cell-Proliferation Assay
4.4. Western Blotting
4.5. FACS Analysis
4.6. Chondrogenesis Assays
4.7. Implantation of Synovial Pellets onto Osteochondral Defects
4.8. Histology and Immunohistochemistry
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACI | autologous chondrocyte implantation |
| ACAN | aggrecan |
| ACLR | anterior cruciate ligament reconstruction |
| bFGF | basic fibroblast growth factor |
| BMP | bone morphogenetic protein |
| COL10 | collagen type X |
| COL10A1 | collagen type X alpha 1 |
| COL2 | collagen type II |
| COL2A1 | collagen type II alpha 1 |
| CSA | cross-section area |
| ECM | extracellular matrix |
| FACS | fluorescence-activated cell sorting |
| FGF | Fibroblast growth factor |
| FGFR | fibroblast growth factor receptor |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GM | growth medium |
| HE | hematoxylin and eosin |
| MSC | mesenchymal stem cell |
| OCD | osteochondritis dissecans |
| PDL | population doubling level |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| Saf-O | Safranin-O |
| scid | severe combined immunodeficient |
| sGAG | sulfated glycosaminoglycan |
| SMSC | Synovial mesenchymal stem cell |
| SOX9 | sex-determining region Y-box 9 |
| TEC | tissue-engineered construct |
| TGF-β | transforming growth factor β |
References
- Newman, A.P. Articular Cartilage Repair. Am. J. Sports Med. 1998, 26, 309–324. [Google Scholar] [CrossRef]
- Wakitani, S.; Mitsuoka, T.; Nakamura, N.; Toritsuka, Y.; Nakamura, Y.; Horibe, S. Autologous Bone Marrow Stromal Cell Transplantation for Repair of Full-Thickness Articular Cartilage Defects in Human Patellae: Two Case Reports. Cell Transplant. 2004, 13, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, C.H.; Gil Lee, Y.; Shin, W.H.; Kim, H.; Chai, J.W.; Jeong, E.C.; Kim, J.E.; Shim, H.; Shin, J.S.; Shin, I.S.; et al. Intra-Articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritis of the Knee: A Proof-of-Concept Clinical Trial. Stem Cells 2014, 32, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef]
- Segawa, Y.; Muneta, T.; Makino, H.; Nimura, A.; Mochizuki, T.; Ju, Y.J.; Ezura, Y.; Umezawa, A.; Sekiya, I. Mesenchymal stem cells derived from synovium, meniscus, anterior cruciate ligament, and articular chondrocytes share similar gene expression profiles. J. Orthop. Res. 2009, 27, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Koga, H.; Muneta, T.; Nagase, T.; Nimura, A.; Ju, Y.J.; Mochizuki, T.; Sekiya, I. Comparison of mesenchymal tissues-derived stem cells for in vivo chondrogenesis: Suitable conditions for cell therapy of cartilage defects in rabbit. Cell Tissue Res. 2008, 333, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, K.; Ando, W.; Tateishi, K.; Nansai, R.; Fujie, H.; Hart, D.A.; Kohda, H.; Kita, K.; Kanamoto, T.; Mae, T.; et al. The influence of skeletal maturity on allogenic synovial mesenchymal stem cell-based repair of cartilage in a large animal model. Biomaterials 2010, 31, 8004–8011. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, K.; Ebina, K.; Hart, D.A.; Hirao, M.; Noguchi, T.; Sugita, N.; Yasui, Y.; Chijimatsu, R.; Yoshikawa, H.; Nakamura, N. Synovial mesenchymal stem cells from osteo- or rheumatoid arthritis joints exhibit good potential for cartilage repair using a scaffold-free tissue engineering approach. Osteoarthr. Cartil. 2016, 24, 1413–1422. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, K.; Yasui, Y.; Koizumi, K.; Chijimatsu, R.; Hart, D.A.; Yonetani, Y.; Ando, W.; Nishii, T.; Kanamoto, T.; Horibe, S.; et al. First-in-Human Pilot Study of Implantation of a Scaf-fold-Free Tissue-Engineered Construct Generated From Autologous Synovial Mesenchymal Stem Cells for Repair of Knee Chondral Lesions. Am. J. Sports Med. 2018, 46, 2384–2393. [Google Scholar] [CrossRef]
