Phenotypic Characterization of Transgenic Mice Expressing Human IGFBP-5


Abstract
:1. Introduction
2. Results
2.1. Generation of Transgenic IGFBP-5 Mice
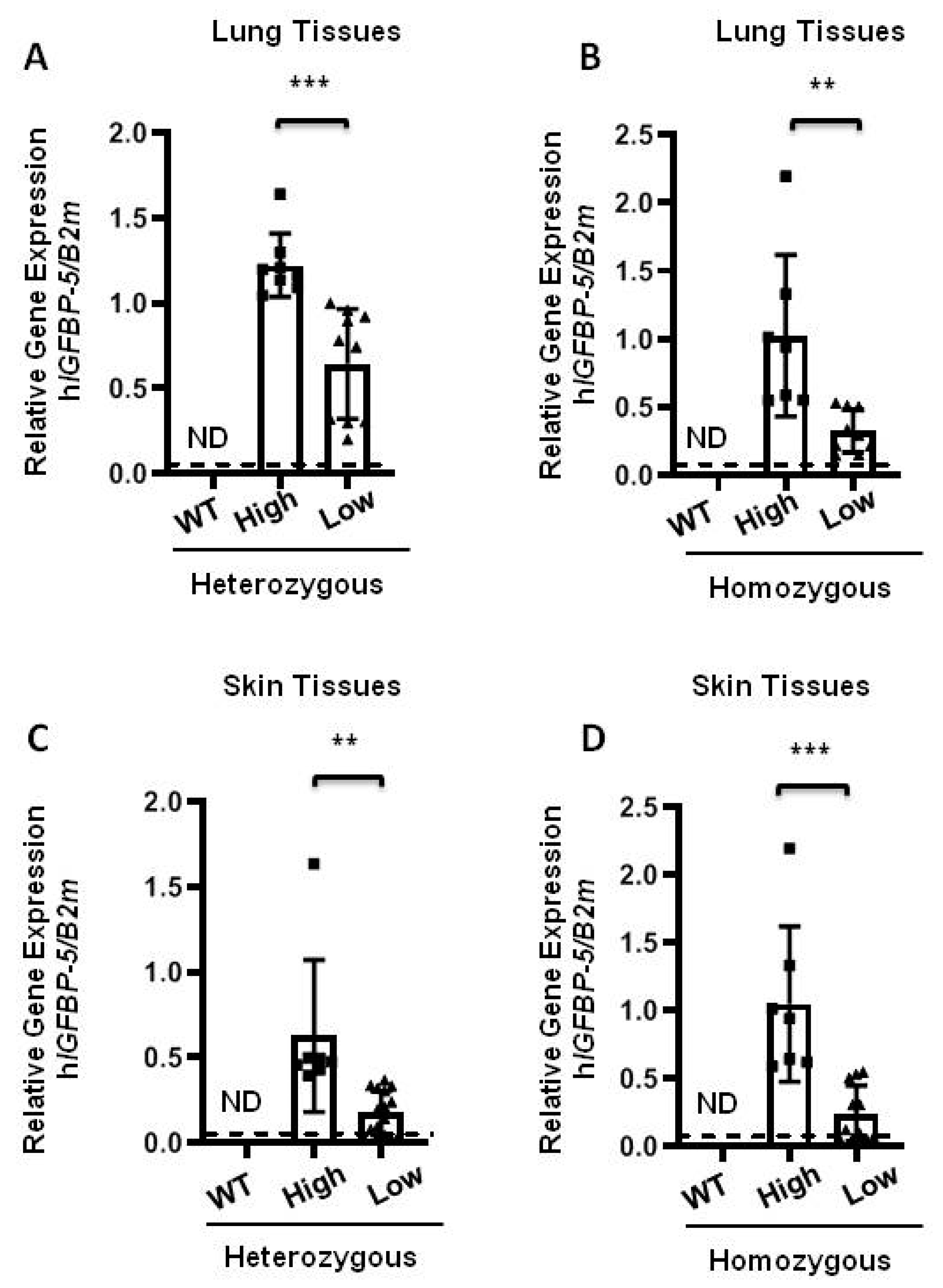
2.2. Expression of Human IGFBP-5
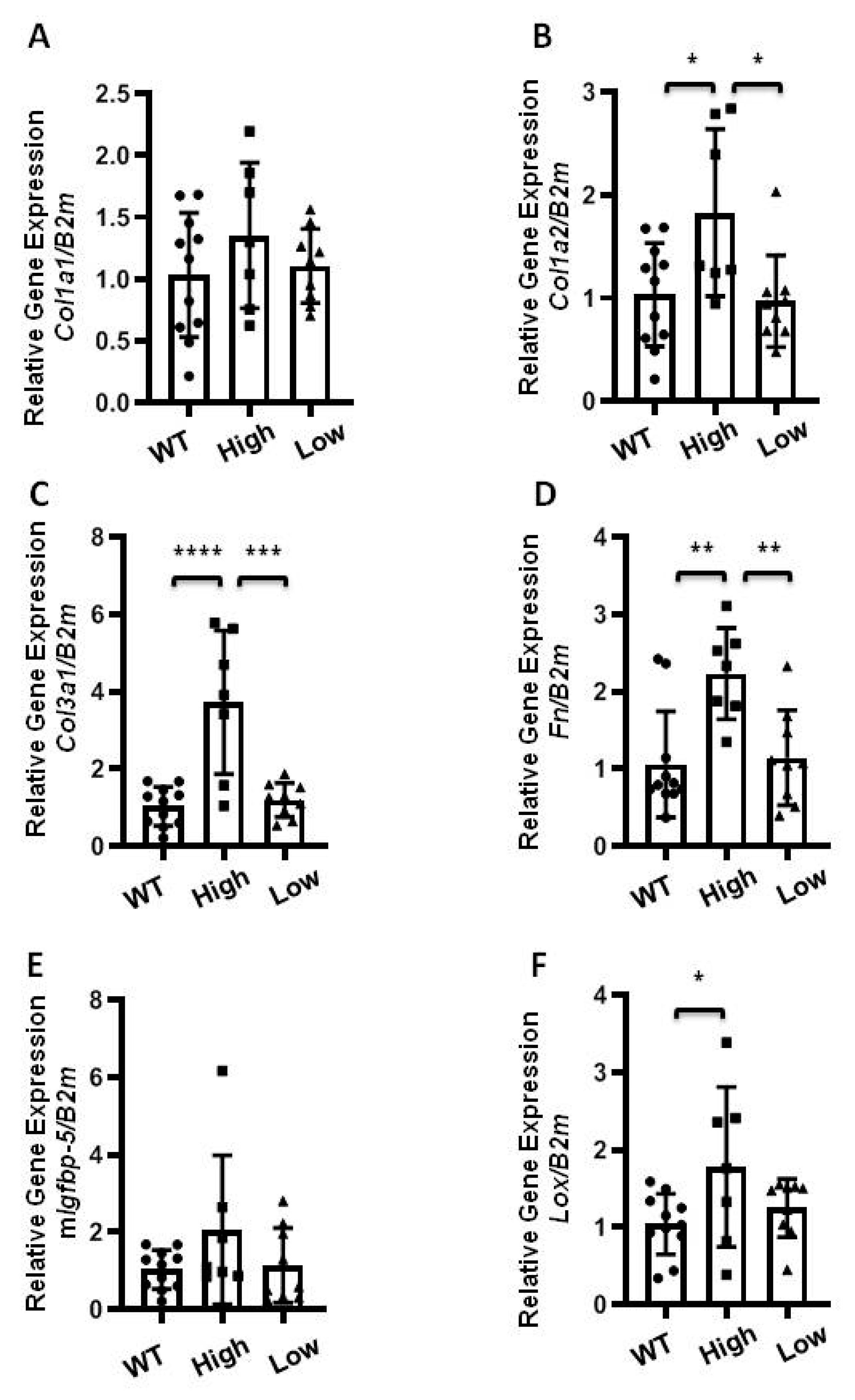
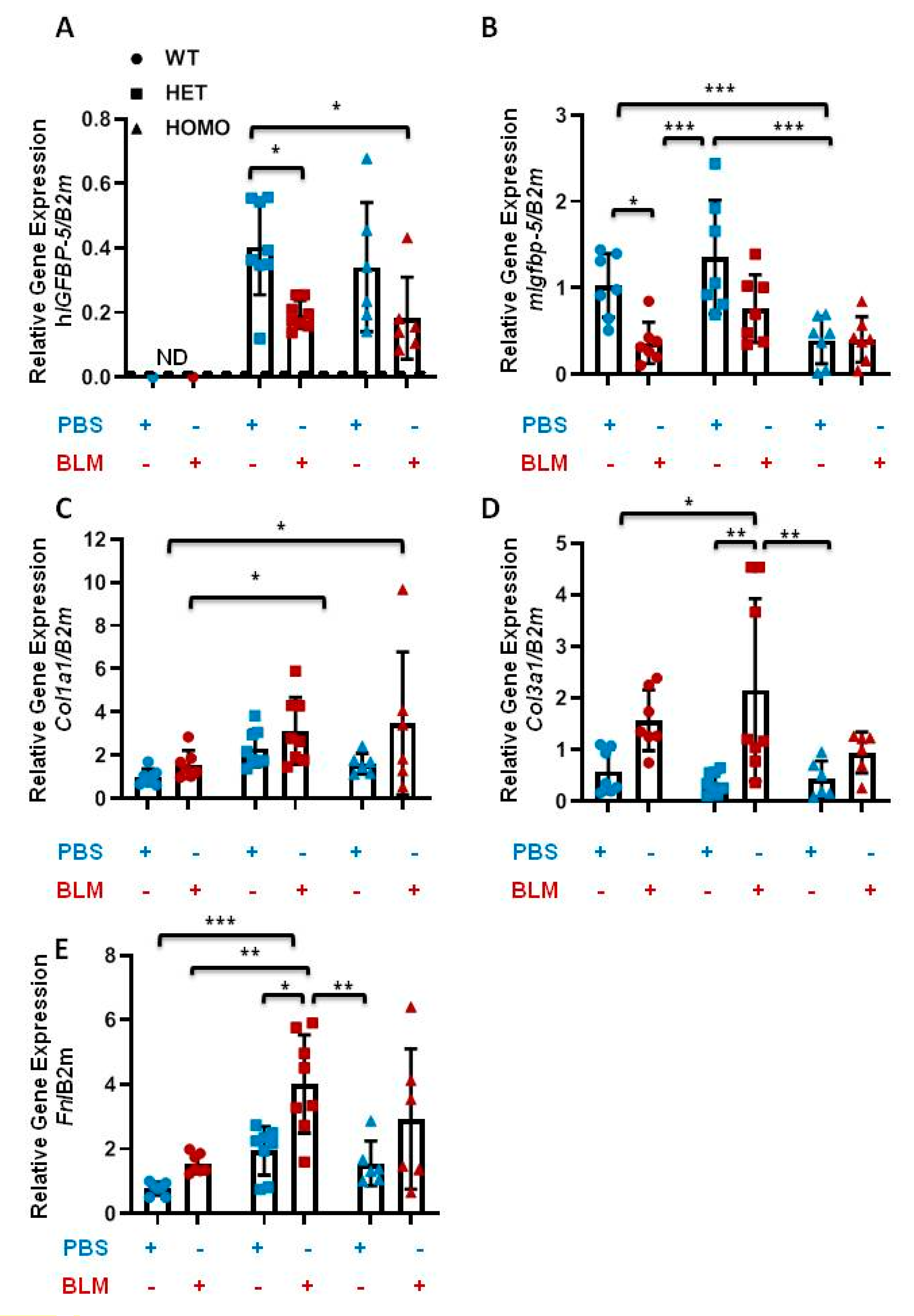
2.3. The Effect of Transgenic hIGFBP-5 on Extracellular Matrix Gene Expression in Lung
2.4. The Effect of Transgenic hIGFBP-5 on Extracellular Matrix Gene Expression in Skin
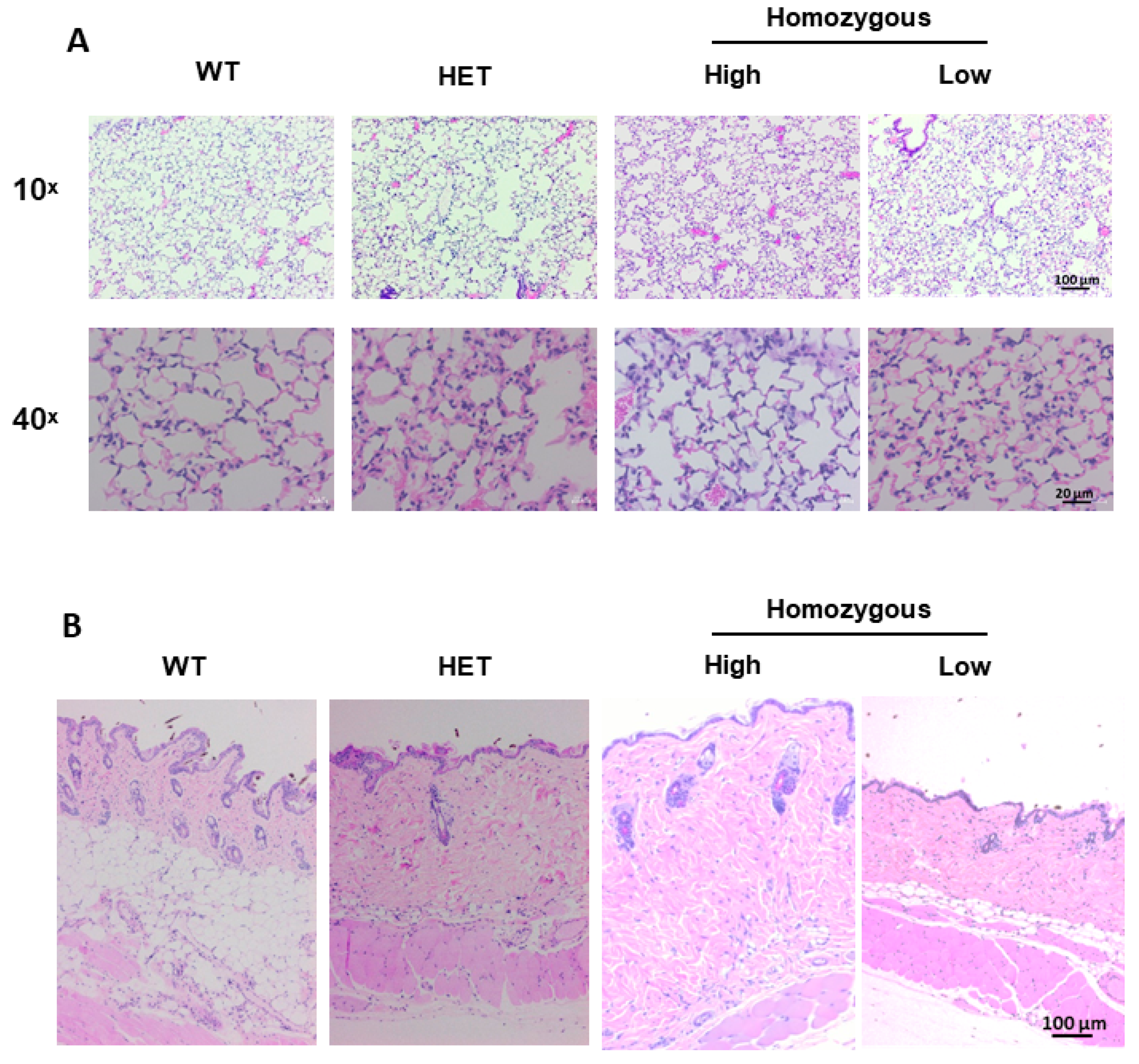
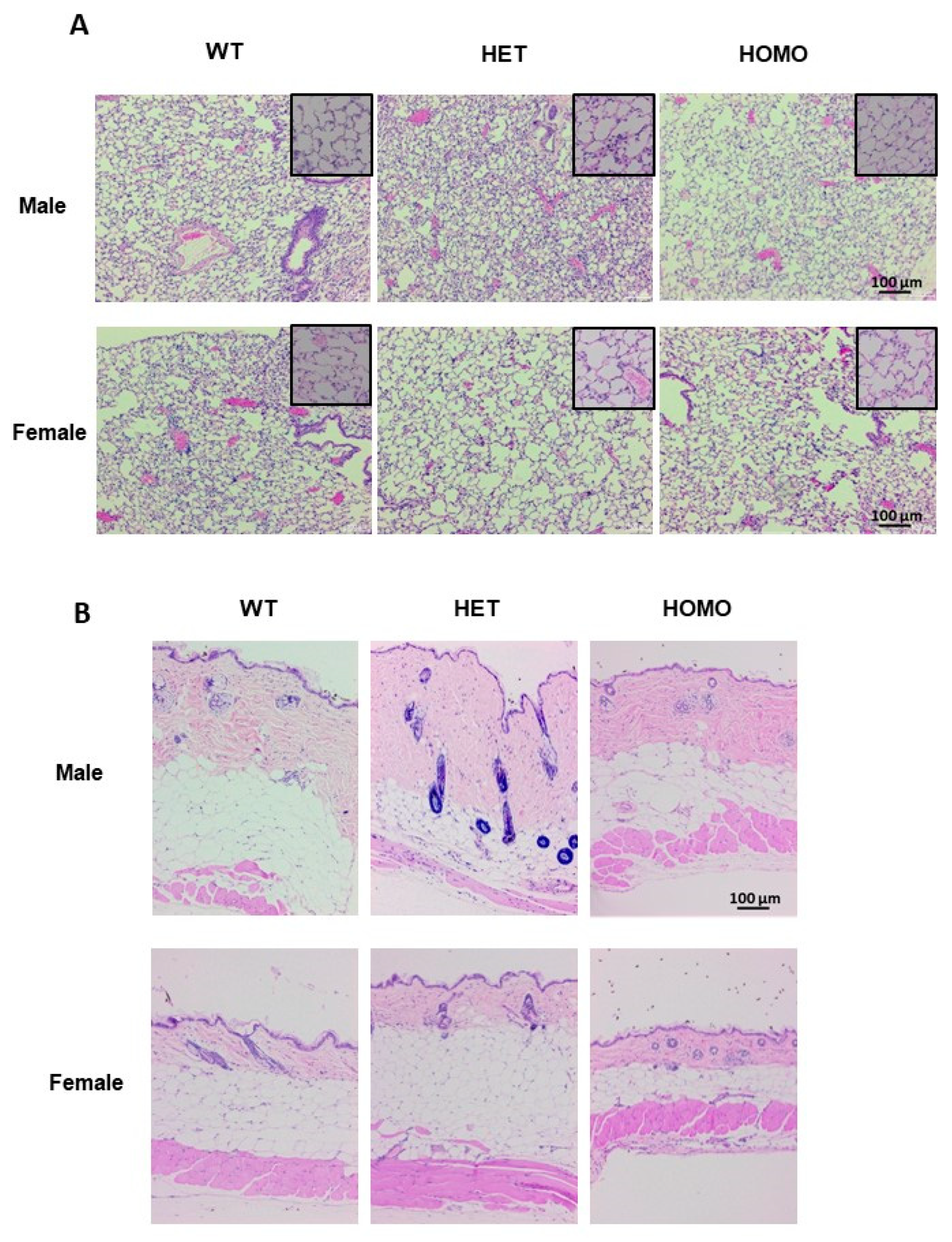
2.5. Histological Evaluation of Lung and Skin Tissues
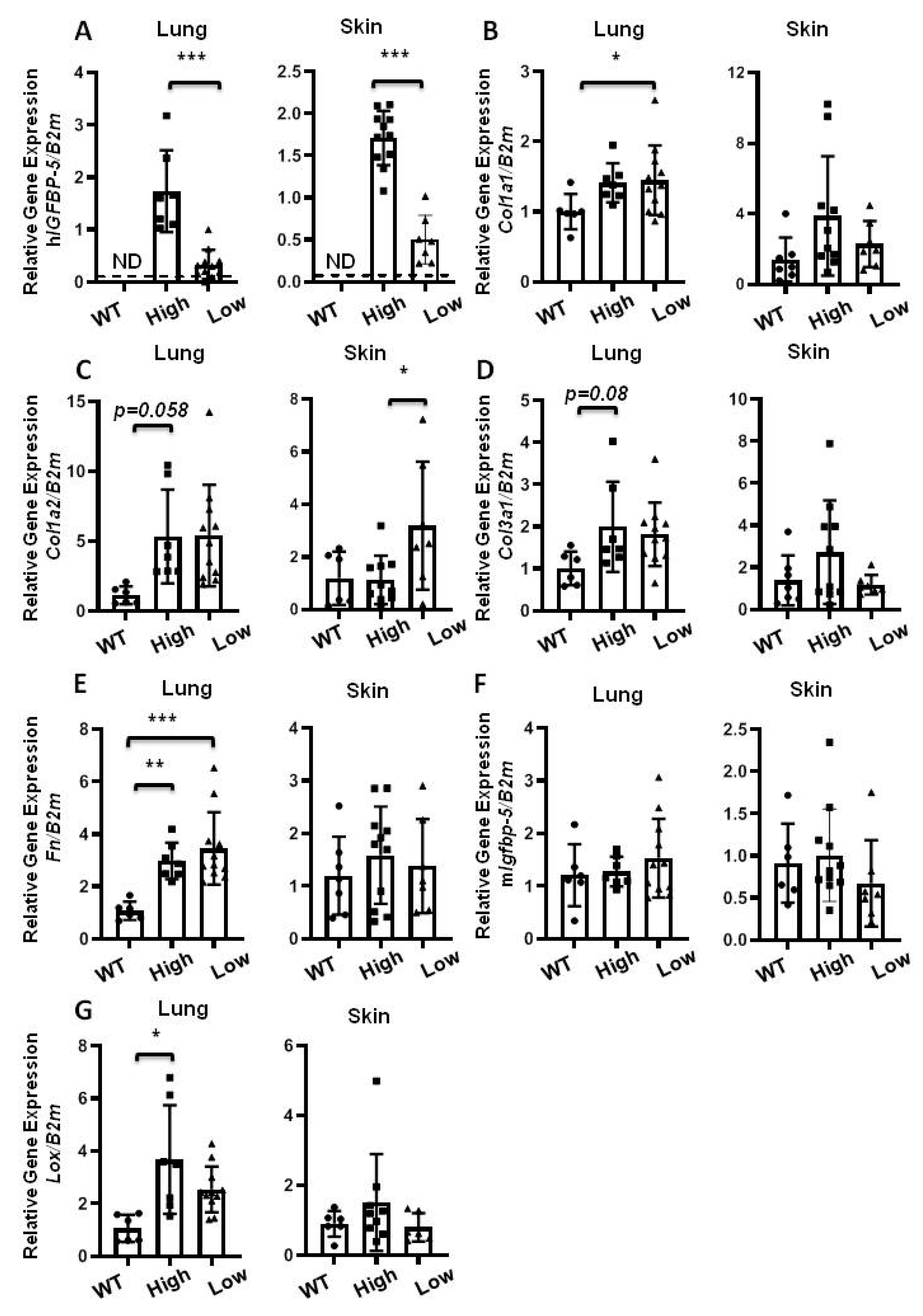
2.6. The Effect of Transgene Expression in Aging Mice
2.7. The Effect Transgene Expression on Bleomycin-Induced Pulmonary Fibrosis
2.8. The Effect of hIGFBP-5 Transgene Expression on the Phenotype of Lung Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Generation of Transgenic Mice Overexpressing Human IGFBP-5
4.3. Mouse Strains and Husbandry
4.4. Mouse Model for Oral Bleomycin (BLM) Delivery
4.5. Mouse Specimen Collection
4.6. Identification of Mouse Genotype
4.6.1. Genomic DNA Isolation from Mouse Tail Biopsy
4.6.2. Polymerase Chain Reaction
4.7. Histological Analysis
4.8. Culture of Primary Mouse Lung Fibroblasts
4.9. Quantitative Real-Time RT-PCR
4.10. Western Blot
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| α-SMA | Alpha-smooth muscle actin |
| B2m | Beta-2-microglobulin |
| BLM | Bleomycin |
| Col1a1 | Collagen type 1α1 |
| Col1a2 | Collagen type 1α2 |
| Col3a1 | Collagen type 3α1 |
| CTGF | Connective tissue growth factor |
| ECM | Extracellular matrix |
| EF1α | Elongation factor 1 alpha |
| Fn | Fibronectin |
| H&E | Hematoxylin and eosin |
| HET | Heterozygous |
| HOMO | Homozygous |
| hIGFBP-5 | Human IGFBP-5 |
| IGF-1, 2 | Insulin growth factor-1, 2 |
