Increasing Oxygen Partial Pressures Induce a Distinct Transcriptional Response in Human PBMC: A Pilot Study on the “Normobaric Oxygen Paradox”

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
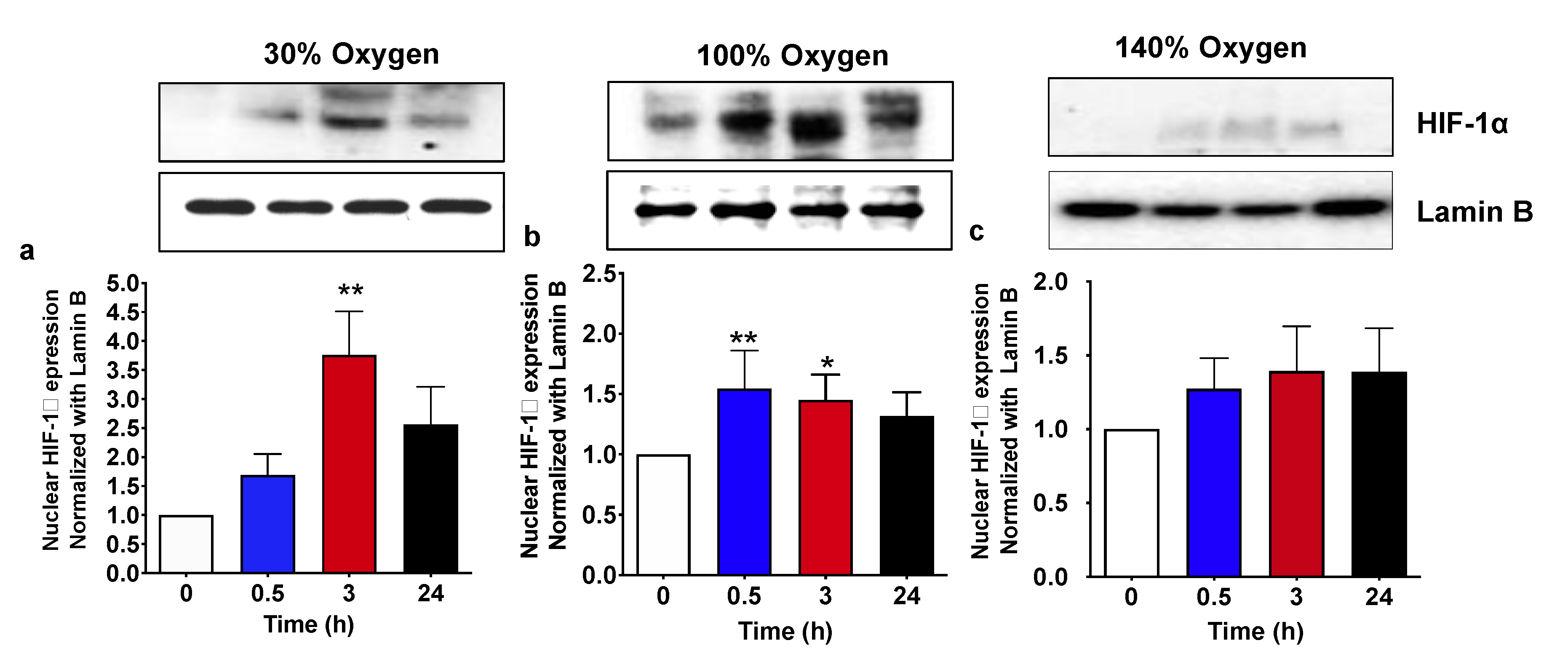
2.1. Pulsed Mild Hyperoxia, but Not High Hyperoxia, Induces HIF-1α Nuclear Transfer in PBMC
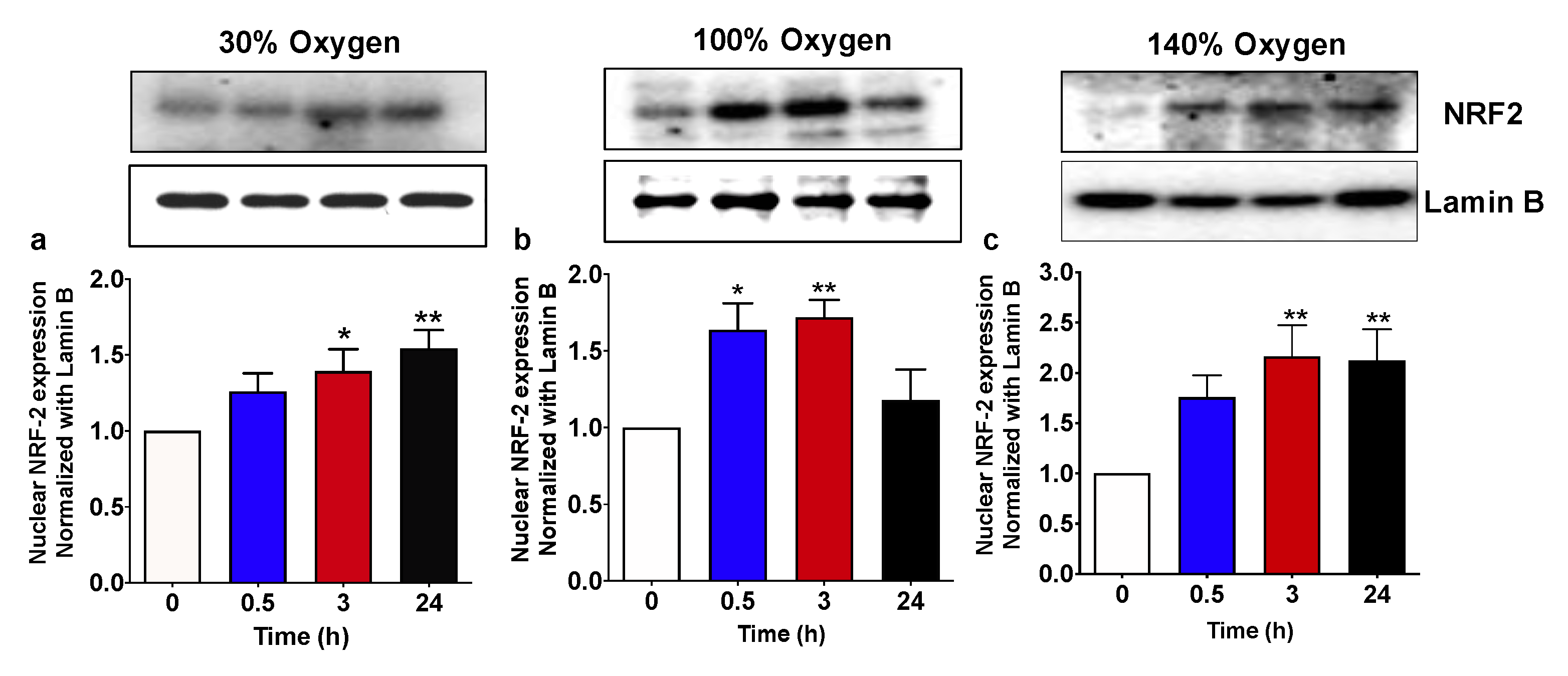
2.2. Pulsed Hyperoxic Treatment, Significantly Affects NRF2 Nuclear Transfer in PBMC
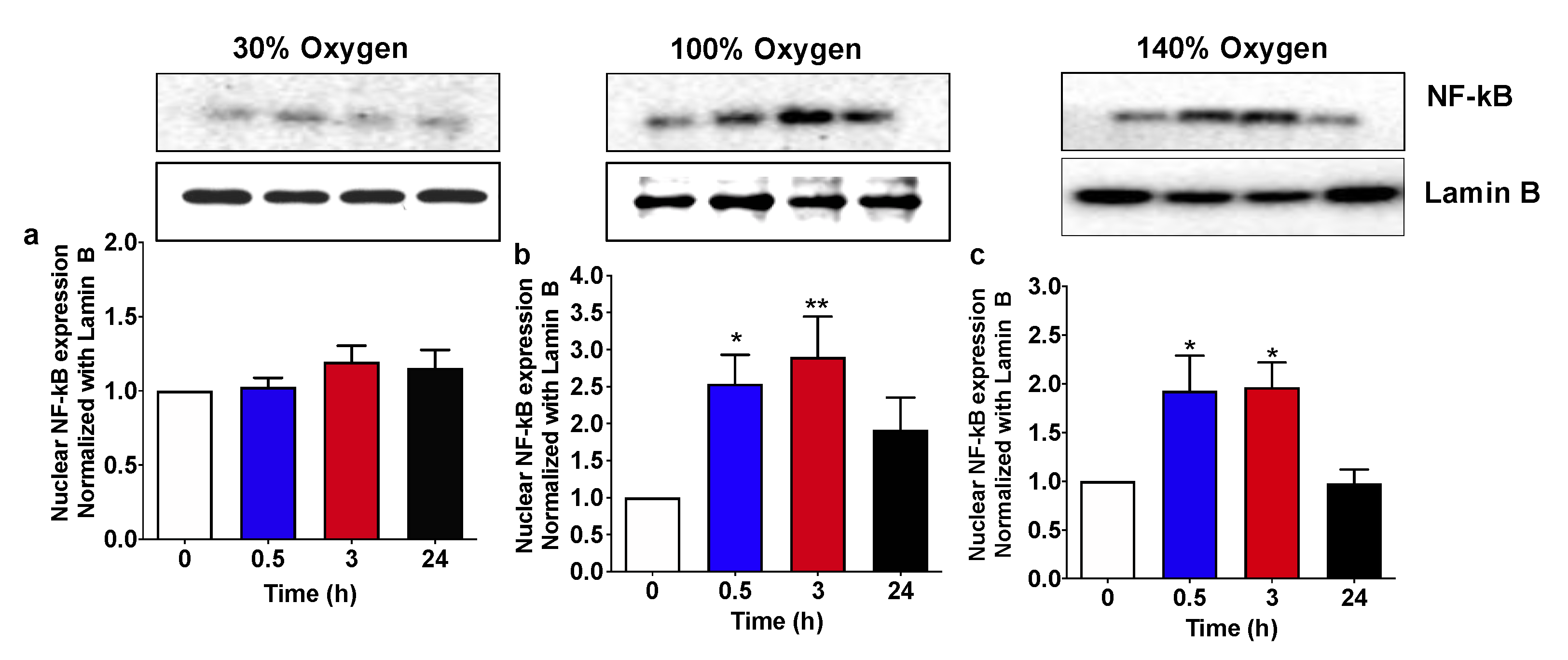
2.3. High and Very High Hyperoxia but Not Mild Hyperoxia Activate NF-KB Nuclear Traslocation in Human PBMCs
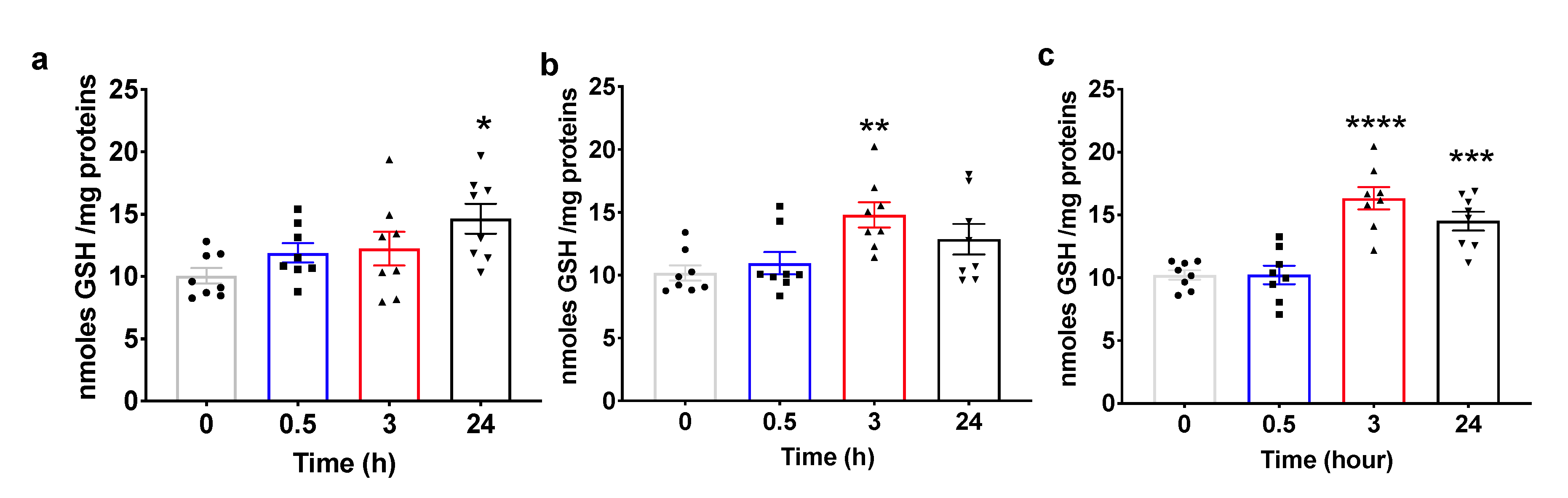
2.4. Pulsed Hyperoxic Treatment Modulates Plasma Total Glutathione Levels
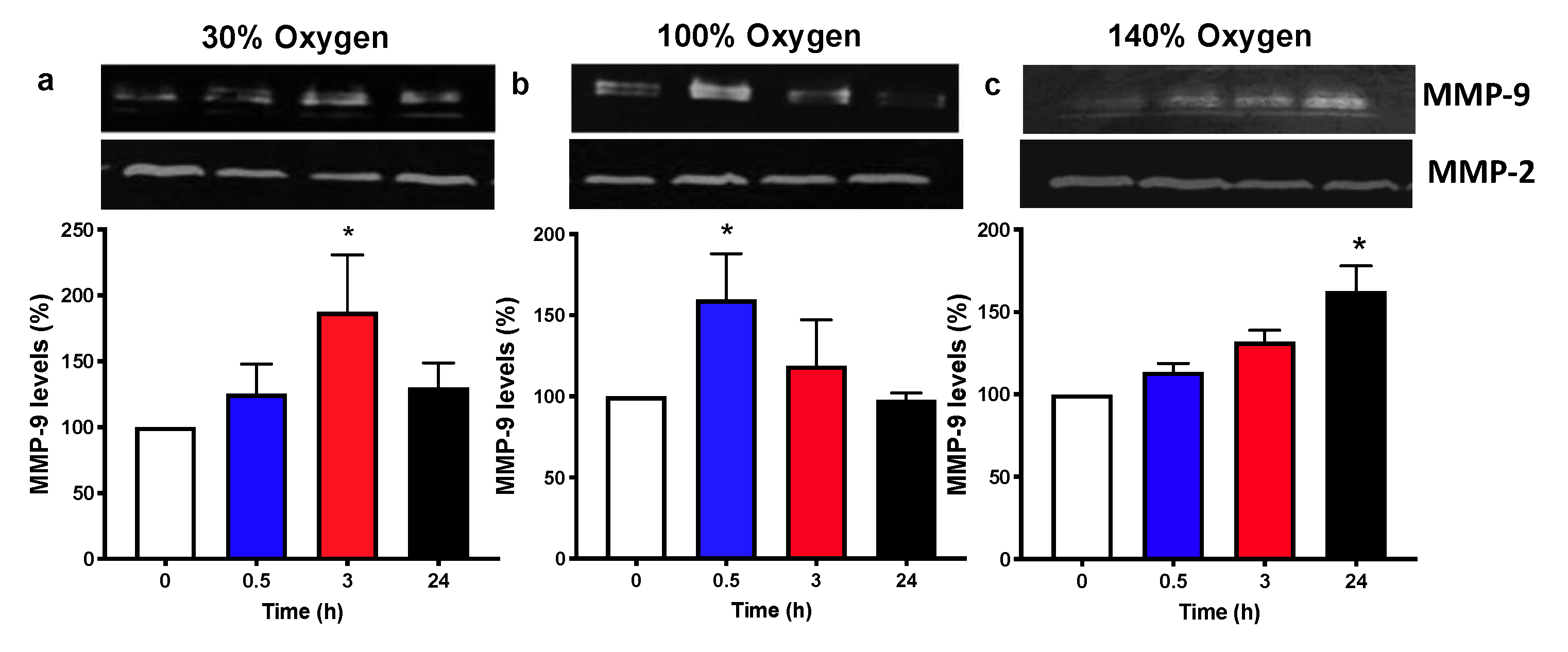
2.5. Pulsed Hyperoxic Treatment Modulates MMP-9 Release and Activity in Human Plasma
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Protocol
5.2. Nuclear Lysate Preparation and Western Blotting Analysis
5.3. Intracellular-Reduced Glutathione (GSH) Assay
5.4. Zymographic Analysis of Matrix Metalloproteinase (MMP)-2 and -9 Plasma Levels
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| EPO | Erythropoietin |
| GSH | Intracellular reduced glutathione |
| HIF-1α | Hypoxia-inducible factor 1α subunit |
| MPPs | Matrix metallopeptidases |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NOP | Normobaric Oxygen Paradox |
| NRF2 | Nuclear factor (erythroid-derived 2)-like 2 |
| PBMC | Peripheral Blood Mononuclear Cells |
| PO2 | Oxygen Partial Pressure |
References
- Mattson, M.P. Hormesis and disease resistance: Activation of cellular stress response pathways. Hum. Exp. Toxicol. 2008, 27, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu. Rev. Pathol. 2014, 9, 47–71. [Google Scholar] [CrossRef]