- Narcisi, R.; Cleary, M.A.; Brama, P.A.; Hoogduijn, M.J.; Tüysüz, N.; Berge, D.T.; Van Osch, G.J.V.M. Long-Term Expansion, Enhanced Chondrogenic Potential, and Suppression of Endochondral Ossification of Adult Human MSCs via WNT Signaling Modulation. Stem Cell Rep. 2015, 4, 459–472. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Han, S.; Kim, E.; Lee, S.; Seong, S.; Lee, M.C. Chondrogenic potentials of human synovium-derived cells sorted by specific surface markers. Osteoarthr. Cartil. 2013, 21, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Kim, J.H.; Jo, C.H.; Seong, S.C.; Lee, J.C.; Lee, M.C. Effect of serum and growth factors on chondrogenic differen-tiation of synovium-derived stromal cells. Tissue Eng. Part A 2009, 15, 3401–3415. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Chijimatsu, R.; Hart, D.A.; Koizumi, K.; Sugita, N.; Shimomura, K.; Myoui, A.; Yoshikawa, H.; Nakamura, N. Preparation of Scaffold-Free Tissue-Engineered Constructs Derived from Human Synovial Mesenchymal Stem Cells Under Low Oxygen Tension Enhances Their Chondrogenic Differentiation Capacity. Tissue Eng. Part A 2016, 22, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, S.; Shimazu, A.; Miyazaki, K.; Pan, H.; Koike, C.; Yoshida, E.; Takagishi, K.; Kato, Y. Retention of Multilineage Differentiation Potential of Mesenchymal Cells during Proliferation in Response to FGF. Biochem. Biophys. Res. Commun. 2001, 288, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solchaga, L.A.; Penick, K.; Porter, J.D.; Goldberg, V.M.; Caplan, A.I.; Welter, J.F. FGF-2 enhances the mitotic and chon-drogenic potentials of human adult bone marrow-derived mesenchymal stem cells. J. Cell Physiol. 2005, 203, 398–409. [Google Scholar] [CrossRef]
- Buckley, C.; Kelly, D.J. Expansion in the presence of FGF-2 enhances the functional development of cartilaginous tissues engineered using infrapatellar fat pad derived MSCs. J. Mech. Behav. Biomed. Mater. 2012, 11, 102–111. [Google Scholar] [CrossRef]
- Dvořak, P.; Hampl, A. Basic fibroblast growth factor and its receptors in human embryonic stem cells. Folia Histochem. Cytobiol. 2005, 43, 203–208. [Google Scholar]
- Maric, D.; Pla, A.F.; Chang, Y.H.; Barker, J.L. Self-Renewing and Differentiating Properties of Cortical Neural Stem Cells Are Selectively Regulated by Basic Fibroblast Growth Factor (FGF) Signaling via Specific FGF Receptors. J. Neurosci. 2007, 27, 1836–1852. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, M.C.; Seong, S.C.; Park, K.H.; Lee, S. Enhanced Proliferation and Chondrogenic Differentiation of Human Synovium-Derived Stem Cells Expanded with Basic Fibroblast Growth Factor. Tissue Eng. Part A 2011, 17, 991–1002. [Google Scholar] [CrossRef]
- Tojyo, I.; Yamaguti, A.; Ozaki, H.; Yoshida, H.; Fujita, S. The expression of fibroblast growth factor receptor-3 in synovial osteochondromatosis of the temporomandibular joint. Arch. Oral Biol. 2004, 49, 591–594. [Google Scholar] [CrossRef]
- Itoh, N.; Ornitz, D.M. Evolution of the Fgf and Fgfr gene families. Trends Genet. 2004, 20, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Maddaluno, L.; Urwyler, C.; Werner, S. Fibroblast growth factors: Key players in regeneration and tissue repair. Development 2017, 144, 4047–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornitz, D.; Itoh, N.M. Fibroblast growth factors. Genome Biol. 2001, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M. FGFR inhibitors: Effects on cancer cells, tumor microenvironment and whole-body homeostasis (Review). Int. J. Mol. Med. 2016, 38, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; De Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2005, 97, 33–44. [Google Scholar] [CrossRef]