| IGFBP-5 | Insulin-like factor binding protein-5 |
| IPF | Idiopathic pulmonary fibrosis |
| Il-6 | Interleukin-6 |
| Lox | Lysyl oxidase |
| mIgfbp-5 | Mouse Igfbp-5 |
| PBS | Phosphate-buffered saline |
| PCNA | Proliferating cell nuclear antigen |
| SSc | Systemic sclerosis |
| WT | Wild-type |
References
- Varga, J.; Abraham, D. Systemic sclerosis: A prototypic multisystem fibrotic disorder. J. Clin. Investig. 2007, 117, 557–567. [Google Scholar] [CrossRef]
- Fan, M.H.; Feghali-Bostwick, C.A.; Silver, R.M. Update on scleroderma-associated interstitial lung disease. Curr. Opin. Rheumatol. 2014, 26, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Hsu, E.; Shi, H.; Jordan, R.M.; Lyons-Weiler, J.; Pilewski, J.M.; Feghali-Bostwick, C.A. Lung tissues in patients with systemic sclerosis have gene expression patterns unique to pulmonary fibrosis and pulmonary hypertension. Arthritis Rheum. 2011, 63, 783–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, E.L.; Mathur, A.; Tager, A.M.; Feghali-Bostwick, C.; Schneider, F.; Varga, J. Review: Interstitial lung disease associated with systemic sclerosis and idiopathic pulmonary fibrosis: How similar and distinct? Arthritis Rheumatol. 2014, 66, 1967–1978. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Nishimoto, T.; Feghali-Bostwick, C. IGFBP-5 Promotes Fibrosis Independently of Its Translocation to the Nucleus and Its Interaction with Nucleolin and IGF. PLoS ONE 2015, 10, e0130546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotarkonda, L.K.; Kulshrestha, R.; Ravi, K. Role of insulin like growth factor axis in the bleomycin induced lung injury in rats. Exp. Mol. Pathol. 2017, 102, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Pilewski, J.M.; Liu, L.; Henry, A.C.; Knauer, A.V.; Feghali-Bostwick, C.A. Insulin-like growth factor binding proteins 3 and 5 are overexpressed in idiopathic pulmonary fibrosis and contribute to extracellular matrix deposition. Am. J. Pathol. 