
Delivery Systems of Retinoprotective Proteins in the Retina



Abstract
:1. Introduction
2. Overview of Retinoprotective Proteins
2.1. Autocrine Retinoprotective Proteins
2.2. Paracrine Retinoprotective Proteins
2.3. Endocrine Retinoprotective Proteins
3. PEDF
3.1. Biogenesis
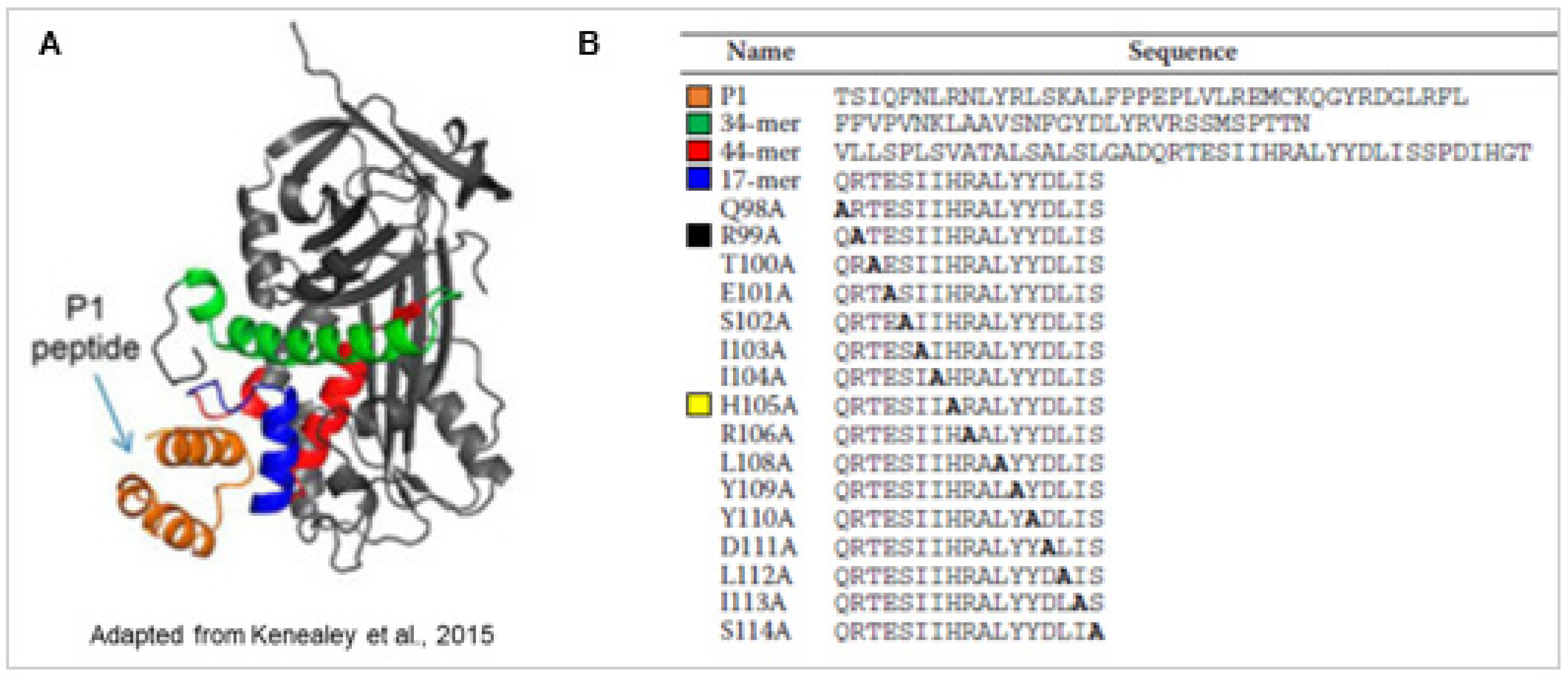
3.2. Structure
3.3. Mechanisms of Action
3.4. Regulation of PEDF Expression
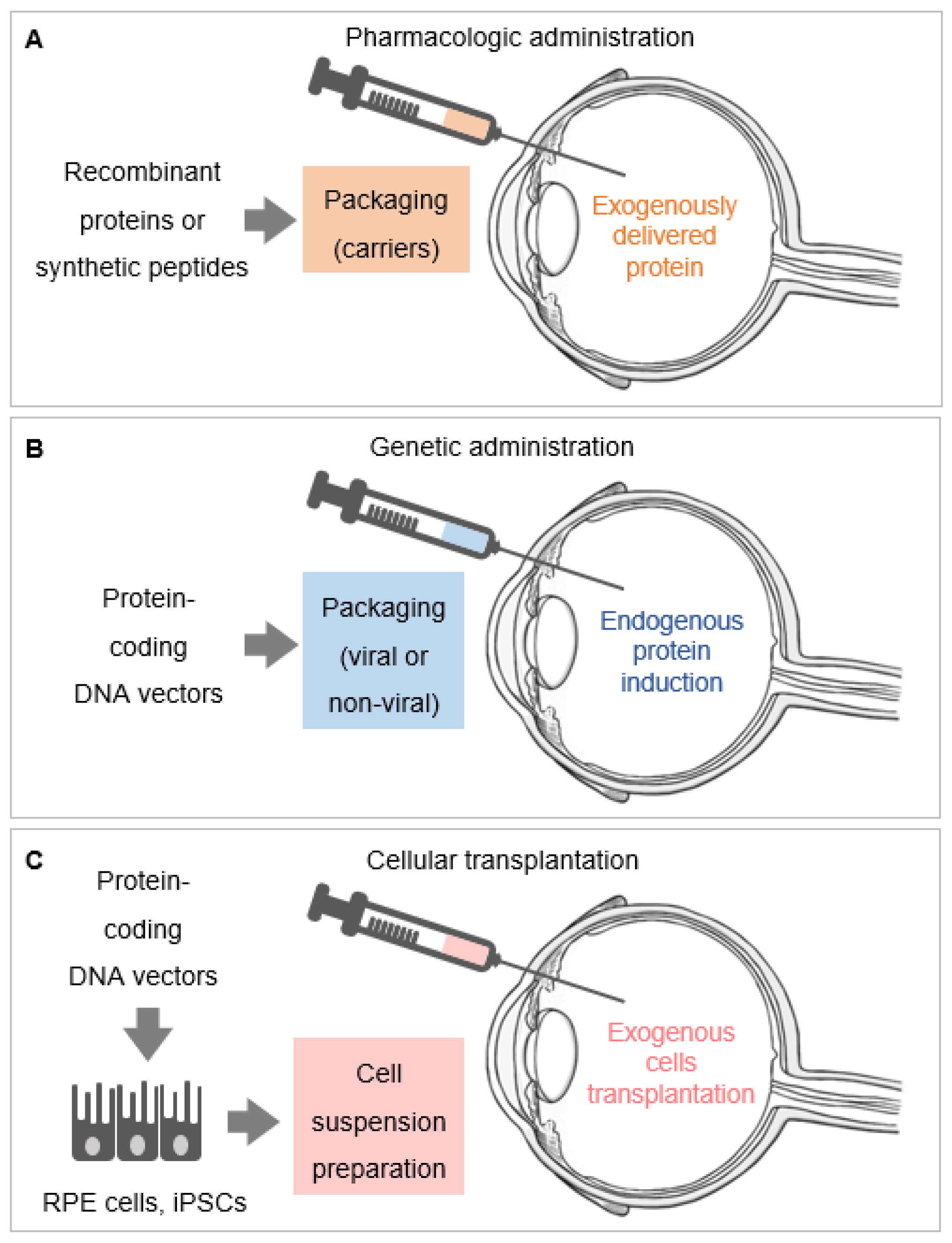
4. PEDF Delivery Systems
4.1. Pharmacological Administration of PEDF
4.2. Genetic Administration of PEDF and Cell-Based Therapies
5. Future Directions for Protein Delivery Systems in Ocular Research
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swaroop, A.; Kim, D.; Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nat. Rev. Neurosci. 2010, 11, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Baden, T.; Euler, T.; Berens, P. Understanding the retinal basis of vision across species. Nat. Rev. Neurosci. 2019, 21, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Sengillo, J.D.; Justus, S.; Tsai, Y.-T.; Cabral, T.; Tsang, S.H. Gene and cell-based therapies for inherited retinal disorders: An update. Am. J. Med. Genet. Part C Semin. Med. Genet. 2016, 172, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Cervia, D.; Catalani, E.; Casini, G. Neuroprotective Peptides in Retinal Disease. J. Clin. Med. 2019, 8, 1146. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Sun, Y.; Cakir, B.; Tomita, Y.; Huang, S.; Wang, Z.; Liu, C.-H.; Cho, S.S.; Britton, W.; Kern, T.S.; et al. Targeting Neurovascular Interaction in Retinal Disorders. Int. J. Mol. Sci. 2020, 21, 1503. [Google Scholar] [CrossRef] [Green Version]
- Arroba, A.I.; Campos-Caro, A.; Aguilar-Diosdado, M.; Valverde, Á.M. IGF-1, Inflammation and Retinal Degeneration: A Close Network. Front. Aging Neurosci. 2018, 10, 203. [Google Scholar] [CrossRef]
- Wyse-Jackson, A.C.; Roche, S.L.; Ruiz-Lopez, A.M.; Moloney, J.N.; Byrne, A.M.; Cotter, T.G. Progesterone analogue protects stressed photoreceptors via bFGF-mediated calcium influx. Eur. J. Neurosci. 2016, 44, 3067–3079. [Google Scholar] [CrossRef]
- Li, S.; Sato, K.; Gordon, W.C.; Sendtner, M.; Bazan, N.G.; Jin, M. Ciliary neurotrophic factor (CNTF) protects retinal cone and rod photoreceptors by suppressing excessive formation of the visual pigments. J. Biol. Chem. 2018, 293, 15256–15268. [Google Scholar] [CrossRef] [Green Version]
- Daly, C.; Shine, L.; Heffernan, T.; Deeti, S.; Reynolds, A.L.; O’Connor, J.J.; Dillon, E.T.; Duffy, D.J.; Kolch, W.; Cagney, G.; et al. A Brain-Derived Neurotrophic Factor Mimetic Is Sufficient to Restore Cone Photoreceptor Visual Function in an Inherited Blindness Model. Sci. Rep. 2017, 7, 11320. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, M.; Mantelli, F.; Rocco, M.L.; Micera, A.; Brandolini, L.; Focareta, L.; Pisano, C.; Aloe, L.; Lambiase, A. Recombinant Human Nerve Growth Factor Treatment Promotes Photoreceptor Survival in the Retinas of Rats with Retinitis Pigmentosa. Curr. Eye Res. 2017, 42, 1064–1068. [Google Scholar] [CrossRef]
- Cachafeiro, M.; Bemelmans, A.-P.; Samardzija, M.; Afanasieva, T.; Pournaras, J.-A.; Grimm, C.; Kostic, C.; Philippe, S.; Wenzel, A.; Arsenijevic, Y. Hyperactivation of retina by light in mice leads to photoreceptor cell death mediated by VEGF and retinal pigment epithelium permeability. Cell Death Dis. 2013, 4, e781. [Google Scholar] [CrossRef]