- Heyman, S.N.; Leibowitz, D.; Levi, I.M.-Y.; Liberman, A.; Eisenkraft, A.; Alcalai, R.; Khamaisi, M.; Rosenberger, C. Adaptive response to hypoxia and remote ischaemia pre-conditioning: A new hypoxia-inducible factors era in clinical medicine. Acta Physiol. 2016, 216, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Radak, Z.; Zhao, Z.; Koltai, E.; Ohno, H.; Atalay, M. Oxygen consumption and usage during physical exercise: The balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid. Redox Signal. 2013, 18, 1208–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perovic, A.; Unic, A.; Dumic, J. Recreational scuba diving: Negative or positive effects of oxidative and cardiovascular stress? Biochem. Med. 2014, 24, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Matzi, V.; Greilberger, J.F.; Lindenmann, J.; Neuboeck, N.; Nuhsbaumer, S.; Zelzer, S.; Tafeit, E.; Maier, A.; Smolle-Juettner, M.F. Application of hyperbaric oxygen reduce oxidative damage of plasmatic carbonyl proteins and 8-OHdG by activating glutathion peroxidase. Clin. Lab. 2015, 61, 587–593. [Google Scholar] [CrossRef]
- Davies, K.J.; Quintanilha, A.T.; Brooks, G.A.; Packer, L. Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res. Commun. 1982, 107, 1198–1205. [Google Scholar] [CrossRef]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Fratantonio, D.; Cimino, F.; Speciale, A.; Virgili, F. Need (more than) two to Tango: Multiple tools to adapt to changes in oxygen availability. Biofactors 2018, 44, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Balestra, C.; Germonpre, P.; Poortmans, J.R.; Marroni, A. Serum erythropoietin levels in healthy humans after a short period of normobaric and hyperbaric oxygen breathing: The “normobaric oxygen paradox”. J. Appl. Physiol. 