
The Meiosis-Specific Crs1 Cyclin Is Required for Efficient S-Phase Progression and Stable Nuclear Architecture


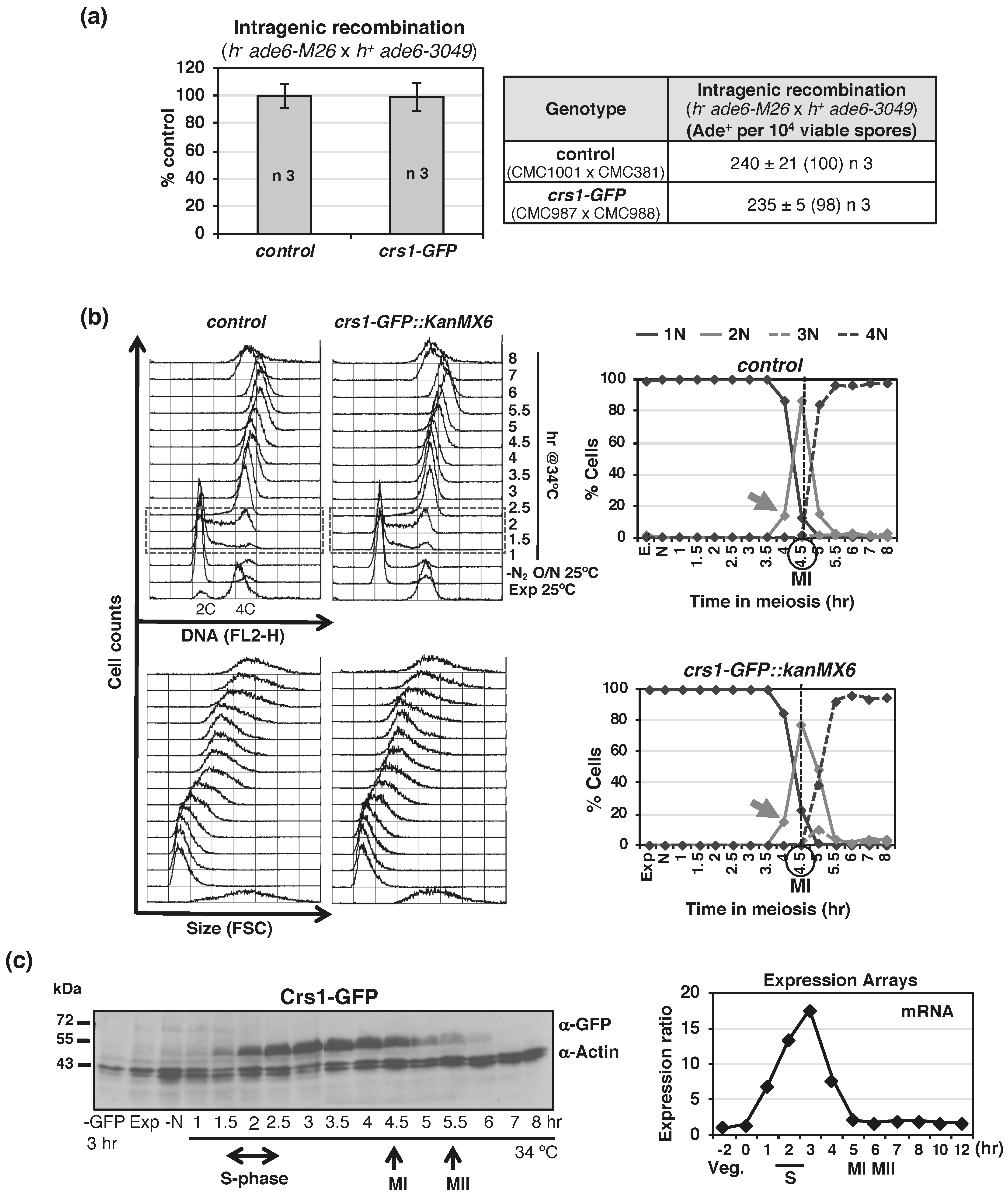{kind=link}
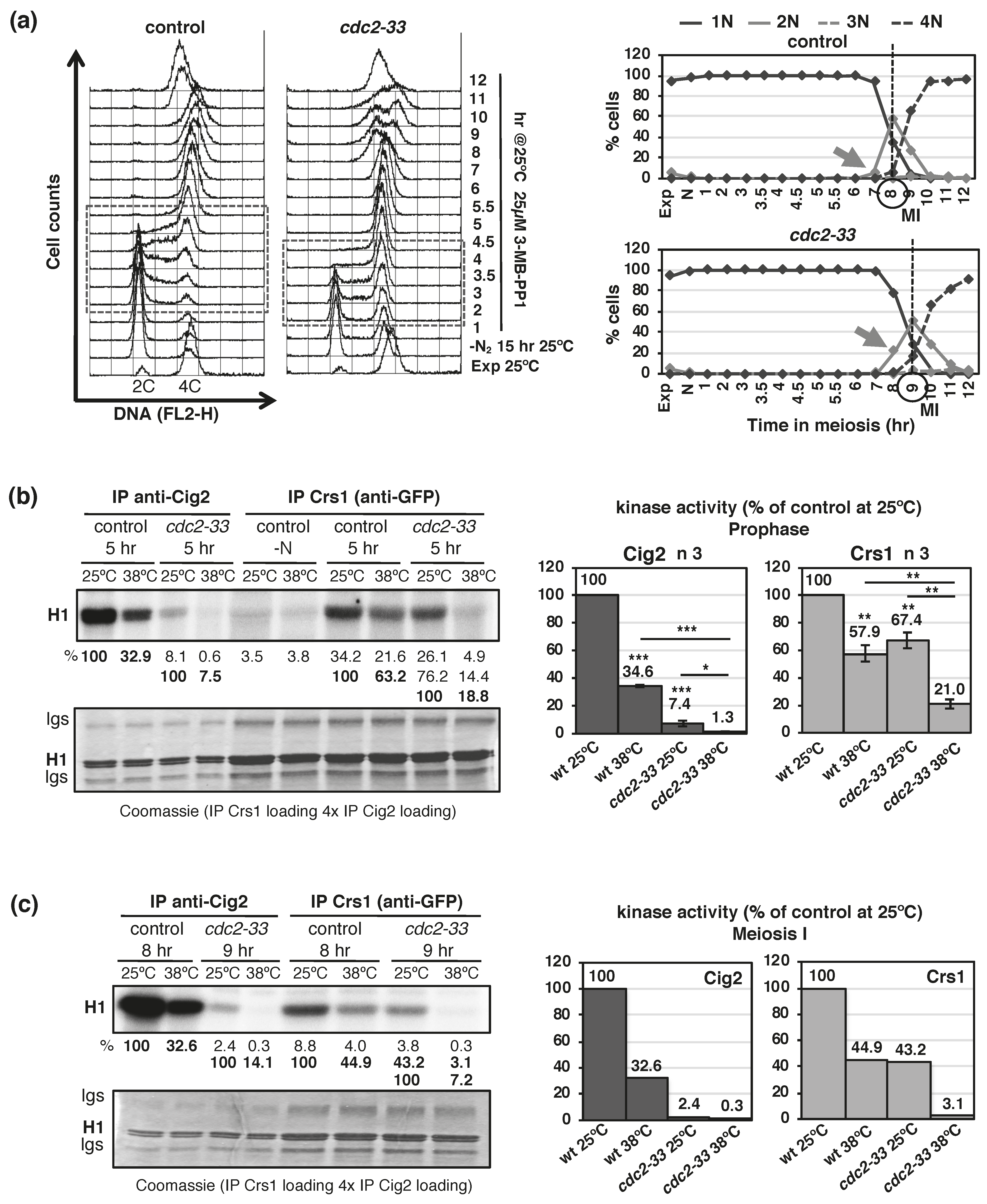