- Shioda, S.; Takenoya, F.; Wada, N.; Hirabayashi, T.; Seki, T.; Nakamachi, T. Pleiotropic and retinoprotective functions of PACAP. Anat. Sci. Int. 2016, 91, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Akrouh, A.; Kerschensteiner, D. Morphology and function of three VIP-expressing amacrine cell types in the mouse retina. J. Neurophysiol. 2015, 114, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Szabadfi, K.; Pintér, E.; Reglodi, R.; Gábriel, R. Neuropeptides, Trophic Factors, and Other Substances Providing Morphofunctional and Metabolic Protection in Experimental Models of Diabetic Retinopathy. Int. Rev. Cell Mol. Biol. 2014, 311, 1–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sall, J.W.; Klisovic, D.D.; O’Dorisio, M.; Katz, S.E. Somatostatin inhibits IGF-1 mediated induction of VEGF in human retinal pigment epithelial cells. Exp. Eye Res. 2004, 79, 465–476. [Google Scholar] [CrossRef]
- Fernández-Sánchez, L.; Lax, P.; Isiegas, C.; Ayuso, E.; Ruiz, J.M.; De La Villa, P.; Bosch, F.; De La Rosa, E.J.; Cuenca, N. Proinsulin Slows Retinal Degeneration and Vision Loss in the P23H Rat Model of Retinitis Pigmentosa. Hum. Gene Ther. 2012, 23, 1290–1300. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, S.; Barhoum, R.; Boya, P.; Arroba, A.I.; Rodríguez-Muela, N.; Gómez-Vicente, V.; Bosch, F.; de Pablo, F.; de la Villa, P.; de la Rosa, E.J. Attenuation of Vision Loss and Delay in Apoptosis of Photoreceptors Induced by Proinsulin in a Mouse Model of Retinitis Pigmentosa. Investig. Opthalmology Vis. Sci. 2008, 49, 4188–4194. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Li, F.; Steinberg, R.H.; Lavail, M.M. Development of Normal and Injury-induced Gene Expression of aFGF, bFGF, CNTF, BDNF, GFAP and IGF-I in the Rat Retina. Exp. Eye Res. 2001, 72, 591–604. [Google Scholar] [CrossRef]
- West, E.L.; Pearson, R.A.; Duran, Y.; Cordero, A.G.; MacLaren, R.E.; Smith, A.J.; Sowden, J.C.; Ali, R.R. Manipulation of the Recipient Retinal Environment by Ectopic Expression of Neurotrophic Growth Factors Can Improve Transplanted Photoreceptor Integration and Survival. Cell Transplant. 2012, 21, 871–887. [Google Scholar] [CrossRef] [Green Version]
- Rai, U.; Thrimawithana, T.R.; Valery, C.; Young, S.A. Therapeutic uses of somatostatin and its analogues: Current view and potential applications. Pharmacol. Ther. 2015, 152, 98–110. [Google Scholar] [CrossRef]
- Atlasz, T.; Werling, D.; Song, S.; Szabo, E.; Vaczy, A.; Kovari, P.; Tamas, A.; Reglodi, D.; Yu, R. Retinoprotective Effects of TAT-Bound Vasoactive Intestinal Peptide and Pituitary Adenylate Cyclase Activating Polypeptide. J. Mol. Neurosci. 2018, 68, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himawan, E.; Ekström, P.; Buzgo, M.; Gaillard, P.; Stefánsson, E.; Marigo, V.; Loftsson, T.; Paquet-Durand, F. Drug delivery to retinal photoreceptors. Drug Discov. Today 2019, 24, 1637–1643. [Google Scholar] [CrossRef]
- Punzo, C.; Kornacker, K.; Cepko, C.L. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nat. Neurosci. 2008, 12, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polato, F.; Becerra, S.P. Pigment Epithelium-Derived Factor, a Protective Factor for Photoreceptors in Vivo. Adv. Exp. Med. Biol. 2016, 854, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Pagan-Mercado, G.; Becerra, S.P. Signaling Mechanisms Involved in PEDF-Mediated Retinoprotection. Adv. Exp. Med. Biol. 2019, 1185, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Liu, J.; Pu, J.; Milne, G.; Chen, M.; Xu, H.; Shipley, A.; Forrester, J.V.; McCaig, C.D.; Lois, N. Polarized retinal pigment epithelium generates electrical signals that diminish with age and regulate retinal pathology. J. Cell. Mol. Med. 2018, 22, 5552–5564. [Google Scholar] [CrossRef]