2005, 166, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Yasuoka, H.; Yamaguchi, Y.; Feghali-Bostwick, C.A. The pro-fibrotic factor IGFBP-5 induces lung fibroblast and mononuclear cell migration. Am. J. Respir. Cell Mol. Biol. 2009, 41, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, X.X.; Muhammad, L.; Nietert, P.J.; Feghali-Bostwick, C. IGFBP-5 Promotes Fibrosis via Increasing Its Own Expression and That of Other Pro-fibrotic Mediators. Front. Endocrinol. (Lausanne) 2018, 9, 601. [Google Scholar] [CrossRef]
- Yasuoka, H.; Jukic, D.M.; Zhou, Z.; Choi, A.M.; Feghali-Bostwick, C.A. Insulin-like growth factor binding protein 5 induces skin fibrosis: A novel murine model for dermal fibrosis. Arthritis Rheum. 2006, 54, 3001–3010. [Google Scholar] [CrossRef]
- Yasuoka, H.; Zhou, Z.; Pilewski, J.M.; Oury, T.D.; Choi, A.M.; Feghali-Bostwick, C.A. Insulin-like growth factor-binding protein-5 induces pulmonary fibrosis and triggers mononuclear cellular infiltration. Am. J. Pathol. 2006, 169, 1633–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuoka, H.; Larregina, A.T.; Yamaguchi, Y.; Feghali-Bostwick, C.A. Human skin culture as an ex vivo model for assessing the fibrotic effects of insulin-like growth factor binding proteins. Open Rheumatol. J. 2008, 2, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, V.T.; Weber, T.; Graf, R.; Sommermann, T.; Petsch, K.; Sack, U.; Volchkov, P.; Rajewsky, K.; Kuhn, R. Efficient generation of Rosa26 knock-in mice using CRISPR/Cas9 in C57BL/6 zygotes. BMC Biotechnol. 2016, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, S.; Nagata, S. pEF-BOS, a powerful mammalian expression vector. Nucleic Acids Res. 1990, 18, 5322. [Google Scholar] [CrossRef] [PubMed]
