Effects of Gut Metabolites and Microbiota in Healthy and Marginal Livers Submitted to Surgery

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
Relevance of the Gut–Liver Axis
2. Relationship between Liver Transplantation and Gut Microbiota
3. Relationship between Partial Hepatectomy and Gut Microbiota
4. Gut Microbiota-Based Therapy
4.1. Pro/Prebiotics
4.2. Antibiotics
4.3. Diet and Components that Modulate the Gut Microbiota
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAs | Amino acids |
| ACR | Acute-cell rejection |
| ALD | Alcoholic liver disease |
| ALT | Alanine transaminase |
| Amp | Ampicillin |
| AR | Acute rejection |
| AST | Aspartate transaminase |
| Atb | Antibody |
| BA | Bile acid |
| BCAA | Branched-chain amino acid |
| BD | Brain-dead |
| BN | Brown Norway |
| BrdU | Bromodeoxyuridine |
| BT | Bacterial translocation |
| CD | Cardiac death |
| CNIs | Calcineurin inhibitors |
| CVD | Cardiovascular disease |
| CYP7A1 | Cytochrome P450 7A1 |
| DCA | Deoxycholic acid |
| DCD | Donation after circulatory death |
| ESLD | End-stage liver disease |
| FGF15 | Fibroblast growth factor 15 |
| FGFR4-KLB | Fibroblast growth factor receptor-4 and its co-receptor klotho-beta |
| FMT | Fecal microbial transplantation |
| FXR | Farnesoid X receptor |
| GC | Glucocorticoids |
| GLP-1 | Glucagon-like peptide 1 |
| GM | Gut microbiota |
| Gln | Glutamine |
| HCC | Hepatocellular carcinoma |
| HGF | Hepatocyte growth factor |
| HWP | Hydrolyzed whey peptide |
| I/R | Ischemia/reperfusion |
| ICAM | Intercellular adhesion molecule |
| ICU | Intensive care unit |
| IL | Interleukin |
| IL-1β | Interleukin-1 β |
| IL-6 | Interleukin-6 |
| IL-10 | Interleukin-10 |
| IL-12 | Interleukin-12 |
| IRI | Ischemia/reperfusion injury |
| IS | Immunosuppression |
| Ki67 | Marker of proliferation Ki-67 |
| L | Lewis |
| LCA | Lithocholic acid |
| LD | Linear dichroism |
| LN | Liver transplantation in normal receptors |
| LP-F19 | Lactobacillus paracasei F19 |
| LPS | Lipopolysaccharides |
| LR | Liver regeneration |
| LT | Liver transplantation |
| LTA | Abnormal liver function |
| LTC | Liver transplantation in cirrhotic receptors |
| LTN | Liver transplantation in normal receptors |
| LTN | Normal liver function |
| LTRs | Liver-transplant recipients |
| MDA | Malondialdehyde |
| Met | Metronidazole |
| MP | Methylprednisolone |
| MPA | Mycophenolate |
| Muc2 | Mucin 2 |
| Muc3 | Mucin 3 |
| NAFLD | Non-alcoholic fatty liver disease |
| NASH | Non-alcoholic steatohepatitis |
| Neo | Neomycin sulfate |
| NFκB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NKT cell | Natural killer-T cell |
| NLRs | Nod-like receptors |
| NR | Non-rejection group |
| OLT | Orthotropic liver transplant |
| PCNA | Proliferating cell nuclear antigen |
| PGE2 | Prostaglandin E2 |
| PH | Partial hepatectomy |
| RA | All-trans retinoic acid |
| ROS | Reactive oxygen species |
| R TGR5 | Receptor of Takeda G protein-coupled receptor 5 |
| SCFAs | Short-chain fatty acids |
| SD | Sprague Dawley |
| sIgA | Secretory IgA |
| SOD | Superoxide dismutase |
| TGF-β | Transforming growth factor β |
| TGR5 | Takeda G protein-coupled receptor 5 |
| TJ | Tight junction |
| TLR | Toll-like receptor |
| TLR4 | Toll-like receptor 4 |
| TMP-SMX | Trimethoprim-sulfamethoxazole |
| TNF | Tumor necrosis factor |
| TNFα | Tumor necrosis factor α |
| Treg | Regulatory T cells |
| TRF | Time-restricted feeding |
| Vanco | Vancomycin |
| W | Wistar |
References
- Álvarez-Mercado, A.I.; Gulfo, J.; Romero Gómez, M.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. Use of Steatotic Grafts in Liver Transplantation: Current Status. Liver Transplant. 2019, 25, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Cillo, U.; Bechstein, W.O.; Berlakovich, G.; Dutkowski, P.; Lehner, F.; Nadalin, S.; Saliba, F.; Schlitt, H.J.; Pratschke, J. Identifying risk profiles in liver transplant candidates and implications for induction immunosuppression. Transplant. Rev. 2018, 32, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Núñez, K.; Thevenot, P.; Alfadhli, A.; Cohen, A. Complement activation in liver transplantation: Role of donor macrosteatosis and implications in delayed graft function. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornide-Petronio, M.E.; Álvarez-Mercado, A.I.; Jiménez-Castro, M.B.; Peralta, C. Current Knowledge about the Effect of Nutritional Status, Supplemented Nutrition Diet, and Gut Microbiota on Hepatic Ischemia-Reperfusion and Regeneration in Liver Surgery. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralta, C.; Jiménez-Castro, M.B.; Gracia-Sancho, J. Hepatic ischemia and reperfusion injury: Effects on the liver sinusoidal milieu. J. Hepatol. 2013, 59, 1094–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydari, M.; Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Peralta, C. Data on adiponectin from 2010 to 2020: Therapeutic target and prognostic factor for liver diseases? Int. J. Mol. Sci. 2020, 21, 1–24. [Google Scholar] [CrossRef]
- Miyauchi, T.; Uchida, Y.; Kadono, K.; Hirao, H.; Kawasoe, J.; Watanabe, T.; Ueda, S.; Jobara, K.; Kaido, T.; Okajima, H.; et al. Preventive Effect of Antioxidative Nutrient-Rich Enteral Diet Against Liver Ischemia and Reperfusion Injury. J. Parenter. Enter. Nutr. 2019, 43, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Monson, K.M.; Dowlatshahi, S.; Crockett, E.T. CXC-chemokine regulation and neutrophil trafficking in hepatic ischemia-reperfusion injury in P-selectin/ICAM-1 deficient mice. J. Inflamm. 2007, 4. [Google Scholar] [CrossRef] [Green Version]
- Peralta, C.; Fernández, L.; Panés, J.; Prats, N.; Sans, M.; Piqué, J.M.; Gelpí, E.; Roselló-Catafau, J. Preconditioning protects against systemic disorders associated with hepatic ischemia-reperfusion through blockade of tumor necrosis factor-induced P-selectin up-regulation in the rat. Hepatology 2001, 33, 100–113. [Google Scholar] [CrossRef]