- Fudalej, E.; Justyniarska, M.; Kasarełło, K.; Dziedziak, J.; Szaflik, J.P.; Cudnoch-Jędrzejewska, A. Neuroprotective factors of the retina and their role in promoting survival of retinal ganglion cells: A review. Ophthalmic Res. 2021. [Google Scholar] [CrossRef]
- Becerra, S.P.; Notario, V. The effects of PEDF on cancer biology: Mechanisms of action and therapeutic potential. Nat. Rev. Cancer 2013, 13, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Polato, F.; Samardzija, M.; Abu-Asab, M.; Grimm, C.; Crawford, S.E.; Becerra, S.P. PEDF deficiency increases the susceptibility of rd10 mice to retinal degeneration. Exp. Eye Res. 2020, 198, 108121. [Google Scholar] [CrossRef]
- Notari, L.; Baladron, V.; Aroca-Aguilar, J.D.; Balko, N.; Heredia, R.; Meyer, C.; Notario, P.M.; Saravanamuthu, S.; Nueda, M.-L.; Sanchez-Sanchez, F.; et al. Identification of a Lipase-linked Cell Membrane Receptor for Pigment Epithelium-derived Factor. J. Biol. Chem. 2006, 281, 38022–38037. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, P.; Notario, P.M.; Becerra, S.P. Pigment epithelium-derived factor receptor (PEDF-R): A plasma membrane-linked phospholipase with PEDF binding affinity. Single Mol. Single Cell Seq. 2009, 664, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, P.; Locatelli-Hoops, S.; Kenealey, J.; DesJardin, J.; Notari, L.; Becerra, S.P. Pigment Epithelium-derived Factor (PEDF) Prevents Retinal Cell Death via PEDF Receptor (PEDF-R). J. Biol. Chem. 2013, 288, 23928–23942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenealey, J.; Subramanian, P.; Comitato, A.; Bullock, J.; Keehan, L.; Polato, F.; Hoover, D.; Marigo, V.; Becerra, S.P. Small Retinoprotective Peptides Reveal a Receptor-binding Region on Pigment Epithelium-derived Factor. J. Biol. Chem. 2015, 290, 25241–25253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Pinto, A.; Polato, F.; Subramanian, P.; de la Rocha-Muñoz, A.; Vitale, S.; de la Rosa, E.J.; Becerra, S.P. PEDF peptides promote photoreceptor survival in rd10 retina models. Exp. Eye Res. 2019, 184, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Winokur, P.N.; Subramanian, P.; Bullock, J.L.; Arocas, V.; Becerra, S.P. Comparison of two neurotrophic serpins reveals a small fragment with cell survival activity. Mol. Vis. 2017, 23, 372–384. [Google Scholar]
- Kawahara, K.; Yoshida, T.; Maruno, T.; Oki, H.; Ohkubo, T.; Koide, T.; Kobayashi, Y. Spatiotemporal regulation of PEDF signaling by type I collagen remodeling. Proc. Natl. Acad. Sci. USA 2020, 117, 11450–11458. [Google Scholar] [CrossRef]
- García-Onrubia, L.; Valentín-Bravo, F.; Coco-Martin, R.; González-Sarmiento, R.; Pastor, J.; Usategui-Martín, R.; Pastor-Idoate, S. Matrix Metalloproteinases in Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2020, 21, 5934. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, J.; Geng, H.; Li, L.; Li, J.; Cheng, B.; Ma, X.; Li, H.; Hou, L. Photoreceptor degeneration in microphthalmia (Mitf) mice: Partial rescue by pigment epithelium-derived factor. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Valiente-Soriano, F.J.; Di Pierdomenico, J.; García-Ayuso, D.; Ortín-Martínez, A.; De Imperial-Ollero, J.A.M.; Gallego-Ortega, A.; Jiménez-López, M.; Villegas-Pérez, M.P.; Becerra, S.P.; Vidal-Sanz, M. Pigment Epithelium-Derived Factor (PEDF) Fragments Prevent Mouse Cone Photoreceptor Cell Loss Induced by Focal Phototoxicity In Vivo. Int. J. Mol. Sci. 2020, 21, 7242. [Google Scholar] [CrossRef]