2006, 100, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Cimino, F.; Balestra, C.; Germonpre, P.; de Bels, D.; Tillmans, F.; Saija, A.; Speciale, A.; Virgili, F. Pulsed high oxygen induces a hypoxic-like response in human umbilical endothelial cells and in humans. J. Appl. Physiol. 2012, 113, 1684–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadanny, A.; Efrati, S. The hyperoxic-hypoxic paradox. Biomolecules 2020, 10, 958. [Google Scholar] [CrossRef] [PubMed]
- Balestra, C.; Germonpre, P. Hypoxia, a multifaceted phenomenon: The example of the “normobaric oxygen paradox”. Eur. J. Appl. Physiol. 2012, 112, 4173–4175. [Google Scholar] [CrossRef] [PubMed]
- Burk, R. Oxygen breathing may be a cheaper and safer alternative to exogenous erythropoietin (EPO). Med. Hypotheses 2007, 69, 1200–1204. [Google Scholar] [CrossRef] [PubMed]
- Balestra, C.; Germonpre, P. Increasing EPO using the normobaric oxygen paradox: A “not so simple” task. Acta Physiol. 2011, 203, 287–288. [Google Scholar] [CrossRef]
- De Bels, D.; Corazza, F.; Germonpre, P.; Balestra, C. The normobaric oxygen paradox: A novel way to administer oxygen as an adjuvant treatment for cancer? Med. Hypotheses 2011, 76, 467–470. [Google Scholar] [CrossRef]
- Donati, A.; Damiani, E.; Zuccari, S.; Domizi, R.; Scorcella, C.; Girardis, M.; Giulietti, A.; Vignini, A.; Adrario, E.; Romano, R.; et al. Effects of short-term hyperoxia on erythropoietin levels and microcirculation in critically ill patients: A prospective observational pilot study. BMC Anesthesiol. 2017, 17, 49. [Google Scholar] [CrossRef] [Green Version]
- Lafere, P.; Schubert, T.; de Bels, D.; Germonpre, P.; Balestra, C. Can the normobaric oxygen paradox (NOP) increase reticulocyte count after traumatic hip surgery? J. Clin. Anesth. 2013, 25, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Rocco, M.; D’Itri, L.; de Bels, D.; Corazza, F.; Balestra, C. The “normobaric oxygen paradox”: A new tool for the anesthetist? Minerva Anestesiol. 2014, 80, 366–372. [Google Scholar]