- Murakami, S.; Kan, M.; McKeehan, W.L.; De Crombrugghe, B. Up-regulation of the chondrogenic Sox9 gene by fibroblast growth factors is mediated by the mitogen-activated protein kinase pathway. Proc. Natl. Acad. Sci. USA 2000, 97, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Wang, C.; Acton, A.J.; Eckert, G.J.; Trippel, S.B. Role of Sox9 in Growth Factor Regulation of Articular Chondrocytes. J. Cell. Biochem. 2015, 116, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Osathanon, T.; Nowwarote, N.; Pavasant, P. Basic fibroblast growth factor inhibits mineralization but induces neuronal dif-ferentiation by human dental pulp stem cells through a FGFR and PLCgamma signaling pathway. J. Cell. Biochem. 2011, 112, 1807–1816. [Google Scholar] [CrossRef]
- Nowwarote, N.; Sukarawan, W.; Pavasant, P.; Foster, B.L.; Osathanon, T. Basic fibroblast growth factor regulates phos-phate/pyrophosphate regulatory genes in stem cells isolated from human exfoliated deciduous teeth. Stem Cell Res 2018, 9, 345. [Google Scholar]
- Ito, T.; Sawada, R.; Fujiwara, Y.; Seyama, Y.; Tsuchiya, T. FGF-2 suppresses cellular senescence of human mesenchymal stem cells by down-regulation of TGF-beta2. Biochem. Biophys. Res. Commun. 2007, 359, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Rombouts, W.J.C.; Ploemacher, R.E. Primary murine MSC show highly efficient homing to the bone marrow but lose homing ability following culture. Leukemia 2003, 17, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepúlveda, J.C.; Tomé, M.; Fernández, M.E.; Delgado, M.; Campisi, J.; Bernad, A.; González, M.A. Cell senescence abrogates the therapeutic potential of human mesenchymal stem cells in the lethal endotoxemia model. Steam Cells 2014, 32, 1865–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, A.; Tamamura, Y.; Morioka, M.; Karagiannis, P.; Shima, N.; Tsumaki, N. Considerations in hiPSC-derived cartilage for articular cartilage repair. Inflamm. Regen. 2018, 38, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I.; Bruder, S.P. Mesenchymal stem cells: Building blocks for molecular medicine in the 21st century. Trends Mol. Med. 2001, 7, 259–264. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Önfelt, B.; Nedvetzki, S.; Benninger, R.K.P.; Purbhoo, M.A.; Sowinski, S.; Hume, A.N.; Seabra, M.C.; Neil, M.A.A.; French, P.M.W.; Davis, D.M. Structurally Distinct Membrane Nanotubes between Human Macrophages Support Long-Distance Vesicular Traffic or Surfing of Bacteria. J. Immunol. 2006, 177, 8476–8483. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and mi-croRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Fellows, C.R.; Matta, C.; Zakany, R.; Khan, I.M.; Mobasheri, A. Adipose, Bone Marrow and Synovial Joint-Derived Mes-enchymal Stem Cells for Cartilage Repair. Front. Genet. 2016, 7, 213. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, N.; Muneta, T.; Koga, H.; Nakagawa, Y.; Mizuno, M.; Tsuji, K.; Mabuchi, Y.; Akazawa, C.; Kobayashi, E.; Matsumoto, K.; et al. Not single but periodic injections of synovial mesenchymal stem cells maintain viable cells in knees and inhibit osteoarthritis progression in rats. Osteoarthr. Cartil. 