- Hayward, M.D.; Jones, B.K.; Saparov, A.; Hain, H.S.; Trillat, A.C.; Bunzel, M.M.; Corona, A.; Li-Wang, B.; Strenkowski, B.; Giordano, C.; et al. An extensive phenotypic characterization of the hTNFalpha transgenic mice. BMC Physiol. 2007, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feghali, C.A.; Bost, K.L.; Boulware, D.W.; Levy, L.S. Mechanisms of pathogenesis in scleroderma. I. Overproduction of interleukin 6 by fibroblasts cultured from affected skin sites of patients with scleroderma. J. Rheumatol. 1992, 19, 1207–1211. [Google Scholar] [PubMed]
- Feghali, C.A.; Bost, K.L.; Boulware, D.W.; Levy, L.S. Control of IL-6 expression and response in fibroblasts from patients with systemic sclerosis. Autoimmunity 1994, 17, 309–318. [Google Scholar] [CrossRef]
- Redente, E.F.; Jacobsen, K.M.; Solomon, J.J.; Lara, A.R.; Faubel, S.; Keith, R.C.; Henson, P.M.; Downey, G.P.; Riches, D.W. Age and sex dimorphisms contribute to the severity of bleomycin-induced lung injury and fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L510–L518. [Google Scholar] [CrossRef]
- Watanabe, T.; Nishimoto, T.; Mlakar, L.; Heywood, J.; Malaab, M.; Hoffman, S.; Feghali-Bostwick, C. Optimization of a murine and human tissue model to recapitulate dermal and pulmonary features of systemic sclerosis. PLoS ONE 2017, 12, e0179917. [Google Scholar] [CrossRef] [Green Version]
- Feghali, C.A.; Wright, T.M. Identification of multiple, differentially expressed messenger RNAs in dermal fibroblasts from patients with systemic sclerosis. Arthritis Rheum. 1999, 42, 1451–1457. [Google Scholar] [CrossRef]