- Keramidas, M.E.; Kounalakis, S.N.; Debevec, T.; Norman, B.; Gustafsson, T.; Eiken, O.; Mekjavic, I.B. Acute normobaric hyperoxia transiently attenuates plasma erythropoietin concentration in healthy males: Evidence against the ‘normobaric oxygen paradox’ theory. Acta Physiol. 2011, 202, 91–98. [Google Scholar] [CrossRef]
- Keramidas, M.E.; Norman, B.; Gustafsson, T.; Eiken, O.; Mekjavic, I.B. Long-term intermittent hyperoxic exposures do not enhance erythropoiesis. Eur. J. Clin. Investig. 2012, 42, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duansak, N.; Schmid-Schonbein, G.W. The oxygen free radicals control MMP-9 and transcription factors expression in the spontaneously hypertensive rat. Microvasc. Res. 2013, 90, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.Y.; Kleeberger, S.R. Association of Nrf2 with airway pathogenesis: Lessons learned from genetic mouse models. Arch. Toxicol. 2015, 89, 1931–1957. [Google Scholar] [CrossRef] [PubMed]
- Papaiahgari, S.; Zhang, Q.; Kleeberger, S.R.; Cho, H.Y.; Reddy, S.P. Hyperoxia stimulates an Nrf2-ARE transcriptional response via ROS-EGFR-PI3K-Akt/ERK MAP kinase signaling in pulmonary epithelial cells. Antioxid. Redox Signal. 2006, 8, 43–52. [Google Scholar] [CrossRef]
- Cho, H.Y.; Jedlicka, A.E.; Reddy, S.P.; Kensler, T.W.; Yamamoto, M.; Zhang, L.Y.; Kleeberger, S.R. Role of NRF2 in protection against hyperoxic lung injury in mice. Am. J. Respir. Cell Mol. Biol. 2002, 26, 175–182. [Google Scholar] [CrossRef]