2016, 24, 1061–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Sekiya, I.; Muneta, T.; Hatsushika, D.; Horie, M.; Tsuji, K.; Kawarasaki, T.; Watanabe, A.; Hishikawa, S.; Fujimoto, Y.; et al. Arthroscopic, histological and MRI analyses of cartilage repair after a minimally invasive method of transplantation of allogeneic synovial mesenchymal stromal cells into cartilage defects in pigs. Cytotherapy 2012, 14, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filová, E.; Rampichová, M.; Litvinec, A.; Držík, M.; Míčková, A.; Buzgo, M.; Košťáková, E.; Martinová, L.; Usvald, D.; Prosecká, E.; et al. A cell-free nanofiber composite scaffold regenerated osteochondral de-fects in miniature pigs. Int. J. Pharm. 2013, 447, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Cucchiarini, M.; Madry, H.; Ma, C.; Thurn, T.; Zurakowski, D.; Menger, M.D.; Kohn, D.; Trippel, S.B.; Terwilliger, E.F. Improved tissue repair in articular cartilage defects in vivo by rAAV-mediated overexpression of human fibroblast growth factor 2. Mol. Ther. J. Am. Soc. Gene Ther. 2005, 12, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Ando, W.; Tateishi, K.; Hart, D.A.; Katakai, D.; Tanaka, Y.; Nakata, K.; Hashimoto, J.; Fujie, H.; Shino, K.; Yoshikawa, H.; et al. Cartilage repair using an in vitro generated scaffold-free tissue-engineered construct derived from porcine synovial mesenchymal stem cells. Biomaterials 2007, 28, 5462–5470. [Google Scholar] [CrossRef]
- Chijimatsu, R.; Kobayashi, M.; Ebina, K.; Iwahashi, T.; Okuno, Y.; Hirao, M.; Fukuhara, A.; Nakamura, N.; Yoshikawa, H. Impact of dexamethasone concentration on cartilage tissue formation from human synovial derived stem cells in vitro. Cytotechnology 2018, 70, 819–829. [Google Scholar] [CrossRef]
- Cheng, T.; Yang, C.; Weber, N.; Kim, H.T.; Kuo, A.C. Fibroblast growth factor 2 enhances the kinetics of mesenchymal stem cell chondrogenesis. Biochem. Biophys. Res. Commun. 2012, 426, 544–550. [Google Scholar] [CrossRef]
- Toh, W.S.; Guo, X.M.; Choo, A.B.; Lu, K.; Lee, E.H.; Cao, T. Differentiation and enrichment of expandable chondrogenic cells from human embryonic stem cells in vitro. J. Cell. Mol. Med. 2009, 13, 3570–3590. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Ebina, K.; Hirao, M.; Morimoto, T.; Koizumi, K.; Kitaguchi, K.; Matsuoka, H.; Iwahashi, T.; Yoshikawa, H. Oxygen ultrafine bubbles water administration prevents bone loss of glucocorticoid-induced osteoporosis in mice by suppressing osteoclast differentiation. Osteoporos Int. 2017, 28, 1063–1075. [Google Scholar] [CrossRef]
- Shirasawa, S.; Sekiya, I.; Sakaguchi, Y.; Yagishita, K.; Ichinose, S.; Muneta, T. In vitro chondrogenesis of human synovi-um-derived mesenchymal stem cells: Optimal condition and comparison with bone marrow-derived cells. J. Cell. Biochem. 2006, 97, 84–97. [Google Scholar] [CrossRef]
- Chijimatsu, R.; Ikeya, M.; Yasui, Y.; Ikeda, Y.; Ebina, K.; Moriguchi, Y.; Shimomura, K.; Hart, D.A.; Yoshikawa, H.; Nakamura, N. Characterization of Mesenchymal Stem Cell-Like Cells Derived From Human iPSCs via Neural Crest Development and Their Application for Osteochondral Repair. Stem Cells Int. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaguchi, K.; Kashii, M.; Ebina, K.; Kaito, T.; Okada, R.; Makino, T.; Noguchi, T.; Ishimoto, T.; Nakano, T.; Yoshikawa, H. Effects of single or combination therapy of teriparatide and anti-RANKL monoclonal antibody on bone defect regeneration in mice. Bone 2018, 106, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, K.; Moriguchi, Y.; Ando, W.; Nansai, R.; Fujie, H.; Hart, D.A.; Gobbi, A.; Kita, K.; Horibe, S.; Shino, K.; et al. Osteochondral repair using a scaffold-free tissue-engineered construct derived from synovial mes-enchymal stem cells and a hydroxyapatite-based artificial bone. Tissue Eng. Part A 2014, 20, 2291–2304. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, S.W.; Keeley, F.W.; Salter, R.B. Durability of regenerated articular cartilage produced by free autogenous periosteal grafts in major full-thickness defects in joint surfaces under the influence of continuous passive motion. A follow-up report at one year. J. Bone Jt. Surg. Am. Vol. 1988, 70, 595–606. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2012, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological score description | Defect only [n =12] | bFGF (−) [n =13] | bFGF (+) [n =13] |