- Comitato, A.; Subramanian, P.; Turchiano, G.; Montanari, M.; Becerra, S.P.; Marigo, V. Pigment epithelium-derived factor hinders photoreceptor cell death by reducing intracellular calcium in the degenerating retina. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.L.; He, J.; Kakazu, A.H.; Jun, B.; Bazan, N.G.; Bazan, H.E.P. Defining a mechanistic link between pigment epithelium–derived factor, docosahexaenoic acid, and corneal nerve regeneration. J. Biol. Chem. 2017, 292, 18486–18499. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.-I.; Yu, S.-H.; Chu, H.-S.; Huang, C.-T.; Tsao, Y.-P.; Cheng, C.-M.; Chen, W.-L. Pigment Epithelium-Derived Factor Peptide Promotes Corneal Nerve Regeneration: An In Vivo and In Vitro Study. Investig. Opthalmol. Vis. Sci. 2021, 62, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Subramanian, P.; Shen, D.; Tuo, J.; Becerra, S.P.; Chan, C.-C. Pigment Epithelium-Derived Factor Reduces Apoptosis and Pro-Inflammatory Cytokine Gene Expression in a Murine Model of Focal Retinal Degeneration. ASN Neuro 2013, 5, e00126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holekamp, N.M.; Bouck, N.P.; Volpert, O.V. Pigment epithelium-derived factor is deficient in the vitreous of patients with choroidal neovascularization due to age-related macular degeneration. Am. J. Ophthalmol. 2002, 134, 220–227. [Google Scholar] [CrossRef]
- Ogata, N.; Nishikawa, M.; Nishimura, T.; Mitsuma, Y.; Matsumura, M. Unbalanced vitreous levels of pigment epithelium-derived factor and vascular endothelial growth factor in diabetic retinopathy. Am. J. Ophthalmol. 2002, 134, 348–353. [Google Scholar] [CrossRef]
- Ogata, N.; Matsuoka, M.; Imaizumi, M.; Arichi, M.; Matsumura, M. Decreased levels of pigment Epithelium–derived factor in eyes with neuroretinal dystrophic diseases. Am. J. Ophthalmol. 2004, 137, 1129–1130. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Fariss, R.N.; Wu, Y.Q.; Montuenga, L.M.; Wong, P.; Pfeffer, B.A. Pigment epithelium-derived factor in the monkey retinal pigment epithelium and interphotoreceptor matrix: Apical secretion and distribution. Exp. Eye Res. 2004, 78, 223–234. [Google Scholar] [CrossRef]
- Samuel, W.; Jaworski, C.; Postnikova, O.A.; Kutty, R.K.; Duncan, T.; Tan, L.X.; Poliakov, E.; Lakkaraju, A.; Redmond, T.M. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol. Vis. 2017, 23, 60–89. [Google Scholar]
- Choudhary, P.; Booth, H.; Gutteridge, A.; Surmacz, B.; Louca, I.; Steer, J.; Kerby, J.; Whiting, P.J. Directing Differentiation of Pluripotent Stem Cells Toward Retinal Pigment Epithelium Lineage. STEM CELLS Transl. Med. 2016, 6, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.W.J.; Wang, J.; Zheng, L.; Wang, M.; Lu, Y.; Li, Z.; Lian, C.; Mao, S.; Hou, X.; Li, S.; et al. miR-25 mediates retinal degeneration via inhibiting ITGAV and PEDF in rat. Curr. Mol. Med. 2018, 17, 359–374. [Google Scholar] [CrossRef]