- Cimino, F.; Speciale, A.; Anwar, S.; Canali, R.; Ricciardi, E.; Virgili, F.; Trombetta, D.; Saija, A. Anthocyanins protect human endothelial cells from mild hyperoxia damage through modulation of Nrf2 pathway. Genes Nutr. 2013, 8, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Buelna-Chontal, M.; Zazueta, C. Redox activation of Nrf2 & NF-kappaB: A double end sword? Cell. Signal. 2013, 25, 2548–2557. [Google Scholar] [CrossRef]
- Rushworth, S.A.; Zaitseva, L.; Murray, M.Y.; Shah, N.M.; Bowles, K.M.; MacEwan, D.J. The high Nrf2 expression in human acute myeloid leukemia is driven by NF-kappaB and underlies its chemo-resistance. Blood 2012, 120, 5188–5198. [Google Scholar] [CrossRef] [Green Version]
- De Bels, D.; Tillmans, F.; Corazza, F.; Bizzari, M.; Germonpre, P.; Radermacher, P.; Orman, K.G.; Balestra, C. Hyperoxia alters ultrastructure and induces apoptosis in leukemia cell lines. Biomolecules 2020, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Reddy, N.M.; Kleeberger, S.R.; Bream, J.H.; Fallon, P.G.; Kensler, T.W.; Yamamoto, M.; Reddy, S.P. Genetic disruption of the Nrf2 compromises cell-cycle progression by impairing GSH-induced redox signaling. Oncogene 2008, 27, 5821–5832. [Google Scholar] [CrossRef] [Green Version]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, F.J., Jr.; Gentene, L.J. Hyperbaric oxygen effect on MMP-9 after a vascular insult. J. Cardiovasc. Transl. Res. 2010, 3, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zheng, Y.L. Hypoxia promotes invasion of retinoblastoma cells in vitro by upregulating HIF-1alpha/MMP9 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5361–5369. [Google Scholar] [CrossRef]
- Balestra, C.; Germonpre, P.; Poortmans, J.; Marroni, A.; Schiettecatte, J.; Collard, J.F.; Snoeck, T. Erythropoietin production can be enhanced by normobaric oxygen breathing in healthy humans. Undersea Hyperb. Med. 2004, 31, 53–57. [Google Scholar]