- Yang, H.J.; Tang, L.M.; Zhou, X.J.; Qian, J.; Zhu, J.; Lu, L.; Wang, X.H. Ankaflavin ameliorates steatotic liver ischemia-reperfusion injury in mice. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 619–625. [Google Scholar] [CrossRef]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriss, M.; Verna, E.C.; Rosen, H.R.; Lozupone, C.A. Functional Microbiomics in Liver Transplantation: Identifying Novel Targets for Improving Allograft Outcomes. Transplantation 2019, 103, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Rocha, C.S.; Dandekar, S.; Yvonne Wan, Y.J. Functional analysis of the relationship between intestinal microbiota and the expression of hepatic genes and pathways during the course of liver regeneration. J. Hepatol. 2016, 64, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Q.; Yu, L.; Chen, D.; Li, L. Variation in the gut microbial community is associated with the progression of liver regeneration. Hepatol. Res. 2020, 50, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, J.S.; Kakiyama, G.; Cox, I.J.; Nittono, H.; Takei, H.; White, M.; Fagan, A.; Gavis, E.A.; Heuman, D.M.; Gilles, H.C.; et al. Alterations in gut microbial function following liver transplant. Liver Transplant. 2018, 24, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Meroni, M.; Longo, M.; Dongiovanni, P. Alcohol or Gut Microbiota: Who Is the Guilty? Int. J. Mol. Sci. 2019, 20, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Milosevic, I.; Vujovic, A.; Barac, A.; Djelic, M.; Korac, M.; Spurnic, A.R.; Gmizic, I.; Stevanovic, O.; Djordjevic, V.; Lekic, N.; et al. Gut-liver axis, gut microbiota, and its modulation in the management of liver diseases: A review of the literature. Int. J. Mol. Sci. 2019, 20, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Lu, P.C.; Lo, M.H.; Lin, I.C.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Gut microbiota-dependent trimethylamine N-oxide pathway associated with cardiovascular risk in children with early-stage chronic kidney disease. Int. J. Mol. Sci. 2018, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, H.; Enomoto, H.; Nishiguchi, S.; Iijima, H. Liver cirrhosis and sarcopenia from the viewpoint of dysbiosis. Int. J. Mol. Sci. 2020, 21, 1–24. [Google Scholar] [CrossRef]
- Dumas, M.E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511–12516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Yang, L.; Chin, Y.; Liu, F.; Li, R.W.; Yuan, S.; Xue, C.; Xu, J.; Tang, Q. Astaxanthin n-octanoic acid diester ameliorates insulin resistance and modulates gut microbiota in high-fat and high-sucrose diet-fed mice. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.; Harsch, I.; Konturek, K.; Schink, M.; Konturek, T.; Neurath, M.; Zopf, Y. Gut–Liver Axis: How Do Gut Bacteria Influence the Liver? Med. Sci. 2018, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- Poeta, M.; Pierri, L.; Vajro, P. Gut–Liver Axis Derangement in Non-Alcoholic Fatty Liver Disease. Children 2017, 4, 66. [Google Scholar] [CrossRef] [Green Version]
- Adolph, T.E.; Grander, C.; Moschen, A.R.; Tilg, H. Liver–Microbiome Axis in Health and Disease. Trends Immunol. 2018, 39, 712–723. [Google Scholar] [CrossRef]
- Brun, P.; Castagliuolo, I.; Di Leo, V.; Buda, A.; Pinzani, M.; Palù, G.; Martines, D. Increased intestinal permeability in obese mice: New evidence in the pathogenesis of nonalcoholic steatohepatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Harte, A.L.; Da Silva, N.F.; Creely, S.J.; McGee, K.C.; Billyard, T.; Youssef-Elabd, E.M.; Tripathi, G.; Ashour, E.; Abdalla, M.S.; Sharada, H.M.; et al. Elevated endotoxin levels in non-alcoholic fatty liver disease. J. Inflamm. 2010, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhu, C.; Shao, L.; Ye, J.; Shen, Y.; Ren, Y. Role of bile acids in dysbiosis and treatment of nonalcoholic fatty liver disease. Mediators Inflamm. 2019, 2019. [Google Scholar] [CrossRef]
- Benten, D.; Wiest, R. Gut microbiome and intestinal barrier failure-The “achilles heel” in hepatology? J. Hepatol. 2012, 56, 1221–1223. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ji, F.; Guo, J.; Shi, D.; Fang, D.; Li, L. Dysbiosis of small intestinal microbiota in liver cirrhosis and its association with etiology. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Minicis, S.; Rychlicki, C.; Agostinelli, L.; Saccomanno, S.; Candelaresi, C.; Trozzi, L.; Mingarelli, E.; Facinelli, B.; Magi, G.; Palmieri, C.; et al. Dysbiosis contributes to fibrogenesis in the course of chronic liver injury in mice. Hepatology 2014, 59, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Douhara, A.; Moriya, K.; Yoshiji, H.; Noguchi, R.; Namisaki, T.; Kitade, M.; Kaji, K.; Aihara, Y.; Nishimura, N.; Takeda, K.; et al. Reduction of endotoxin attenuates liver fibrosis through suppression of hepatic stellate cell activation and remission of intestinal permeability in a rat non-alcoholic steatohepatitis model. Mol. Med. Rep. 2015, 11, 1693–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccioni, L.; Gao, B.; Leclercq, S.; Pirlot, B.; Horsmans, Y.; De Timary, P.; Leclercq, I.; Fouts, D.; Schnabl, B.; Stärkel, P. Intestinal permeability, microbial translocation, changes in duodenal and fecal microbiota, and their associations with alcoholic liver disease progression in humans. Gut Microbes 2020, 12. [Google Scholar] [CrossRef]
- Malaguarnera, G.; Giordano, M.; Nunnari, G.; Bertino, G.; Malaguarnera, M. Gut microbiota in alcoholic liver disease: Pathogenetic role and therapeutic perspectives. World J. Gastroenterol. 2014, 20, 16639–16648. [Google Scholar] [CrossRef]
- Seo, B.; Jeon, K.; Moon, S.; Lee, K.; Kim, W.K.; Jeong, H.; Cha, K.H.; Lim, M.Y.; Kang, W.; Kweon, M.N.; et al. Roseburia spp. Abundance Associates with Alcohol Consumption in Humans and Its Administration Ameliorates Alcoholic Fatty Liver in Mice. Cell Host Microbe 2020, 27, 25–40.e6. [Google Scholar] [CrossRef]
- Chen, L.; van den Munckhof, I.C.L.; Schraa, K.; ter Horst, R.; Koehorst, M.; van Faassen, M.; van der Ley, C.; Doestzada, M.; Zhernakova, D.V.; Kurilshikov, A.; et al. Genetic and Microbial Associations to Plasma and Fecal Bile Acids in Obesity Relate to Plasma Lipids and Liver Fat Content. Cell Rep. 2020, 33, 108212. [Google Scholar] [CrossRef]
- Udayappan, S.; Manneras-Holm, L.; Chaplin-Scott, A.; Belzer, C.; Herrema, H.; Dallinga-Thie, G.M.; Duncan, S.H.; Stroes, E.S.G.; Groen, A.K.; Flint, H.J.; et al. Oral treatment with Eubacterium hallii improves insulin sensitivity in db/db mice. NPJ Biofilms Microb. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Tonin, F.; Arends, I.W.C.E. Latest development in the synthesis of ursodeoxycholic acid (UDCA): A critical review. Beilstein J. Org. Chem. 2018, 14, 470–483. [Google Scholar] [CrossRef] [Green Version]
- Kurilshikov, A.; Van Den Munckhof, I.C.L.; Chen, L.; Bonder, M.J.; Schraa, K.; Rutten, J.H.W.; Riksen, N.P.; De Graaf, J.; Oosting, M.; Sanna, S.; et al. Gut Microbial Associations to Plasma Metabolites Linked to Cardiovascular Phenotypes and Risk: A Cross-Sectional Study. Circ. Res. 2019, 124, 1808–1820. [Google Scholar] [CrossRef]
- Munukka, E.; Rintala, A.; Toivonen, R.; Nylund, M.; Yang, B.; Takanen, A.; Hänninen, A.; Vuopio, J.; Huovinen, P.; Jalkanen, S.; et al. Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J. 2017, 11, 1667–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.F.; Ren, Z.G.; Li, A.; Zhang, H.; Xu, S.Y.; Jiang, J.W.; Zhou, L.; Ling, Q.; Wang, B.H.; Cui, G.Y.; et al. Fecal microbiome data distinguish liver recipients with normal and abnormal liver function from healthy controls. Front. Microbiol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ma, L.; Shen, S.; Guo, Y.; Cao, Q.; Cai, X.; Feng, J.; Yan, Y.; Hu, T.; Luo, S.; et al. Intestinal dysbacteriosis-induced IL-25 promotes development of HCC via alternative activation of macrophages in tumor microenvironment. J. Exp. Clin. Cancer Res. 2019, 38, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lukovic, E.; Moitra, V.K.; Freedberg, D.E. The microbiome: Implications for perioperative and critical care. Curr. Opin. Anaesthesiol. 2019, 32, 412–420. [Google Scholar] [CrossRef]