- Van Keuren, M.L.; Gavrilina, G.B.; Filipiak, W.E.; Zeidler, M.G.; Saunders, T.L. Generating transgenic mice from bacterial artificial chromosomes: Transgenesis efficiency, integration and expression outcomes. Transgenic. Res. 2009, 18, 769–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Clarke, J.B.; Busby, W.H.; Duan, C.; Clemmons, D.R. Insulin-like growth factor-binding protein-5 is cleaved by physiological concentrations of thrombin. Endocrinology 1998, 139, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Busby, W.H., Jr.; Nam, T.J.; Moralez, A.; Smith, C.; Jennings, M.; Clemmons, D.R. The complement component C1s is the protease that accounts for cleavage of insulin-like growth factor-binding protein-5 in fibroblast medium. J. Biol. Chem. 2000, 275, 37638–37644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overgaard, M.T.; Boldt, H.B.; Laursen, L.S.; Sottrup-Jensen, L.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A2 (PAPP-A2), a novel insulin-like growth factor-binding protein-5 proteinase. J. Biol. Chem. 2001, 276, 21849–21853. [Google Scholar] [CrossRef] [Green Version]
- Laursen, L.S.; Overgaard, M.T.; Soe, R.; Boldt, H.B.; Sottrup-Jensen, L.; Giudice, L.C.; Conover, C.A.; Oxvig, C. Pregnancy-associated plasma protein-A (PAPP-A) cleaves insulin-like growth factor binding protein (IGFBP)-5 independent of IGF: Implications for the mechanism of IGFBP-4 proteolysis by PAPP-A. FEBS Lett. 2001, 504, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.; Thompson, G.R.; Amaar, Y.G.; Hathaway, G.; Tschesche, H.; Baylink, D.J. ADAM-9 is an insulin-like growth factor binding protein-5 protease produced and secreted by human osteoblasts. Biochemistry 2002, 41, 15394–15403. [Google Scholar] [CrossRef]
- Loechel, F.; Fox, J.W.; Murphy, G.; Albrechtsen, R.; Wewer, U.M. ADAM 12-S cleaves IGFBP-3 and IGFBP-5 and is inhibited by TIMP-3. Biochem. Biophys. Res. Commun. 2000, 278, 511–515. [Google Scholar] [CrossRef]
- Cortesio, C.L.; Jiang, W. Mannan-binding lectin-associated serine protease 3 cleaves synthetic peptides and insulin-like growth factor-binding protein 5. Arch. Biochem. Biophys. 2006, 449, 164–170. [Google Scholar] [CrossRef]