- Zhang, T.; Yin, P.; Zhang, Z.; Xu, B.; Che, D.; Dai, Z.; Dong, C.; Jiang, P.; Hong, H.; Yang, Z.; et al. Deficiency of pigment epithelium-derived factor in nasopharyngeal carcinoma cells triggers the epithelial–mesenchymal transition and metastasis. Cell Death Dis. 2017, 8, e2838. [Google Scholar] [CrossRef]
- Castro-Balado, A.; Mondelo-García, C.; Zarra-Ferro, I.; Fernández-Ferreiro, A. New ophthalmic drug delivery systems. Farm Hosp. 2020, 44, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.-J.; Huang, L.-Z.; Xu, X.-L.; Du, W.; Zhou, A.-Y.; Yu, W.-Z.; Li, X.-X. Polyethylene Glycol-Modified Pigment Epithelial-Derived Factor: New Prospects for Treatment of Retinal Neovascularization. J. Pharmacol. Exp. Ther. 2012, 342, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, X.; Ren, Y.; Liu, Y.; Han, S.; Zhao, J.; Gou, X.; He, Y. PEDF protects human retinal pigment epithelial cells against oxidative stress via upregulation of UCP2 expression. Mol. Med. Rep. 2018, 19, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Sheibani, N.; Wang, S.; Darjatmoko, S.R.; Fisk, D.L.; Shahi, P.K.; Pattnaik, B.R.; Sorenson, C.M.; Bhowmick, R.; Volpert, O.V.; Albert, D.M.; et al. Novel anti-angiogenic PEDF-derived small peptides mitigate choroidal neovascularization. Exp. Eye Res. 2019, 188, 107798. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Roh, K.-H.; Kim, S.W.; Park, S.J.; Lim, N.-Y.; Jung, H.; Choi, I.-W.; Park, S. Type I pig collagen enhances the efficacy of PEDF 34-mer peptide in a mouse model of laser-induced choroidal neovascularization. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Calado, S.M.; Diaz-Corrales, F.J.; Silva, G.A.; Calado, M.S.M. pEPito-driven PEDF Expression Ameliorates Diabetic Retinopathy Hallmarks. Hum. Gene Ther. Methods 2016, 27, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.M.; Zhang, Z.R.; Zhang, Y.G.; Gao, Y.S. Neural Stem Cell-based Intraocular Administration of Pigment Epithelium-derived Factor Promotes Retinal Ganglion Cell Survival and Axon Regeneration after Optic Nerve Crush Injury in Rat: An Experimental Study. Iran. J. Med. Sci. 2016, 41, 382. [Google Scholar]
- Nascimento-Dos-Santos, G.; Teixeira-Pinheiro, L.C.; Da Silva-Júnior, A.J.; De Carvalho, L.R.P.; Mesentier-Louro, L.A.; Hauswirth, W.W.; Mendez-Otero, R.; Santiago, M.F.; Petrs-Silva, H. Effects of a combinatorial treatment with gene and cell therapy on retinal ganglion cell survival and axonal outgrowth after optic nerve injury. Gene Ther. 2019, 27, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.; Park, S.H.; Lee, D.; Kim, G.H.; Noh, J.E.; Lee, K.J.; Kim, G.J. Overexpression of pigment epithelium-derived factor in placenta-derived mesenchymal stem cells promotes mitochondrial biogenesis in retinal cells. Lab. Investig. 2021, 101, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Askou, A.L.; Alsing, S.; Benckendorff, J.N.; Holmgaard, A.; Mikkelsen, J.G.; Aagaard, L.; Bek, T.; Corydon, T.J. Suppression of Choroidal Neovascularization by AAV-Based Dual-Acting Antiangiogenic Gene Therapy. Mol. Ther. Nucleic Acids 2019, 16, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Garcia, L.; Recalde, S.; Hernandez, M.; Bezunartea, J.; Rodriguez-Madoz, J.R.; Johnen, S.; Diarra, S.; Marie, C.; Izsvák, Z.; Ivics, Z.; et al. Long-Term PEDF Release in Rat Iris and Retinal Epithelial Cells after Sleeping Beauty Transposon-Mediated Gene Delivery. Mol. Ther. Nucleic Acids 2017, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.-J.; Mo, B.; Liu, L.; Yue, Y.-K.; Yue, C.-L.; Liu, W. Inhibition of choroidal neovascularization by lentivirus-mediated PEDF gene transfer in rats. Int. J. Ophthalmol. 2016, 9, 1112–1120. [Google Scholar] [CrossRef]