- Kageyama, S.; Kadono, K.; Hirao, H.; Nakamura, K.; Ito, T.; Gjertson, D.W.; Sosa, R.A.; Reed, E.F.; Kaldas, F.M.; Busuttil, R.W.; et al. Ischemia-Reperfusion Injury in Allogeneic Liver Transplantation. Transplantation 2020. [Google Scholar] [CrossRef]
- Charlton, M.; Levitsky, J.; Aqel, B.; O’Grady, J.; Hemibach, J.; Rinella, M.; Fung, J.; Ghabril, M.; Thomason, R.; Burra, P.; et al. International Liver Transplantation Society Consensus Statement on Immunosuppression in Liver Transplant Recipients. Transplantation 2018, 102, 727–743. [Google Scholar] [CrossRef]
- Yoon, Y.I.; Song, G.W.; Lee, S.G.; Hwang, S.; Kim, K.H.; Kim, S.H.; Kang, W.H.; Cho, H.D.; Jwa, E.K.; Kwon, J.H.; et al. Outcome of ABO-incompatible adult living-donor liver transplantation for patients with hepatocellular carcinoma. J. Hepatol. 2018, 68, 1153–1162. [Google Scholar] [CrossRef]
- Schubert, K.; Olde Damink, S.W.M.; von Bergen, M.; Schaap, F.G. Interactions between bile salts, gut microbiota, and hepatic innate immunity. Immunol. Rev. 2017, 279, 23–35. [Google Scholar] [CrossRef]
- Parséus, A.; Sommer, N.; Sommer, F.; Caesar, R.; Molinaro, A.; Stahlman, M.; Greiner, T.U.; Perkins, R.; Bäckhed, F. Microbiota-induced obesity requires farnesoid X receptor. Gut 2017, 66, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Sinal, C.J.; Tohkin, M.; Miyata, M.; Ward, J.M.; Lambert, G.; Gonzalez, F.J. Targeted disruption of the nuclear receptor FXR/BAR impairs bile acid and lipid homeostasis. Cell 2000, 102, 731–744. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-Chain fatty acids suppress lipopolysaccharide-Induced production of nitric oxide and proinflammatory cytokines through inhibition of NF-κB Pathway in RAW264.7 cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Suriguga, S.; Luangmonkong, T.; Mutsaers, H.A.M.; Groothuis, G.M.M.; Olinga, P. Host microbiota dictates the proinflammatory impact of LPS in the murine liver. Toxicol. Vitr. 2020, 67, 104920. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Gracia-Sancho, J.; Peralta, C. Inflammasome-Mediated Inflammation in Liver Ischemia-Reperfusion Injury. Cells 2019, 8, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, J.; Mulholland, M.; Zhang, W. mTOR activation protects liver from ischemia/reperfusion-induced injury through NF-κB pathway. FASEB J. 2017, 31, 3018–3026. [Google Scholar] [CrossRef]
- Kim, C.H. Immune regulation by microbiome metabolites. Immunology 2018, 154, 220–229. [Google Scholar] [CrossRef]
- Ichikawa, R.; Takayama, T.; Yoneno, K.; Kamada, N.; Kitazume, M.T.; Higuchi, H.; Matsuoka, K.; Watanabe, M.; Itoh, H.; Kanai, T.; et al. Bile acids induce monocyte differentiation toward interleukin-12 hypo-producing dendritic cells via a TGR5-dependent pathway. Immunology 2012, 136, 153–162. [Google Scholar] [CrossRef]
- Chávez-talavera, O.; Tailleux, A.; Lefebvre, P.; Staels, B. Bile Acid Control of Metabolism and Inflammation in Obesity, Type 2 Diabetes, Dyslipidemia and NAFLD. Gastroenterology 2017, 152, 1679–1694. [Google Scholar] [CrossRef]
- Gulfo, J.; Rotondo, F.; Ávalos de León, C.G.; Cornide-Petronio, M.E.; Fuster, C.; Gracia-Sancho, J.; Jiménez-Castro, M.B.; Peralta, C. FGF15 improves outcomes after brain dead donor liver transplantation with steatotic and non-steatotic grafts in rats. J. Hepatol. 2020, 73, 1131–1143. [Google Scholar] [CrossRef]
- León, C.G.A.; Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Gulfo, J.; Rotondo, F.; Gracia-Sancho, J.; Casillas-Ramírez, A.; Peralta, C. The Effect of Fibroblast Growth Factor 15 Signaling in Non-Steatotic and Steatotic Liver Transplantation from Cardiocirculatory Death. Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Ducastel, S.; Touche, V.; Trabelsi, M.S.; Boulinguiez, A.; Butruille, L.; Nawrot, M.; Peschard, S.; Chávez-Talavera, O.; Dorchies, E.; Vallez, E.; et al. The nuclear receptor FXR inhibits Glucagon-Like Peptide-1 secretion in response to microbiota-derived Short-Chain Fatty Acids. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pathak, P.; Xie, C.; Nichols, R.G.; Ferrell, J.M.; Boehme, S.; Krausz, K.W.; Patterson, A.D.; Gonzalez, F.J.; Chiang, J.Y.L. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018, 68, 1574–1588. [Google Scholar] [CrossRef] [PubMed]
- Avalos-de León, C.G.; Jiménez-Castro, M.B.; Cornide-Petronio, M.E.; Casillas-Ramírez, A.; Peralta, C. The Role of GLP1 in Rat Steatotic and Non-Steatotic Liver Transplantation from Cardiocirculatory Death Donors. Cells 2019, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Chen, Y.; Xia, F.; Abudukerimu, B.; Zhang, W.; Guo, Y.; Wang, N.; Lu, Y. A glucagon-like peptide-1 receptor agonist lowers weight by modulating the structure of gut microbiota. Front. Endocrinol. (Lausanne) 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; van Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur. J. Pharmacol. 2018, 831, 52–59. [Google Scholar] [CrossRef]
- Ang, Z.; Er, J.Z.; Tan, N.S.; Lu, J.; Liou, Y.C.; Grosse, J.; Ding, J.L. Human and mouse monocytes display distinct signalling and cytokine profiles upon stimulation with FFAR2/FFAR3 short-chain fatty acid receptor agonists. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef]
- Qiao, Y.L.; Qian, J.M.; Wang, F.R.; Ma, Z.Y.; Wang, Q.W. Butyrate protects liver against ischemia reperfusion injury by inhibiting nuclear factor kappa B activation in Kupffer cells. J. Surg. Res. 2014, 187, 653–659. [Google Scholar] [CrossRef]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Asarat, M.; Apostolopoulos, V.; Vasiljevic, T.; Donkor, O. Short-chain fatty acids regulate cytokines and Th17/treg cells in human peripheral blood mononuclear cells in vitro. Immunol. Invest. 2016, 45, 205–222. [Google Scholar] [CrossRef] [Green Version]
- Clavien, P.A.; Muller, X.; de Oliveira, M.L.; Dutkowski, P.; Sanchez-Fueyo, A. Can immunosuppression be stopped after liver transplantation? Lancet Gastroenterol. Hepatol. 2017, 2, 531–537. [Google Scholar] [CrossRef]
- Wu, Z.W.; Ling, Z.X.; Lu, H.F.; Zuo, J.; Sheng, J.F.; Zheng, S.S.; Li, L.J. Changes of gut bacteria and immune parameters in liver transplant recipients. Hepatobiliary Pancreat. Dis. Int. 2012, 11, 40–50. [Google Scholar] [CrossRef]
- Buchman, A.L. Glutamine: Is it a conditionally required nutrient for the human gastrointestinal system? J. Am. Coll. Nutr. 1996, 15, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruzat, V.; Rogero, M.M.; Keane, K.N.; Curi, R.; Newsholme, P. Glutamine: Metabolism and immune function, supplementation and clinical translation. Nutrients 2018, 10, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Hammad, A.; Kaido, T.; Aliyev, V.; Mandato, C.; Uemoto, S. Nutritional therapy in liver transplantation. Nutrients 2017, 9, 1–25. [Google Scholar] [CrossRef]