- Gibson, T.L.; Cohen, P. Inflammation-related neutrophil proteases, cathepsin G and elastase, function as insulin-like growth factor binding protein proteases. Growth Horm. IGF Res. 1999, 9, 241–253. [Google Scholar] [CrossRef]
- Thrailkill, K.M.; Quarles, L.D.; Nagase, H.; Suzuki, K.; Serra, D.M.; Fowlkes, J.L. Characterization of insulin-like growth factor-binding protein 5-degrading proteases produced throughout murine osteoblast differentiation. Endocrinology 1995, 136, 3527–3533. [Google Scholar] [CrossRef]
- Campbell, P.G.; Andress, D.L. Plasmin degradation of insulin-like growth factor-binding protein-5 (IGFBP-5): Regulation by IGFBP-5-(201-218). Am. J. Physiol. 1997, 273, E996–E1004. [Google Scholar] [CrossRef] [PubMed]
- Rehault, S.; Monget, P.; Mazerbourg, S.; Tremblay, R.; Gutman, N.; Gauthier, F.; Moreau, T. Insulin-like growth factor binding proteins (IGFBPs) as potential physiological substrates for human kallikreins hK2 and hK3. Eur. J. Biochem. 2001, 268, 2960–2968. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Clemmons, D.R.; Smeekens, S. Expression and characterization of a serine protease that preferentially cleaves insulin-like growth factor binding protein-5. J. Cell. Biochem. 2005, 94, 470–484. [Google Scholar] [CrossRef] [PubMed]
- Garrett, S.M.; Baker Frost, D.; Feghali-Bostwick, C. The mighty fibroblast and its utility in scleroderma research. J. Scleroderma Relat. Disord. 2017, 2, 69–134. [Google Scholar] [CrossRef] [PubMed]
- Wolf, E.; Schneider, M.R.; Zhou, R.; Fisch, T.M.; Herbach, N.; Dahlhoff, M.; Wanke, R.; Hoeflich, A. Functional consequences of IGFBP excess-lessons from transgenic mice. Pediatr. Nephrol. 