- Sabel, B.A.; You, Q. Nanoparticles as a tool to deliver drugs to the retina and brain: An update. Neural Regen. Res. 2021, 16, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Kang-Mieler, J.J.; Rudeen, K.M.; Liu, W.; Mieler, W.F. Advances in ocular drug delivery systems. Eye 2020, 34, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- DeJulius, C.; Bernardo-Colón, A.; Naguib, S.; Backstrom, J.; Kavanaugh, T.; Gupta, M.; Duvall, C.; Rex, T. Microsphere antioxidant and sustained erythropoietin-R76E release functions cooperate to reduce traumatic optic neuropathy. J. Control. Release 2021, 329, 762–773. [Google Scholar] [CrossRef]
- Üstündağ Okur, N.; Çağlar,, E.Ş.; Siafaka, P.I. Novel Ocular Drug Delivery Systems: An Update on Microemulsions. J. Ocul. Pharmacol. Ther. 2020, 36, 342–354. [Google Scholar] [CrossRef]
- Dubald, M.; Bourgeois, S.; Andrieu, V.; Fessi, H. Ophthalmic Drug Delivery Systems for Antibiotherapy—A Review. Pharmaceutics 2018, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.R.; Kondiah, P.P.D.; Choonara, Y.E.; Du Toit, L.C.; Ally, N.; Pillay, V. Hydrogel Biomaterials for Application in Ocular Drug Delivery. Front. Bioeng. Biotechnol. 2020, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Maguire, A.M.; Bennett, J.; Aleman, E.M.; Leroy, B.P.; Aleman, T.S. Clinical Perspective: Treating RPE65-Associated Retinal Dystrophy. Mol. Ther. 2021, 29, 442–463. [Google Scholar] [CrossRef]
- Chamberlain, K.; Riyad, J.M.; Weber, T. Expressing Transgenes That Exceed the Packaging Capacity of Adeno-Associated Virus Capsids. Hum. Gene Ther. Methods 2016, 27, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Backstrom, J.R.; Sheng, J.; Wang, M.C.; Bernardo-Colón, A.; Rex, T.S. Optimization of S. aureus dCas9 and CRISPRi Elements for a Single Adeno-Associated Virus that Targets an Endogenous Gene. Mol. Ther. Methods Clin. Dev. 2020, 19, 139–148. [Google Scholar] [CrossRef]
- Latella, M.C.; Di Salvo, M.T.; Cocchiarella, F.; Benati, D.; Grisendi, G.; Comitato, A.; Marigo, V.; Recchia, A. In vivo Editing of the Human Mutant Rhodopsin Gene by Electroporation of Plasmid-based CRISPR/Cas9 in the Mouse Retina. Mol. Ther. Nucleic Acids 2016, 5, e389. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarzebska, N.T.; Lauchli, S.; Iselin, C.; French, L.E.; Johansen, P.; Guenova, E.; Kündig, T.M.; Pascolo, S. Functional differences between protamine preparations for the transfection of mRNA. Drug Deliv. 2020, 27, 1231–1235. [Google Scholar] [CrossRef]
- Boehm, B.O.; Lang, G.; Volpert, O.; Jehle, P.M.; Kurkhaus, A.; Rosinger, S.; Bouck, N. Low content of the natural ocular anti-angiogenic agent pigment epithelium-derived factor (PEDF) in aqueous humor predicts progression of diabetic retinopathy. Diabetologia 2003, 46, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, N.; Xu, B.; Wu, M.; Yan, X.; Zhong, L.; Cai, H.; Wang, T.; Wang, Q.; Long, F.; et al. Polymer-based nanoparticles for chemo/gene-therapy: Evaluation its therapeutic efficacy and toxicity against colorectal carcinoma. Biomed. Pharmacother. 2019, 118, 109257. [Google Scholar] [CrossRef]