- Chandel, N.S.; McClintock, D.S.; Feliciano, C.E.; Wood, T.M.; Melendez, J.A.; Rodriguez, A.M.; Schumacker, P.T. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1alpha during hypoxia: A mechanism of O2 sensing. J. Biol. Chem. 2000, 275, 25130–25138. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sanchez-Perez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Done, A.J.; Traustadottir, T. Nrf2 mediates redox adaptations to exercise. Redox Biol. 2016, 10, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Hachmo, Y.; Hadanny, A.; Abu Hamed, R.; Daniel-Kotovsky, M.; Catalogna, M.; Fishlev, G.; Lang, E.; Polak, N.; Doenyas, K.; Friedman, M.; et al. Hyperbaric oxygen therapy increases telomere length and decreases immunosenescence in isolated blood cells: A prospective trial. Aging 2020, 12. [Google Scholar] [CrossRef]
- Balan, E.; Decottignies, A.; Deldicque, L. Physical activity and nutrition: Two promising strategies for telomere maintenance? Nutrients 2018, 10, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diman, A.; Boros, J.; Poulain, F.; Rodriguez, J.; Purnelle, M.; Episkopou, H.; Bertrand, L.; Francaux, M.; Deldicque, L.; Decottignies, A. Nuclear respiratory factor 1 and endurance exercise promote human telomere transcription. Sci. Adv. 2016, 2, e1600031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Cuevas, I.; Parra-Llorca, A.; Sanchez-Illana, A.; Nunez-Ramiro, A.; Kuligowski, J.; Chafer-Pericas, C.; Cernada, M.; Escobar, J.; Vento, M. Oxygen and oxidative stress in the perinatal period. Redox Biol. 2017, 12, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Fratantonio, D.; Speciale, A.; Ferrari, D.; Cristani, M.; Saija, A.; Cimino, F. Palmitate-induced endothelial dysfunction is attenuated by cyanidin-3-O-glucoside through modulation of Nrf2/Bach1 and NF-kappaB pathways. Toxicol. Lett. 2015, 239, 152–160. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- St-Pierre, Y.; van Themsche, C.; Esteve, P.O. Emerging features in the regulation of MMP-9 gene expression for the development of novel molecular targets and therapeutic strategies. Curr. Drug Targets Inflamm. Allergy 2003, 2, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Ciccarella, Y.; Balestra, C.; Valsamis, J.; van der Linden, P. Increase in endogenous erythropoietin synthesis through the normobaric oxygen paradox in cardiac surgery patients. Br. J. Anaesth. 2011, 106, 752–753. [Google Scholar] [CrossRef] [Green Version]
- Revelli, L.; Vagnoni, S.; D’Amore, A.; di Stasio, E.; Lombardi, C.P.; Storti, G.; Proietti, R.; Balestra, C.; Ricerca, B.M. EPO modulation in a 14-days undersea scuba dive. Int. J. Sports Med. 2013, 34, 856–860. [Google Scholar] [CrossRef] [Green Version]
- Kiboub, F.Z.; Balestra, C.; Loennechen, O.; Eftedal, I. Hemoglobin and erythropoietin after commercial saturation diving. Front. Physiol. 2018, 9, 1176. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Guerrero, F.; Mazur, A.; Lambrechts, K.; Buzzacott, P.; Belhomme, M.; Theron, M. Reactive oxygen species, mitochondria, and endothelial cell death during in vitro simulated dives. Med. Sci. Sports Exerc. 2015, 47, 1362–1371. [Google Scholar] [CrossRef]
- Korpinar, S.; Uzun, H. The effects of hyperbaric oxygen at different pressures on oxidative stress and antioxidant status in rats. Medicina 2019, 55, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fratantonio, D.; Cimino, F.; Molonia, M.S.; Ferrari, D.; Saija, A.; Virgili, F.; Speciale, A. Cyanidin-3-O-glucoside ameliorates palmitate-induced insulin resistance by modulating IRS-1 phosphorylation and release of endothelial derived vasoactive factors. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Iannetta, M.; Zingaropoli, M.A.; Latronico, T.; Pati, I.; Pontecorvo, S.; Prezioso, C.; Pietropaolo, V.; Cortese, A.; Frontoni, M.; D’Agostino, C.; et al. Dynamic changes of MMP-9 plasma levels correlate with JCV reactivation and immune activation in natalizumab-treated multiple sclerosis patients. Sci. Rep. 2019, 9, 311. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fratantonio, D.; Virgili, F.; Zucchi, A.; Lambrechts, K.; Latronico, T.; Lafère, P.; Germonpré, P.; Balestra, C. Increasing Oxygen Partial Pressures Induce a Distinct Transcriptional Response in Human PBMC: A Pilot Study on the “Normobaric Oxygen Paradox”. Int. J. Mol. Sci. 2021, 22, 458. https://doi.org/10.3390/ijms22010458
Fratantonio D, Virgili F, Zucchi A, Lambrechts K, Latronico T, Lafère P, Germonpré P, Balestra C. Increasing Oxygen Partial Pressures Induce a Distinct Transcriptional Response in Human PBMC: A Pilot Study on the “Normobaric Oxygen Paradox”. International Journal of Molecular Sciences. 2021; 22(1):458. https://doi.org/10.3390/ijms22010458
Chicago/Turabian StyleFratantonio, Deborah, Fabio Virgili, Alessandro Zucchi, Kate Lambrechts, Tiziana Latronico, Pierre Lafère, Peter Germonpré, and Costantino Balestra. 2021. "Increasing Oxygen Partial Pressures Induce a Distinct Transcriptional Response in Human PBMC: A Pilot Study on the “Normobaric Oxygen Paradox”" International Journal of Molecular Sciences 22, no. 1: 458. https://doi.org/10.3390/ijms22010458
APA StyleFratantonio, D., Virgili, F., Zucchi, A., Lambrechts, K., Latronico, T., Lafère, P., Germonpré, P., & Balestra, C. (2021). Increasing Oxygen Partial Pressures Induce a Distinct Transcriptional Response in Human PBMC: A Pilot Study on the “Normobaric Oxygen Paradox”. International Journal of Molecular Sciences, 22(1), 458. https://doi.org/10.3390/ijms22010458