- Mu, J.; Chen, Q.; Zhu, L.; Wu, Y.; Liu, S.; Zhao, Y.; Ma, T. Influence of gut microbiota and intestinal barrier on enterogenic infection after liver transplantation. Curr. Med. Res. Opin. 2019, 35, 241–248. [Google Scholar] [CrossRef]
- Portune, K.J.; Beaumont, M.; Davila, A.M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Echániz, A.; Pita, S.; Otero, A.; Suárez, F.; Gómez, M.; Guerrero, A. Incidencia, factores de riesgo e influencia sobre la supervivencia de las complicaciones infecciosas en el trasplante hepático. Enferm. Infecc. Microbiol. Clin. 2003, 21, 224–231. [Google Scholar] [CrossRef]
- Losada, I.; Cuervas-Mons, V.; Millán, I.; Dámaso, D. Early infection in liver transplant recipients: Incidence, severity, risk factors and antibiotic sensitivity of bacterial isolate. Enferm. Infecc. Microbiol. Clin. 2002, 20, 422–430. [Google Scholar] [CrossRef]
- Kato, K.; Nagao, M.; Miyamoto, K.; Oka, K.; Takahashi, M.; Yamamoto, M.; Matsumura, Y.; Kaido, T.; Uemoto, S.; Ichiyama, S. Longitudinal Analysis of the Intestinal Microbiota in Liver Transplantation. Transplant. Direct. 2017, 3, e144. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, A.; Ponziani, F.R.; Biolato, M.; Valenza, V.; Marrone, G.; Sganga, G.; Gasbarrini, A.; Miele, L.; Grieco, A. Intestinal permeability in the pathogenesis of liver damage: From non-alcoholic fatty liver disease to liver transplantation. World J. Gastroenterol. 2019, 25, 4814–4834. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.R.; Liu, S.L.; Liu, X.; Luo, Z.B.; Zhu, B.; Li, Z.F.; Li, L.J.; He, Y.; Jiang, L.; Li, H.; et al. Intestinal microbiota and innate immunity-related gene alteration in cirrhotic rats with liver transplantation. Transplant. Proc. 2011, 43, 3973–3979. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Luo, Z.; Li, Z.; Deng, M.; Liu, H.; Zhu, B.; Ruan, B.; Li, L. Structural Shifts of Fecal Microbial Communities in Rats with Acute Rejection after Liver Transplantation. Microb. Ecol. 2012, 64, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Jiang, J.; Lu, H.; Chen, X.; He, Y.; Zhang, H.; Xie, H.; Wang, W.; Zheng, S.; Zhou, L. Intestinal microbial variation may predict early acute rejection after liver transplantation in rats. Transplantation 2014, 98, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Chen, H.; Zhu, B.; Qin, N.; Chen, Y.; Li, Z.; Deng, M.; Jiang, H.; Xu, X.; Yang, J.; et al. Effect of Intestinal Microbiota Alteration on Hepatic Damage in Rats with Acute Rejection After Liver Transplantation. Microb. Ecol. 2014, 68, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Heredia, N.; Peralta, C.; Xaus, C.; Roselló-Catafau, J.; Rimola, A.; Marco, A.; Serafín, A.; Deulofeu, R.; Gelpí, E.; et al. Role of ischemic preconditioning and the portosystemic shunt in the prevention of liver and lung damage after rat liver transplantation. Transplantation 2003, 76, 282–289. [Google Scholar] [CrossRef]
- Mendes-Braz, M.; Elias-Miró, M.; Jiménez-Castro, M.B.; Casillas-Ramírez, A.; Ramalho, F.S.; Peralta, C. The current state of knowledge of hepatic ischemia-reperfusion injury based on its study in experimental models. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef]
- Casillas-Ramírez, A.; Mosbah, I.B.; Ramalho, F.; Roselló-Catafau, J.; Peralta, C. Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci. 2006, 79, 1881–1894. [Google Scholar] [CrossRef]
- Jiménez-Castro, M.B.; Meroño, N.; Mendes-Braz, M.; Gracia-Sancho, J.; Martínez-Carreres, L.; Cornide-Petronio, M.E.; Casillas-Ramirez, A.; Rodés, J.; Peralta, C. The effect of brain death in rat steatotic and non-steatotic liver transplantation with previous ischemic preconditioning. J. Hepatol. 2015, 62, 83–91. [Google Scholar] [CrossRef]
- Fernández, L.; Carrasco-Chaumel, E.; Serafín, A.; Xaus, C.; Grande, L.; Rimola, A.; Roselló-Catafau, J.; Peralta, C. Is ischemic preconditioning a useful strategy in steatotic liver transplantation? Am. J. Transplant. 2004, 4, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, A.; Scalera, I.; Perera, M.T.P.R.; Kalisvaart, M.; Mergental, H.; Mirza, D.F.; Isaac, J.; Muiesan, P. Impact of donor age in donation after circulatory death liver transplantation: Is the cutoff “60” still of relevance? Liver Transplant. 2018, 24, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Pan, E.T.; Yoeli, D.; Galvan, N.T.N.; Kueht, M.L.; Cotton, R.T.; O’Mahony, C.A.; Goss, J.A.; Rana, A. Cold ischemia time is an important risk factor for post–liver transplant prolonged length of stay. Liver Transplant. 2018, 24, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.G.; Lee, H.; Oh, S.Y.; Jung, C.W.; Ryu, H.G. Risk Factors for Intensive Care Unit Readmission After Liver Transplantation: A Retrospective Cohort Study. Ann. Transplant. 2018, 23, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; He, J.; Wu, Z.; Xu, W.; Zhang, H.; Ye, P.; Yang, J.; Zhen, S.; Li, L. Assessment of Microbiome Variation During the Perioperative Period in Liver Transplant Patients: A Retrospective Analysis. Microb. Ecol. 2013, 65, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Fagan, A.; Sikaroodi, M.; White, M.B.; Sterling, R.K.; Gilles, H.C.; Heuman, D.; Stravitz, R.T.; Matherly, S.C.; Siddiqui, M.S.; et al. Liver transplant modulates gut microbial dysbiosis and cognitive function in cirrhosis. Liver Transplant. 2017, 23, 907–914. [Google Scholar] [CrossRef]
- Sun, L.Y.; Yang, Y.S.; Qu, W.; Zhu, Z.J.; Wei, L.; Ye, Z.S.; Zhang, J.R.; Sun, X.Y.; Zeng, Z.G. Gut microbiota of liver transplantation recipients. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tourret, J.; Willing, B.P.; Dion, S.; MacPherson, J.; Denamur, E.; Finlay, B.B. Immunosuppressive treatment alters secretion of ileal antimicrobial peptides and gut microbiota, and favors subsequent colonization by uropathogenic Escherichia coli. Transplantation 2017, 101, 74–82. [Google Scholar] [CrossRef]
- Jia, J.; Tian, X.; Jiang, J.; Ren, Z.; Lu, H.; He, N.; Xie, H.; Zhou, L.; Zheng, S. Structural shifts in the intestinal microbiota of rats treated with cyclosporine A after orthotropic liver transplantation. Front. Med. 2019, 13, 451–460. [Google Scholar] [CrossRef]
- Jiang, J.W.; Ren, Z.G.; Lu, H.F.; Zhang, H.; Li, A.; Cui, G.Y.; Jia, J.J.; Xie, H.Y.; Chen, X.H.; He, Y.; et al. Optimal immunosuppressor induces stable gut microbiota after liver transplantation. World J. Gastroenterol. 2018, 24, 3871–3883. [Google Scholar] [CrossRef]
- Gabe, S.M.; Bjarnason, I.; Tolou-Ghamari, Z.; Tredger, J.M.; Johnson, P.G.; Barclay, G.R.; Williams, R.; Silk, D.B.A. The effect of tacrolimus (FK506) on intestinal barrier function and cellular energy production in humans. Gastroenterology 1998, 115, 67–74. [Google Scholar] [CrossRef]