- Ma, B.; Zhou, Y.; Liu, R.; Zhang, K.; Yang, T.; Hu, C.; Gao, Y.; Lan, Q.; Liu, Y.; Yang, X.; et al. Pigment epithelium-derived factor (PEDF) plays anti-inflammatory roles in the pathogenesis of dry eye disease. Ocul. Surf. 2021, 20, 70–85. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Lu, X.; Tombran-Tink, J.; Zhao, S. PEDF Attenuates Ocular Surface Damage in Diabetic Mice Model Through Its Antioxidant Properties. Curr. Eye Res. 2021, 46, 302–308. [Google Scholar] [CrossRef]
- Brook, N.; Brook, E.; Dharmarajan, A.; Chan, A.; Dass, C.R. The role of pigment epithelium-derived factor in protecting against cellular stress. Free. Radic. Res. 2019, 53, 1166–1180. [Google Scholar] [CrossRef]
- Patel, J.I.; Tombran-Tink, J.; Hykin, P.G.; Gregor, Z.J.; Cree, I.A. Vitreous and aqueous concentrations of proangiogenic, antiangiogenic factors and other cytokines in diabetic retinopathy patients with macular edema: Implications for structural differences in macular profiles. Exp. Eye Res. 2006, 82, 798–806. [Google Scholar] [CrossRef]
- Amaral, J.; Fariss, R.N.; Campos, M.M.; Robison, W.G.; Kim, H.; Lutz, R.; Becerra, S.P. Transscleral-RPE Permeability of PEDF and Ovalbumin Proteins: Implications for Subconjunctival Protein Delivery. Investig. Opthalmo. Vis. Sci. 2005, 46, 4383–4392. [Google Scholar] [CrossRef]
- Cayouette, M.; Smith, S.B.; Becerra, S.; Gravel, C. Pigment Epithelium-Derived Factor Delays the Death of Photoreceptors in Mouse Models of Inherited Retinal Degenerations. Neurobiol. Dis. 1999, 6, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Rieke, E.R.; Amaral, J.; Becerra, S.P.; Lutz, R.J. Sustained Subconjunctival Protein Delivery Using a Thermosetting Gel Delivery System. J. Ocul. Pharmacol. Ther. 2010, 26, 55–64. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Mechanism of Action | Retinoprotective Protein | Abbreviation | Expressing | Target | Ref. |
|---|---|---|---|---|---|
| Cells | Cells | ||||
| Autocrine | insulin-like growth factor 1 | IGF-1 | photoreceptors | photoreceptors | [6] |
| fibroblast growth factor 2 | FGF-2 | photoreceptors | photoreceptors | [7] | |
| ciliary neurotrophic factor | CNTF | photoreceptors | photoreceptors | [8] | |
| brain-derived neurotrophic factor | BDNF | photoreceptors | photoreceptors | [9] | |
| nerve Growth factor | NGF | photoreceptors | photoreceptors | [10] | |
| vascular endothelial growth factor | VEGF | photoreceptors | photoreceptors | [11] | |
| Paracrine | pituitary adenylate cyclase-activating polypeptide | PACAP | retinal cells | photoreceptors | [12] |
| vasoactive intestinal peptide | VIP | (except photoreceptors) | photoreceptors | [13] | |
| insulin-like growth factor 1 | IGF-1 | amacrine cells | photoreceptors | [6] | |
| somatostatin, somatotropin release inhibiting factor | SRIF | microglia | photoreceptors | [14] | |
| vascular endothelial growth factor | VEGF | RPE | choroidal endothelium | [15] | |
| Endocrine | proinsulin | pro-INS | skeletal muscle | photoreceptors | [16,17] |
| insulin | INS | none (systemic administration) | photoreceptors | [17] |
| Organism Model | Molecules | Carrier | Route of | Effect | Ref. |
|---|---|---|---|---|---|
| Delivery | |||||
| mouse LED-induced | rhuPEDF | none | intravitreal | photoreceptor survival | [39] |
| Phototoxicity | 17-mer 17-mer[H105A] | injection | |||
| mouse Mitf KO-induced microphtalmia | rhuPEDF | none | eye drops | photoreceptor survival | [38] |
| 17-mer | |||||