| (possible score) | |||
| Cartilage repair | |||
| Cellular morphology (0–4) | 0.7 ± 0.9 | 1.8 ± 0.5 | 2.6 ± 0.9 |
| Safranin-O staining (0–3) | 0.5 ± 0.5 | 0.5 ± 0.5 | 1.5 ± 0.7 |
| Surface regularity (0–3) | 0.8 ± 0.4 | 0.8 ± 0.4 | 1.8 ± 0.6 |
| Structural integrity (0–2) | 0.9 ± 0.3 | 1.1 ± 0.3 | 1.2 ± 0.4 |
| Thickness (0–2) | 0.4 ± 0.5 | 0.6 ± 0.5 | 1.1 ± 0.6 |
| Bonding to adjacent cartilage (0–2) | 1.4 ± 0.6 | 1.8 ± 0.4 | 1.3 ± 0.6 |
| Hypocellularity (0–3) | 0.4 ± 0.5 | 0.8 ± 0.4 | 1.0 ± 0.4 |
| Chondrocyte clustering (0–2) | 1.9 ± 0.3 | 1.9 ± 0.3 | 1.9 ± 0.3 |
| Freedom from degeneration | |||
| of adjacent cartilage (0–3) | 1.8 ± 0.6 | 1.8 ± 0.4 | 2.1 ± 0.5 |
| Subtotal score (Cartilage; 0–24) | 8.8 ± 2.6 | * 11.2 ± 1.5 | **† 14.5 ± 2.6 |
| Subchondral bone repair | |||
| Subchondral bone alignment (0–2) | 1.3 ± 0.4 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Bone integration (0–2) | 1.2 ± 0.4 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Bone infiltration into defect area (0–2) | 1.9 ± 0.3 | 0.7 ± 0.5 | 0.8 ± 0.4 |
| Tidemark continuity (0–2) | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Cellular morphology (0–2) | 1.5 ± 0.5 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Exposure of subchondral bone (0–2) | 1.8 ± 0.4 | 2.0 ± 0.0 | 2.0 ± 0.0 |
| Subtotal score (Subchondral bone; 0–12) | 7.6 ± 1.1 | ** 2.7 ± 0.5 | ** 2.8 ± 0.4 |
| Total Score (0–36) | 16.3 ± 3.5 | 13.9 ± 1.5 | † 17.3 ± 2.6 |
| Patient No. | Age, y | Gender | Surgery | Waiting Period, w |
|---|---|---|---|---|
| #1 | 16 | Female | ACLR | 14 |
| #2 | 13 | Male | OCD 2nd look | 15 |
| #3 | 18 | Female | ACLR | 13 |
| #4 | 18 | Female | ACLR | 20 |
| #5 | 22 | Female | ACLR | 4 |
| mean | 17.4 | 13.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okamura, G.; Ebina, K.; Hirao, M.; Chijimatsu, R.; Yonetani, Y.; Etani, Y.; Miyama, A.; Takami, K.; Goshima, A.; Yoshikawa, H.; et al. Promoting Effect of Basic Fibroblast Growth Factor in Synovial Mesenchymal Stem Cell-Based Cartilage Regeneration. Int. J. Mol. Sci. 2021, 22, 300. https://doi.org/10.3390/ijms22010300
Okamura G, Ebina K, Hirao M, Chijimatsu R, Yonetani Y, Etani Y, Miyama A, Takami K, Goshima A, Yoshikawa H, et al. Promoting Effect of Basic Fibroblast Growth Factor in Synovial Mesenchymal Stem Cell-Based Cartilage Regeneration. International Journal of Molecular Sciences. 2021; 22(1):300. https://doi.org/10.3390/ijms22010300
Chicago/Turabian StyleOkamura, Gensuke, Kosuke Ebina, Makoto Hirao, Ryota Chijimatsu, Yasukazu Yonetani, Yuki Etani, Akira Miyama, Kenji Takami, Atsushi Goshima, Hideki Yoshikawa, and et al. 2021. "Promoting Effect of Basic Fibroblast Growth Factor in Synovial Mesenchymal Stem Cell-Based Cartilage Regeneration" International Journal of Molecular Sciences 22, no. 1: 300. https://doi.org/10.3390/ijms22010300
APA StyleOkamura, G., Ebina, K., Hirao, M., Chijimatsu, R., Yonetani, Y., Etani, Y., Miyama, A., Takami, K., Goshima, A., Yoshikawa, H., Ishimoto, T., Nakano, T., Hamada, M., Kanamoto, T., & Nakata, K. (2021). Promoting Effect of Basic Fibroblast Growth Factor in Synovial Mesenchymal Stem Cell-Based Cartilage Regeneration. International Journal of Molecular Sciences, 22(1), 300. https://doi.org/10.3390/ijms22010300