- Parrilli, G.; Abazia, C.; Sarnelli, G.; Corsaro, M.M.; Coccoli, P.; Viglione, L.; Cuomo, R.; Budillon, G. Effect of chronic administration of tacrolimus and cyclosporine on human gastrointestinal permeability. Liver Transplant. 2003, 9, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Ladirat, S.E.; Schols, H.A.; Nauta, A.; Schoterman, M.H.C.; Keijser, B.J.F.; Montijn, R.C.; Gruppen, H.; Schuren, F.H.J. High-throughput analysis of the impact of antibiotics on the human intestinal microbiota composition. J. Microbiol. Methods 2013, 92, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Tanim Anwar, A.S.M.; Lee, J.M. Medical management of brain-dead organ donors. Acute Crit. Care 2019, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Peng, Z.; Liao, Y.; Sun, H.; Chen, W.; Chen, X.; Wei, Z.; Yang, C.; Nüssler, A.K.; Liu, J.; et al. Organ transplantation and gut microbiota: Current reviews and future challenges. Am. J. Transl. Res. 2018, 10, 3330–3344. [Google Scholar] [PubMed]
- Zieve, L.; Shekleton, M.; Lyftogt, C.; Draves, K. Ammonia, octanoate and a mercaptan depress regeneration of normal rat liver after partial hepatectomy. Hepatology 1985, 5, 28–31. [Google Scholar] [CrossRef]
- Michalopoulos, G.K. Advances in liver regeneration. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 897–907. [Google Scholar] [CrossRef]
- Liu, H.-X.; Keane, R.; Sheng, L.; Wan, Y.-J.Y. Implications of microbiota and bile acid in liver injury and regeneration. J. Hepatol. 2015, 63, 1502–1510. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.X.; Hu, Y.; Wan, Y.J.Y. Microbiota and bile acid profiles in retinoic acid-primed mice that exhibit accelerated liver regeneration. Oncotarget 2016, 7, 1096–1106. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Compare, D.; Liguori, E.; Di Mauro, V.; Rocco, A.; Barone, M.; Napoli, A.; Lapi, D.; Iovene, M.R.; Colantuoni, A. Protective effects of Lactobacillus paracasei F19 in a rat model of oxidative and metabolic hepatic injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 669–676. [Google Scholar] [CrossRef] [Green Version]
- McCormack, L.; Petrowsky, H.; Jochum, W.; Furrer, K.; Clavien, P.A. Hepatic steatosis is a risk factor for postoperative complications after major hepatectomy: A matched case-control study. Ann. Surg. 2007, 245, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.-c.; Li, L.-j.; Xu, K.-j.; Shen, T.; Chen, Y.-b.; Sheng, J.-f.; Yu, Y.-s.; Chen, Y.-g. Intestinal microflora in rats with ischemia/reperfusion liver injury. J. Zhejiang Univ. Sci. B. 2005, 6, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornell, R.P.; Liljequist, B.L.; Bartizal, K.F. Depressed liver regeneration after partial hepatectomy of germ-free, athymic and lipopolysaccharide-resistant mice. Hepatology 1990, 11, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Hu, Y.; French, S.W.; Gonzalez, F.J.; Wan, Y.J.Y. Forced expression of fibroblast growth factor 21 reverses the sustained impairment of liver regeneration in PPARαPAC mice due to dysregulated bile acid synthesis. Oncotarget 2015, 6, 9686–9700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gart, E.; Lima, E.S.; Schuren, F.; de Ruiter, C.G.F.; Attema, J.; Verschuren, L.; Keijer, J.; Salic, K.; Morrison, M.C.; Kleemann, R. Diet-independent correlations between bacteria and dysfunction of gut, adipose tissue, and liver: A comprehensive microbiota analysis in feces and mucosa of the ileum and colon in obese mice with NAFLD. Int. J. Mol. Sci. 2019, 20, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-chain fatty acids and their association with signalling pathways in inflammation, glucose and lipid metabolism. Int. J. Mol. Sci. 2020, 21, 1–16. [Google Scholar] [CrossRef]
- Hoffmann, K.; Büchler, M.W.; Schemmer, P. Supplementation of amino acids to prevent reperfusion injury after liver surgery and transplantation-Where do we stand today? Clin. Nutr. 2011, 30, 143–147. [Google Scholar] [CrossRef]
- Ardito, F.; Lai, Q.; Rinninella, E.; Mimmo, A.; Vellone, M.; Panettieri, E.; Adducci, E.; Cintoni, M.; Mele, M.C.; Gasbarrini, A.; et al. The impact of personalized nutritional support on postoperative outcome within the enhanced recovery after surgery (ERAS) program for liver resections: Results from the NutriCatt protocol. Updates Surg. 2020, 72, 681–691. [Google Scholar] [CrossRef]
- Okabayashi, T.; Nishimori, I.; Sugimoto, T.; Maeda, H.; Dabanaka, K.; Onishi, S.; Kobayashi, M.; Hanazaki, K. Effects of branched-chain amino acids-enriched nutrient support for patients undergoing liver resection for hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2008, 23, 1869–1873. [Google Scholar] [CrossRef]
- Lin, R.; Liu, W.; Piao, M.; Zhu, H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids 2017, 49, 2083–2090. [Google Scholar] [CrossRef]
- Gutiérrez-Repiso, C.; Moreno-indias, I.; de Hollanda, A.; Martín-núñez, G.M.; Vidal, J.; Tinahones, F.J. Gut microbiota specific signatures are related to the successful rate of bariatric surgery. Am. J. Transl. Res. 2019, 11, 942–952. [Google Scholar] [PubMed]
- Aron-Wisnewsky, J.; Clement, K. The Effects of Gastrointestinal Surgery on Gut Microbiota: Potential Contribution to Improved Insulin Sensitivity. Curr. Atheroscler. Rep. 2014, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mardinoglu, A.; Heiker, J.T.; Gärtner, D.; Björnson, E.; Schön, M.R.; Flehmig, G.; Klöting, N.; Krohn, K.; Fasshauer, M.; Stumvoll, M.; et al. Extensive weight loss reveals distinct gene expression changes in human subcutaneous and visceral adipose tissue. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Higashiguchi, T. Effects of glutamine administration on liver regeneration following hepatectomy. Nutrition 1999, 15, 23–28. [Google Scholar] [CrossRef]
- Magalhães, C.R.; Malafaia, O.; Torres, O.J.M.; Moreira, L.B.; Tefil, S.C.d.S.G.; Pinherio, M.d.R.; Harada, B.A. Liver regeneration with l-glutamine supplemented diet: Experimental study in rats. Rev. Col. Bras. Cir. 2014, 41, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.P.; Dong, T.S.; Agopian, V.; Lagishetty, V.; Sundaram, V.; Noureddin, M.; Ayoub, W.S.; Durazo, F.; Benhammou, J.; Enayati, P.; et al. Microbiome and bile acid profiles in duodenal aspirates from patients with liver cirrhosis: The Microbiome, Microbial Markers and Liver Disease Study. Hepatol. Res. 2018, 48, 1108–1117. [Google Scholar] [CrossRef] [Green Version]
- Duytschaever, G.; Huys, G.; Bekaert, M.; Boulanger, L.; De Boeck, K.; Vandamme, P. Dysbiosis of bifidobacteria and Clostridium cluster XIVa in the cystic fibrosis fecal microbiota. J. Cyst. Fibros. 2013, 12, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Cuenca, S.; Sanchez, E.; Santiago, A.; El Khader, I.; Panda, S.; Vidal, S.; Nieto, J.C.; Juárez, C.; Sancho, F.; Guarner, F.; et al. Microbiome composition by pyrosequencing in mesenteric lymph nodes of rats with CCl4-induced cirrhosis. J. Innate Immun. 2014, 6, 263–271. [Google Scholar] [CrossRef]