2005, 20, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.D.; Du, Z.; Buccilli, V.; Jorgetti, V.; Canalis, E. Transgenic mice overexpressing insulin-like growth factor binding protein-5 display transiently decreased osteoblastic function and osteopenia. Endocrinology 2002, 143, 3955–3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonner, E.; Barber, M.C.; Allan, G.J.; Beattie, J.; Webster, J.; Whitelaw, C.B.; Flint, D.J. Insulin-like growth factor binding protein-5 (IGFBP-5) induces premature cell death in the mammary glands of transgenic mice. Development 2002, 129, 4547–4557. [Google Scholar]
- Stout-Delgado, H.W.; Cho, S.J.; Chu, S.G.; Mitzel, D.N.; Villalba, J.; El-Chemaly, S.; Ryter, S.W.; Choi, A.M.; Rosas, I.O. Age-Dependent Susceptibility to Pulmonary Fibrosis Is Associated with NLRP3 Inflammasome Activation. Am. J. Respir. Cell. Mol. Biol. 2016, 55, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Thannickal, V.J. Mechanistic links between aging and lung fibrosis. Biogerontology 2013, 14, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Murtha, L.A.; Morten, M.; Schuliga, M.J.; Mabotuwana, N.S.; Hardy, S.A.; Waters, D.W.; Burgess, J.K.; Ngo, D.T.; Sverdlov, A.L.; Knight, D.A.; et al. The Role of Pathological Aging in Cardiac and Pulmonary Fibrosis. Aging Dis. 2019, 10, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Schafer, M.J.; Miller, J.D.; LeBrasseur, N.K. Cellular senescence: Implications for metabolic disease. Mol. Cell Endocrinol. 2017, 455, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Rodier, F.; Campisi, J. Four faces of cellular senescence. J. Cell. Biol. 2011, 192, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Krizhanovsky, V.; Yon, M.; Dickins, R.A.; Hearn, S.; Simon, J.; Miething, C.; Yee, H.; Zender, L.; Lowe, S.W. Senescence of activated stellate cells limits liver fibrosis. Cell 2008, 134, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voltz, J.W.; Card, J.W.; Carey, M.A.; Degraff, L.M.; Ferguson, C.D.; Flake, G.P.; Bonner, J.C.; Korach, K.S.; Zeldin, D.C. Male sex hormones exacerbate lung function impairment after bleomycin-induced pulmonary fibrosis. Am. J. Respir. Cell. Mol. Biol. 2008, 39, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Steen, V.D.; Medsger, T.A. Changes in causes of death in systemic sclerosis, 1972–2002. Ann. Rheum. Dis. 2007, 66, 940–944. [Google Scholar] [CrossRef] [Green Version]
- Pintar, J.E.; Cerro, J.A.; Wood, T.L. Genetic approaches to the function of insulin-like growth factor-binding proteins during rodent development. Horm. Res. 1996, 45, 172–177. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer 1 | 5′-AAGCACGTTTCCGACTTGAGTTG-3′ |
| Reverse Primer 1 | 5′-GGGTGAGCATGTCTTTAATCTACC-3′ |
| Reverse Primer 2 | 5′-GAGCCAGTACACGACATCACTTTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, X.-X.; Sanderson, M.; Helke, K.; Feghali-Bostwick, C. Phenotypic Characterization of Transgenic Mice Expressing Human IGFBP-5. Int. J. Mol. Sci. 2021, 22, 335. https://doi.org/10.3390/ijms22010335
Nguyen X-X, Sanderson M, Helke K, Feghali-Bostwick C. Phenotypic Characterization of Transgenic Mice Expressing Human IGFBP-5. International Journal of Molecular Sciences. 2021; 22(1):335. https://doi.org/10.3390/ijms22010335
Chicago/Turabian StyleNguyen, Xinh-Xinh, Matthew Sanderson, Kristi Helke, and Carol Feghali-Bostwick. 2021. "Phenotypic Characterization of Transgenic Mice Expressing Human IGFBP-5" International Journal of Molecular Sciences 22, no. 1: 335. https://doi.org/10.3390/ijms22010335
APA StyleNguyen, X. -X., Sanderson, M., Helke, K., & Feghali-Bostwick, C. (2021). Phenotypic Characterization of Transgenic Mice Expressing Human IGFBP-5. International Journal of Molecular Sciences, 22(1), 335. https://doi.org/10.3390/ijms22010335