| mouse retinal | rhuPEDF44-mer | none | intravitreal | photoreceptor survival | [34] |
| degeneration (rd10) | 17-mer | injection | |||
| 17-mer[H105A] | |||||
| mouse retinal degeneration (rd1) | rhuPEDF | none | intravitreal | photoreceptor survival | [40] |
| 17-mer | injection | ||||
| 17-mer[H105A] | |||||
| mouse oxidative stress | rhuPEDF | none | intravitreal | photoreceptor survival | [54] |
| injury (H2O2) | injection | ||||
| rabbit laser-induced | 8-mer (peptide 335, modified from 34-mer) | polymeric | intravitreal | choroidal neovascularization inhibition | [55] |
| choroidal | ester | injection | |||
| neovascularization | prodrug | ||||
| mouse laser-induced | 34-mer | type I | intravitreal | choroidal neovascularization inhibition | [56] |
| choroidal | collagen | injection | |||
| neovascularization | eye drops |
| Organism Model | DNA Vector | Carrier/Host Cells | Route of Delivery | Effect | Ref. |
|---|---|---|---|---|---|
| Ins2Akita diabetic mouse | pEPito-hCMV-PEDF episomal vector | none | subretinal injection | photoreceptors survival | [57] |
| (electroporation) | |||||
| rat optic nerve crush | PEDF-secreting neural stem cell (NSC)-based system, lentivirus | none | subretinal cell transplantation | photoreceptors survival | [58] |
| host cells: human neural stem cells | |||||
| rat optic nerve crush | AAV2.PEDF combined with human mesenchymal stem cell | none | Intravitreal cell transplantation | RGC survival and ON injury | [59] |
| host cells: human mesenchymal stem cells | |||||
| rat H2O2-induced retinal degeneration | Placenta-derived-mesenchymal stem cells overexpressing PEDF plasmid | none | Intravitreal cell transplantation | Mitochondrial biogenesis in RPE cells | [60] |
| host cells: human placental-derived stem cells | |||||
| mouse laser-induced neovascularization | multigenic AAV5.PEDF + multiple miRNAs targeting the VEGF-A gene, p/miR(5,B,7/Irr)-AsR/PEDF-PE | none | Intravitreal injection | Choroidal cells inhibition | [61] |
| rat choroidal neovascularization | IPE or RPE cells with pFAR4-ITRs CMV PEDF BGH plasmid, sleeping beauty transposon | none | Subretinal cell transplantation | choroidal cells inhibition | [62] |
| (electroporation) | |||||
| host cells: primary rat RPE cells | |||||
| rat choroidal neovascularization | Lentivirus-PEDF-green fluorescent protein (GFP) | none | Intravitreal injection | choroidal cells inhibition | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebustini, I.T.; Bernardo-Colón, A.; Nalvarte, A.I.; Becerra, S.P. Delivery Systems of Retinoprotective Proteins in the Retina. Int. J. Mol. Sci. 2021, 22, 5344. https://doi.org/10.3390/ijms22105344
Rebustini IT, Bernardo-Colón A, Nalvarte AI, Becerra SP. Delivery Systems of Retinoprotective Proteins in the Retina. International Journal of Molecular Sciences. 2021; 22(10):5344. https://doi.org/10.3390/ijms22105344
Chicago/Turabian StyleRebustini, Ivan T., Alexandra Bernardo-Colón, Alejandra Isasi Nalvarte, and S. Patricia Becerra. 2021. "Delivery Systems of Retinoprotective Proteins in the Retina" International Journal of Molecular Sciences 22, no. 10: 5344. https://doi.org/10.3390/ijms22105344
APA StyleRebustini, I. T., Bernardo-Colón, A., Nalvarte, A. I., & Becerra, S. P. (2021). Delivery Systems of Retinoprotective Proteins in the Retina. International Journal of Molecular Sciences, 22(10), 5344. https://doi.org/10.3390/ijms22105344