- Soto, G.; Rodríguez, M.J.; Fuentealba, R.; Treuer, A.V.; Castillo, I.; González, D.R.; Zúñiga-Hernández, J. Maresin 1, a proresolving lipid mediator, ameliorates liver ischemia-reperfusion injury and stimulates hepatocyte proliferation in sprague-dawley rats. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Sun, R.; Chen, Y.; Zheng, X.; Bai, L.; Lian, Z.; Wei, H.; Tian, Z. Oral Ampicillin Inhibits Liver Regeneration by Breaking Hepatic Innate Immune Tolerance Normally Maintained by Gut Commensal Bacteria. Hepatology 2015, 62, 253–264. [Google Scholar] [CrossRef]
- Ling, J.; Chaba, T.; Zhu, L.F.; Jacobs, R.L.; Vance, D.E. Hepatic ratio of phosphatidylcholine to phosphatidylethanolamine predicts survival after partial hepatectomy in mice. Hepatology 2012, 55, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, A.A.; Zheng, D.; Shibolet, O.; Elinav, E. The role of the microbiome in NAFLD and NASH. EMBO Mol. Med. 2019, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kakiyama, G.; Pandak, W.M.; Gillevet, P.M.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; Takei, H.; Muto, A.; Nittono, H.; Ridlon, J.M.; et al. Modulation of the fecal bile acid profile by gut microbiota in cirrhosis. J. Hepatol. 2013, 58, 949–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Vitetta, L. Gut microbiota metabolites in nafld pathogenesis and therapeutic implications. Int. J. Mol. Sci. 2020, 21, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Raka, F.; Adeli, K. The Role of the Gut Microbiota in Lipid and Lipoprotein Metabolism. J. Clin. Med. 2019, 8, 2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sittipo, P.; Shim, J.W.; Lee, Y.K. Microbial metabolites determine host health and the status of some diseases. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Doycheva, I.; Leise, M.D.; Watt, K.D. The intestinal microbiome and the liver transplant recipient: What we know and what we need to know. Transplantation 2016, 100, 61–68. [Google Scholar] [CrossRef]
- Xing, H.C.; Li, L.J.; Xu, K.J.; Shen, T.; Chen, Y.B.; Sheng, J.F.; Chen, Y.; Fu, S.Z.; Chen, C.L.; Wang, J.G.; et al. Protective role of supplement with foreign Bifidobacterium and Lactobacillus in experimental hepatic ischemia-reperfusion injury. J. Gastroenterol. Hepatol. 2006, 21, 647–656. [Google Scholar] [CrossRef]
- Ren, Z.; Cui, G.; Lu, H.; Chen, X.; Jiang, J.; Liu, H.; He, Y.; Ding, S.; Hu, Z.; Wang, W.; et al. Liver Ischemic Preconditioning (IPC) Improves Intestinal Microbiota Following Liver Transplantation in Rats through 16s rDNA-Based Analysis of Microbial Structure Shift. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Koh, W.U.; Kim, J.; Lee, J.; Song, G.W.; Hwang, G.S.; Tak, E.; Song, J.G. Remote ischemic preconditioning and diazoxide protect from hepatic ischemic reperfusion injury by inhibiting HMGB1-induced TLR4/MyD88/NF-κB signaling. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Sawas, T.; Al Halabi, S.; Hernaez, R.; Carey, W.D.; Cho, W.K. Patients Receiving Prebiotics and Probiotics Before Liver Transplantation Develop Fewer Infections Than Controls: A Systematic Review and Meta-Analysis. Clin. Gastroenterol. Hepatol. 2015, 13, 1567–1574.e3. [Google Scholar] [CrossRef] [PubMed]
- Wiest, R.; Albillos, A.; Trauner, M.; Bajaj, J.S.; Jalan, R. Targeting the gut-liver axis in liver disease. J. Hepatol. 2017, 67, 1084–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, T.C.D.; Pyrsopoulos, N.; Rustgi, V.K. Microbiota and the liver. Liver Transplant. 2018, 24, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, J.; Wu, J.; Chalson, H.; Merigan, L.; Mitchell, A. Probiotic use in preventing postoperative infection in liver transplant patients. Hepatobiliary Surg. Nutr. 2013, 2, 142–147. [Google Scholar] [CrossRef]
- Rayes, N.; Seehofer, D.; Hansen, S.; Boucsein, K.; Müller, A.R.; Serke, S.; Bengmark, S.; Neuhaus, P. Early enteral supply of Lactobacillus and fiber versus selective bowel decontamination: A controlled trial in liver transplant recipients. Transplantation 2002, 74, 123–128. [Google Scholar] [CrossRef]
- Rayes, N.; Seehofer, D.; Theruvath, T.; Schiller, R.A.; Langrehr, J.M.; Jonas, S.; Bengmark, S.; Neuhaus, P. Supply of pre- and probiotics reduces bacterial infection rates after liver transplantation—A randomized, double-blind trial. Am. J. Transplant. 2005, 5, 125–130. [Google Scholar] [CrossRef]
- Arai, M.; Mochida, S.; Ohno, A.; Arai, S.; Fujiwara, K. Selective bowel decontamination of recipients for prevention against liver injury following orthotopic liver transplantation: Evaluation with rat models. Hepatology 1998, 27, 123–127. [Google Scholar] [CrossRef]
- Sun, H.Y.; Wagener, M.; Cacciarelli, T.V.; Singh, N. Impact of rifaximin use for hepatic encephalopathy on the risk of early post-transplant infections in liver transplant recipients. Clin. Transplant. 2012, 26, 849–852. [Google Scholar] [CrossRef]
- Nakamura, K.; Kageyama, S.; Ito, T.; Hirao, H.; Kadono, K.; Aziz, A.; Dery, K.J.; Everly, M.J.; Taura, K.; Uemoto, S.; et al. Antibiotic pretreatment alleviates liver transplant damage in mice and humans. J. Clin. Invest. 2019, 129, 3420–3434. [Google Scholar] [CrossRef] [Green Version]
- Hellinger, W.C.; Yao, J.D.; Alvarez, S.; Blair, J.E.; Cawley, J.J.; Paya, C.V.; O’Brien, P.C.; Spivey, J.R.; Dickson, R.C.; Harnois, D.M.; et al. A randomized, prospective, double-blinded evaluation of selective bowel decontamination in liver transplantation. Transplantation 2002, 73, 1904–1909. [Google Scholar] [CrossRef]
- Annavajhala, M.K.; Gomez-Simmonds, A.; Macesic, N.; Sullivan, S.B.; Kress, A.; Khan, S.D.; Giddins, M.J.; Stump, S.; Kim, G.I.; Narain, R.; et al. Colonizing multidrug-resistant bacteria and the longitudinal evolution of the intestinal microbiome after liver transplantation. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Zhou, J.; Dai, X.; Zhou, H.; Pan, X.; Wang, X.; Zhang, F.; Rao, J.; Lu, L. Short-term starvation attenuates liver ischemia-reperfusion injury (IRI) by sirt1-autophagy signaling in mice. Am. J. Transl. Res. 2016, 8, 3364–3375. [Google Scholar] [PubMed]
- Rickenbacher, A.; Jang, J.H.; Limani, P.; Ungethüm, U.; Lehmann, K.; Oberkofler, C.E.; Weber, A.; Graf, R.; Humar, B.; Clavien, P.A. Fasting protects liver from ischemic injury through Sirt1-mediated downregulation of circulating HMGB1 in mice. J. Hepatol. 2014, 61, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Dai, X.; Shen, G.; Lu, X.; Wang, X.; Lu, L.; Qian, X.; Rao, J. Preoperative short-term fasting protects liver injury in patients undergoing hepatectomy. Ann. Transl. Med. 2018, 6, 449. [Google Scholar] [CrossRef]
- Page, A.J.; Ejaz, A.; Spolverato, G.; Zavadsky, T.; Grant, M.C.; Galante, D.J.; Wick, E.C.; Weiss, M.; Makary, M.A.; Wu, C.L.; et al. Enhanced Recovery After Surgery Protocols for Open Hepatectomy—Physiology, Immunomodulation, and Implementation. J. Gastrointest. Surg. 2015, 19, 387–399. [Google Scholar] [CrossRef]
- Zaborin, A.; Penalver Bernabe, B.; Keskey, R.; Sangwan, N.; Hyoju, S.; Gottel, N.; Gilbert, J.A.; Zaborina, O.; Alverdy, J.C. Spatial Compartmentalization of the Microbiome between the Lumen and Crypts Is Lost in the Murine Cecum following the Process of Surgery, Including Overnight Fasting and Exposure to Antibiotics. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Caraceni, P.; Nardo, B.; Domenicali, M.; Turi, P.; Vici, M.; Simoncini, M.; De Maria, N.; Trevisani, F.; Van Thiel, D.H.; Derenzini, M.; et al. Ischemia-reperfusion injury in rat fatty liver: Role of nutritional status. Hepatology 1999, 29, 1139–1146. [Google Scholar] [CrossRef]
- Ren, J.; Hu, D.; Mao, Y.; Yang, H.; Liao, W.; Xu, W.; Ge, P.; Zhang, H.; Sang, X.; Lu, X.; et al. Alteration in gut microbiota caused by time-restricted feeding alleviate hepatic ischaemia reperfusion injury in mice. J. Cell. Mol. Med. 2019, 23, 1714–1722. [Google Scholar] [CrossRef] [Green Version]
- Melkani, G.C.; Panda, S. Time-restricted feeding for prevention and treatment of cardiometabolic disorders. J. Physiol. 2017, 595, 3691–3700. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.G.; Liu, H.; Jiang, J.W.; Jiang, L.; Chen, H.; Xie, H.Y.; Zhou, L.; Zheng, S. Sen Protective effect of probiotics on intestinal barrier function in malnourished rats after liver transplantation. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 489–496. [Google Scholar] [CrossRef]
- Kamo, N.; Kaido, T.; Hamaguchi, Y.; Uozumi, R.; Okumura, S.; Kobayashi, A.; Shirai, H.; Yagi, S.; Okajima, H.; Uemoto, S. Impact of Enteral Nutrition with an Immunomodulating Diet Enriched with Hydrolyzed Whey Peptide on Infection After Liver Transplantation. World J. Surg. 2018, 42, 3715–3725. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.L.; Yu, T.T.; Zhong, J.X.; Lei, T. Effect of glutamine on intestinal barrier function following liver transplantation in rats. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2058–2064. [Google Scholar] [PubMed]
- Rayes, N.; Pilarski, T.; Stockmann, M.; Bengmark, S.; Neuhaus, P.; Seehofer, D. Effect of pre- and probiotics on liver regeneration after resection: A randomised, double-blind pilot study. Benef. Microbes 2012, 3, 237–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rifatbegovic, Z.; Mesic, D.; Ljuca, F.; Zildzic, M.; Avdagic, M.; Grbic, K.; Agic, M.; Hadziefendic, B. Effect of probiotics on liver function after surgery resection for malignancy in the liver cirrhotic. Med. Arh. 2010, 64, 208–211. [Google Scholar] [PubMed]
- Gan, Y.; Su, S.; Li, B.; Fang, C. Efficacy of probiotics and prebiotics in prevention of infectious complications following hepatic resections: Systematic review and meta-analysis. J. Gastrointest. Liver Dis. 2019, 28, 205–211. [Google Scholar] [CrossRef]
- Mendes-Braz, M.; Elias-Miró, M.; Kleuser, B.; Fayyaz, S.; Jiménez-Castro, M.B.; Massip-Salcedo, M.; Gracia-Sancho, J.; Ramalho, F.S.; Rodes, J.; Peralta, C. The effects of glucose and lipids in steatotic and non-steatotic livers in conditions of partial hepatectomy under ischaemia-reperfusion. Liver Int. 2014, 34, 271–289. [Google Scholar] [CrossRef]
- Yang, Y.; Shao, C.; Zhang, W.; Wang, G.; Lu, D.C.; Han, W.; Wu, Z.S.; Chen, C.B. Omega-3 polyunsaturated fatty acids prevent progression of liver fibrosis and promote liver regeneration after partial hepatectomy in cirrhotic rats. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10151–10160. [Google Scholar] [CrossRef]
- Beppu, T.; Nitta, H.; Hayashi, H.; Imai, K.; Okabe, H.; Nakagawa, S.; Hashimoto, D.; Chikamoto, A.; Ishiko, T.; Yoshida, M.; et al. Effect of branched-chain amino acid supplementation on functional liver regeneration in patients undergoing portal vein embolization and sequential hepatectomy: A randomized controlled trial. J. Gastroenterol. 2015, 50, 1197–1205. [Google Scholar] [CrossRef]
- Nanno, Y.; Toyama, H.; Terai, S.; Mizumoto, T.; Tanaka, M.; Kido, M.; Ajiki, T.; Fukumoto, T. Preoperative Oral Branched-Chain Amino Acid Supplementation Suppresses Intraoperative and Postoperative Blood Lactate Levels in Patients Undergoing Major Hepatectomy. J. Parenter Enter. Nutr. 2019, 43, 220–225. [Google Scholar] [CrossRef]
- Ucar, M.; Aydogan, M.S.; Vardı, N.; Parlakpınar, H. Protective Effect of Dexpanthenol on Ischemia-Reperfusion-Induced Liver Injury. Transplant. Proc. 2018, 50, 3135–3143. [Google Scholar] [CrossRef]
- Shama, S.; Liu, W. Omega-3 Fatty Acids and Gut Microbiota: A Reciprocal Interaction in Nonalcoholic Fatty Liver Disease. Dig. Dis. Sci. 2020, 65, 906–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhang, X.; Zhu, L.; Yang, X.; He, F.; Wang, T.; Bao, T.; Lu, H.; Wang, H.; Yang, S. Inulin alleviates inflammation of alcoholic liver disease via SCFAs-inducing suppression of M1 and facilitation of M2 macrophages in mice. Int. Immunopharmacol. 2020, 78, 106062. [Google Scholar] [CrossRef] [PubMed]
- Zambom de Souza, A.Z.; Zambom, A.Z.; Abboud, K.Y.; Reis, S.K.; Tannihão, F.; Guadagnini, D.; Saad, M.J.A.; Prada, P.O. Oral supplementation with l-glutamine alters gut microbiota of obese and overweight adults: A pilot study. Nutrition 2015, 31, 884–889. [Google Scholar] [CrossRef] [PubMed]


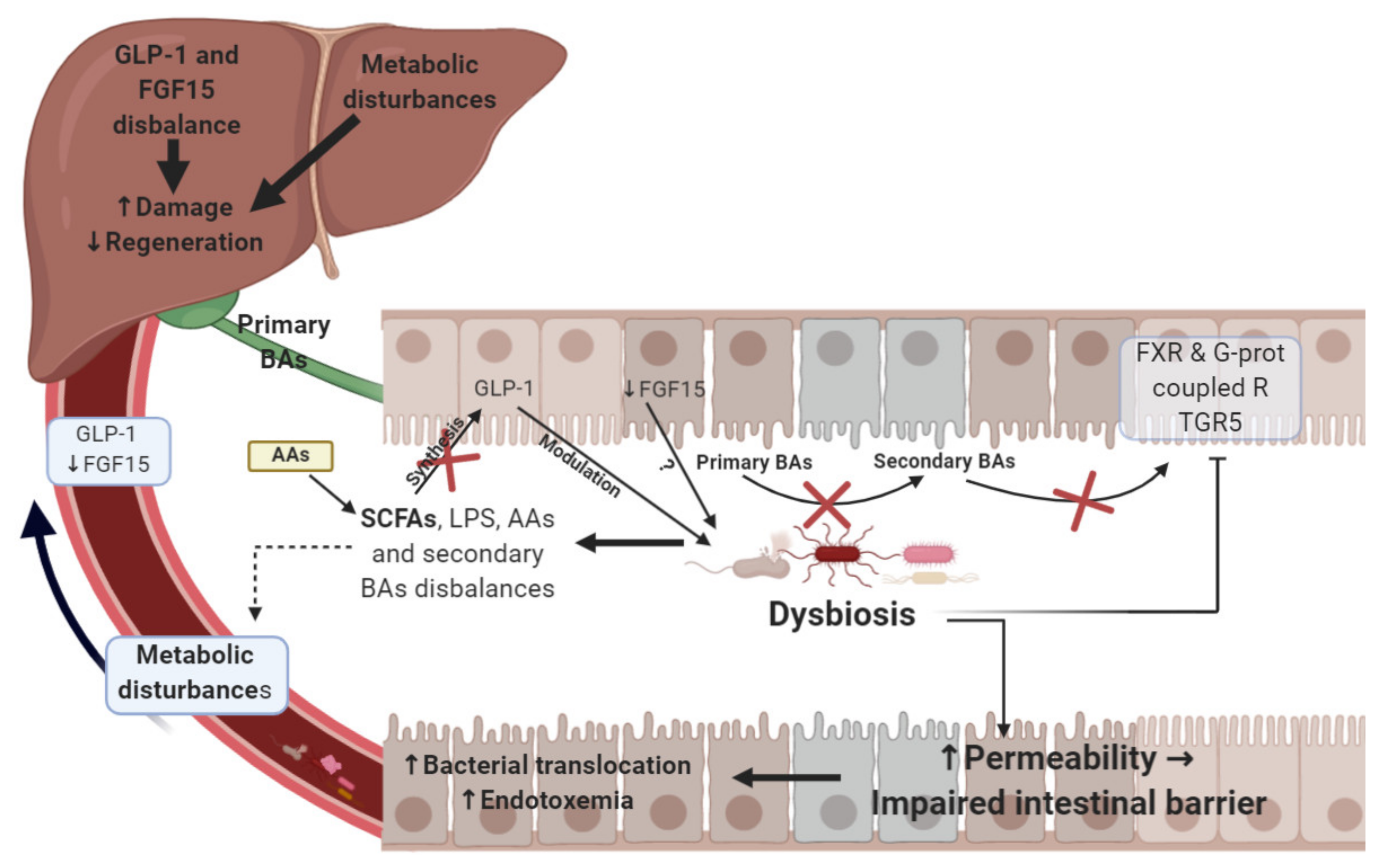{kind=link}
| Study | Surgery (DCD/BD) | Recipients Characteristics | Alteration Post-LT |
|---|---|---|---|
| Xie et al. (2011) [83] | OLT without DCD/BD | Normal SD rats (donors) and normal SD or SD with cirrhosis (recipients). Groups: -LN (n = 12; 6 transplanted rats) -LTC (n = 14; 7 transplanted rats) | ↑ endotoxin, BT and bacteria number in LTC rats. = Lactobacilli, Bacteroides and Enterobacteria between groups. ↑ MUC2/3 and TLR4 in LTC rats. Slight rejection and periportal inflammatory cell infiltration in both LN and LTC rat |
| Xie et al. (2012) [84] | OLT without DCD/BD | L and BN rats (donors) and BN rats (recipients). Groups: -BN rats transplanted from L rats (n =12; 6 transplanted BN rats) -BN rats transplanted from BN rats (n = 12; 6 transplanted BN rats) | ↑ endotoxin and BT to the liver with AR ↑ Bacteroides associated to AR = Clostridium leptum, Enterobacteriaceae and Lactobacillus after 1–2 weeks post-LT Better hepatic architecture in isograft group ↑ Liver function index in allograft group. |
| Ren et al. (2014) [85] | OLT without DCD/BD | L and DA rats (donors) and L rats (recipients). Groups: -NR: OLT from L rats to L rats (n = 18; 9 transplanted rats) -AR: OLT from DA (specific-pathogen free rats) rats to L rats. (n = 18; 9 transplanted rats) | ↓ Faecalibacterium prausnitzii and Lactobacillus during AR ↑ Clostridium bolteae during AR Altered intestinal integrity ↑ endotoxin ↑ hepatic injury in AR rats |
| Xie et al. (2014) [86] | OLT without DCD/BD | L (donors) and BN rats (recipients). Groups: Allograft group (n = 16; 8 transplanted rats) Antibiotic group (n = 16; 8 transplanted rats) Probiotic group (n = 16; 8 transplanted rats) | ↓ endotoxemia in the antibiotic ↑ Lactobacillus, C. leptum and Bifidobacterium in the probiotic group = Bacteroides in all groups at 7 and 14 days after LT ↓ Necrosis, inflammation and AST in the antibiotic and probiotic groups |
| Study | Surgery (DCD/BD) | Population | IS | Antibiotics | Alterations Post-LT |
|---|---|---|---|---|---|
| Wu et al. (2012) [72] | No specified | -28 healthy volunteers -51 cirrhotic patients -111 liver-transplanted patients with cancer or cirrhosis. | -GC -MPA -CNIs | No antibiotics use | ↓ Eubacteria, Bifidobacterium spp., Faecalibacterium prausnitzii and Lactobacillus spp. in LTR. ↑ Enterobacteriaceae and Enterococcus spp. in LTR Enterococcus spp. showed a tendency to restore to normal levels ↑ Plasma endotoxin, IL-6 and fecal sIgA in cirrhotic patients but not in LTR |
| Lu et al. (2013) [95] | No specified | 12 OLT recipients with liver failure from hepatitis B virus (HBV) liver cirrhosis | Different for each patient | No antibiotics use | ↓ microbial diversity in some patients Presence of infection in some patients |
| Bajaj et al. (2017) [96] | No specified | 45 LT patients with cirrhosis | -Steroids -MPA -Tacrolimus | TMP-SMX | Dysbiosis after LT |
| Kato et al. (2017) [81] | Deceased and living donor | 38 LT patients (exclusion of patients with fulminant hepatitis or with a previous transplant) | -Tacrolimus -MPA -Steroids | 13 received broad spectrum of antibiotics | ↓ microbiota diversity associated with high ESLD scores and ACR ↑ Bacteroides, Enterobacteriaceae, Streptococcaceae and Bifidobacteriaceae ↓ Enterococcaceae, Lactobacillaceae, Clostridiaceae, Ruminococcaceae and Peptostreptococcaceae |
| Sun et al. (2017) [97] | No specified | 9 LT with ESLD (4 patients with cirrhosis and 5 with HCC) 15 healthy controls | -Tacrolimus -MP | Cephalosporin | ↓ Actinobacillus, Escherichia, and Shigella after LT ↑ Micromonosporaceae, Desulfobacterales, the Sarcina genus of Eubacteriaceae, and Akkermansia |
| Bajaj et al. (2018) [15] | Deceased donors | -40 patients with cirrhosis -Patients are their own controls | -Steroids -MPA -Tacrolimus | TMP-SMX | ↑ bacterial action after LT |
| Lu et al. (2019) [42] | No specified | -Healthy controls (n = 61) -LTR with HCC (n = 90): 42 with LTA and 48 with LTN | -FK506 tacrolimus | No antibiotic use in the 12 weeks prior to enrollment | ↓ fecal microbiome diversity in recipients in the LTA group ↑ opportunistic pathogens such as Klebsiella and Escherichia/Shigella in all LTR ↓ beneficial butyrate-producing bacteria in LTR |
| Study | Surgery (IR/PH) | Population | Treatment | Effect/Alteration Post-Surgery |
|---|---|---|---|---|
| Xing et al. (2005) [112] | -I/R (20 min ischemia and 22 h of reperfusion) | -SD rats with normal liver. Groups: -Sham -I/R n = 6–10/group | No specified | I/R→↑ALT, AST, MDA, LPS and ↓SOD. I/R→↓Bifidobacteria, Lactobacilli and ↑Enterobacterium, Enterococcus. I/R→↑BT to kidney, ↓intestinal microvilli. |
| Nardone et al. (2010) [110] | -I/R (30 min ischemia and 60 min reperfusion) | -Male W rats with healthy or steatotic liver. Groups: -Rats fed with Standard or Steatogenic diet -Rats fed with Standard or steatogenic diet + 8 week probiotic supplementation n = 7–10/group | LP-F19 supplementation | I/R→↓Bacteroides, Bifidobacterium and Lactobacillus and ↑Enterococcus, and Enterobacteriaceae I/R→necrosis, leukocyte infiltration, ↑MDA, TNFα, IL-1β, IL-6, ALT, AST and LPS, (especially in steatotic livers). LP-F19→↓I/R injury in steatogen diet, ↓↓I/R in standard diet. |
| Wu et al. (2015) [130] | -PH (67%) | -Male C57Bl/6 mice with healthy liver. Groups: -Mice undergoing PH -Mice undergoing PH+Atb treatment: Amp Neo, Met, Vanco (in combination or separately). n = 3–6/group | (Atb-treatment): Amp, Neo, Met, Vanco | Atb in combination: no affect hepatic damage (AST and ALT) but ↓hepatic proliferation (↓PCNA, BrdU-positive hepatocytes). Vanco, Neo, Met alone→no effect on LR. Amp→↓Amp-sensitive commensal bacteria (↓many gram-negative and gram-positive bacteria →↑Kupffer cells and ↑IL-12→↑activation NKT cells→↓LR |
| Liu et al. (2016) [13] | -PH (2/3) | -Male C57BL/6 mice with healthy liver Groups: -Control -Mice undergoing PH Time: 0 h-9 days. n = 3, 4/group. | No specified | -PH: ↑Bacteroidetes (S24, Rikenellaceae), ↓Firmicutes (Clostridiales, Lachnospiraceae, and Ruminococcaceae). -Ruminococus, Bifidobacterium, Lactobacillus and Clostridium modulate bile acid conversion ↓Firmicutes/Bacteroidetes ratio is associated to lean microbiota; ↑Ratio: obese mice. High fat diet: ↑Rikenellaceae and ↓Ruminicoccaceae. Exercice: ↑S24-7→↑butyrate ↑Ki67 following PH and liver size was restored at 7–9 days. |
| Bao et al. (2020) [14] | -PH (2/3) | Adult male SD rats (n = 126) with healthy liver. Groups: -Control -Rats undergoing PH -Rats undergoing PH+Atb treatment -Rats undergoing PH+Atb +FMT treatment Time: 0–336 h | -Atb-treatment: Amp, Vanco, Metro, Neo (in combination) -FMT treatment | PH: -Inverse relation between Bacteroidetes (Bacteroidaceae, Prevotellaceae, Rikenellaceae, Porphyromonadaceae) and Firmicutes (Ruminococcaceae, Lachnospiraceae). Bacteroidetes ↓12–24 h, ↑30–48 h and ↓3–14 days Proteobacteria ↑48 h PH: ↑Ki67, BrdU, TNFα, IL-6 and HGF. -Atb treatment impaired LR (↓Ki67 and BrdU) and FMT treatment restored LR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micó-Carnero, M.; Rojano-Alfonso, C.; Álvarez-Mercado, A.I.; Gracia-Sancho, J.; Casillas-Ramírez, A.; Peralta, C. Effects of Gut Metabolites and Microbiota in Healthy and Marginal Livers Submitted to Surgery. Int. J. Mol. Sci. 2021, 22, 44. https://doi.org/10.3390/ijms22010044
Micó-Carnero M, Rojano-Alfonso C, Álvarez-Mercado AI, Gracia-Sancho J, Casillas-Ramírez A, Peralta C. Effects of Gut Metabolites and Microbiota in Healthy and Marginal Livers Submitted to Surgery. International Journal of Molecular Sciences. 2021; 22(1):44. https://doi.org/10.3390/ijms22010044
Chicago/Turabian StyleMicó-Carnero, Marc, Carlos Rojano-Alfonso, Ana Isabel Álvarez-Mercado, Jordi Gracia-Sancho, Araní Casillas-Ramírez, and Carmen Peralta. 2021. "Effects of Gut Metabolites and Microbiota in Healthy and Marginal Livers Submitted to Surgery" International Journal of Molecular Sciences 22, no. 1: 44. https://doi.org/10.3390/ijms22010044
APA StyleMicó-Carnero, M., Rojano-Alfonso, C., Álvarez-Mercado, A. I., Gracia-Sancho, J., Casillas-Ramírez, A., & Peralta, C. (2021). Effects of Gut Metabolites and Microbiota in Healthy and Marginal Livers Submitted to Surgery. International Journal of Molecular Sciences, 22(1), 44. https://doi.org/10.3390/ijms22010044