{kind=link}
{kind=link}
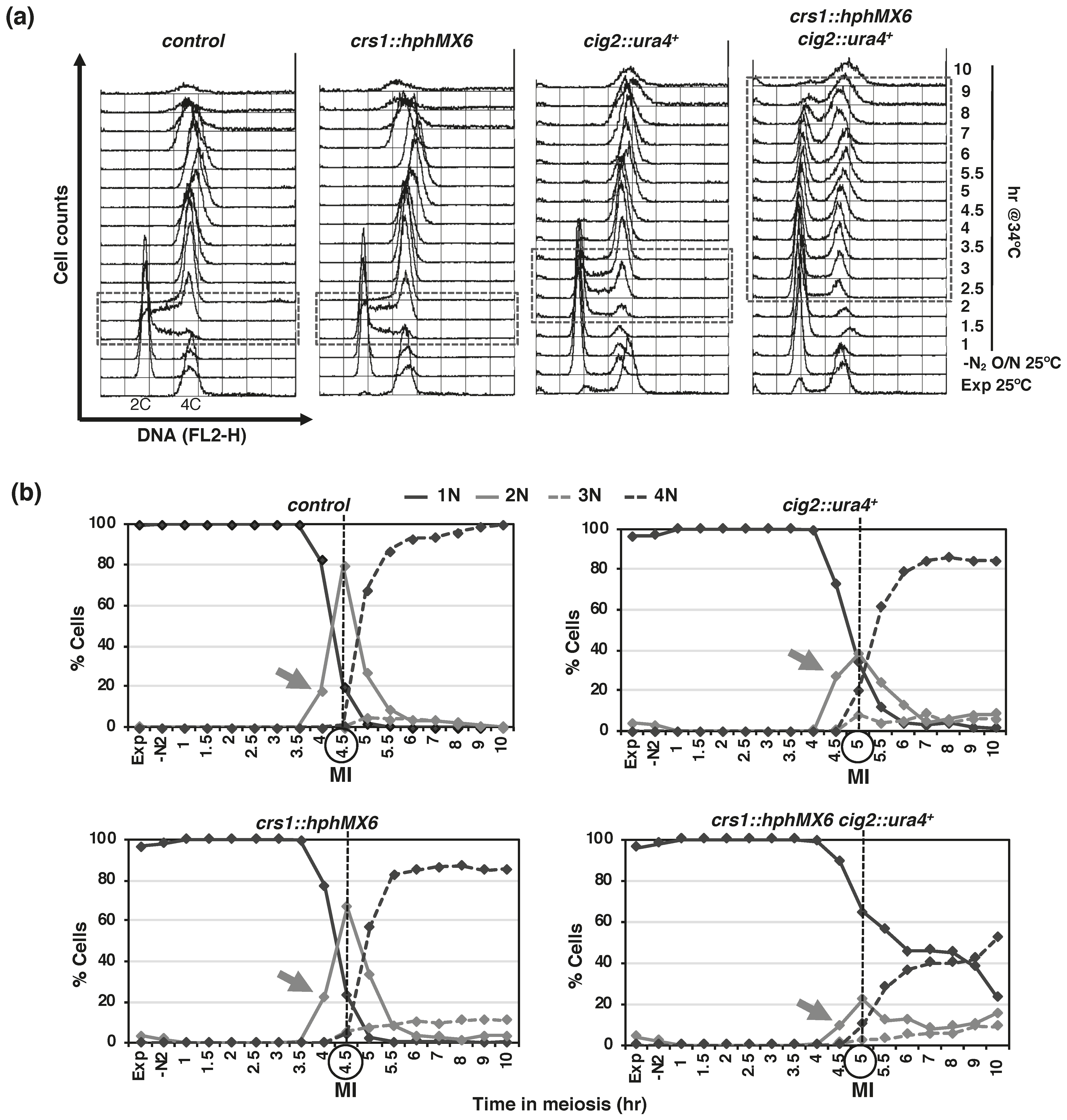{kind=link}
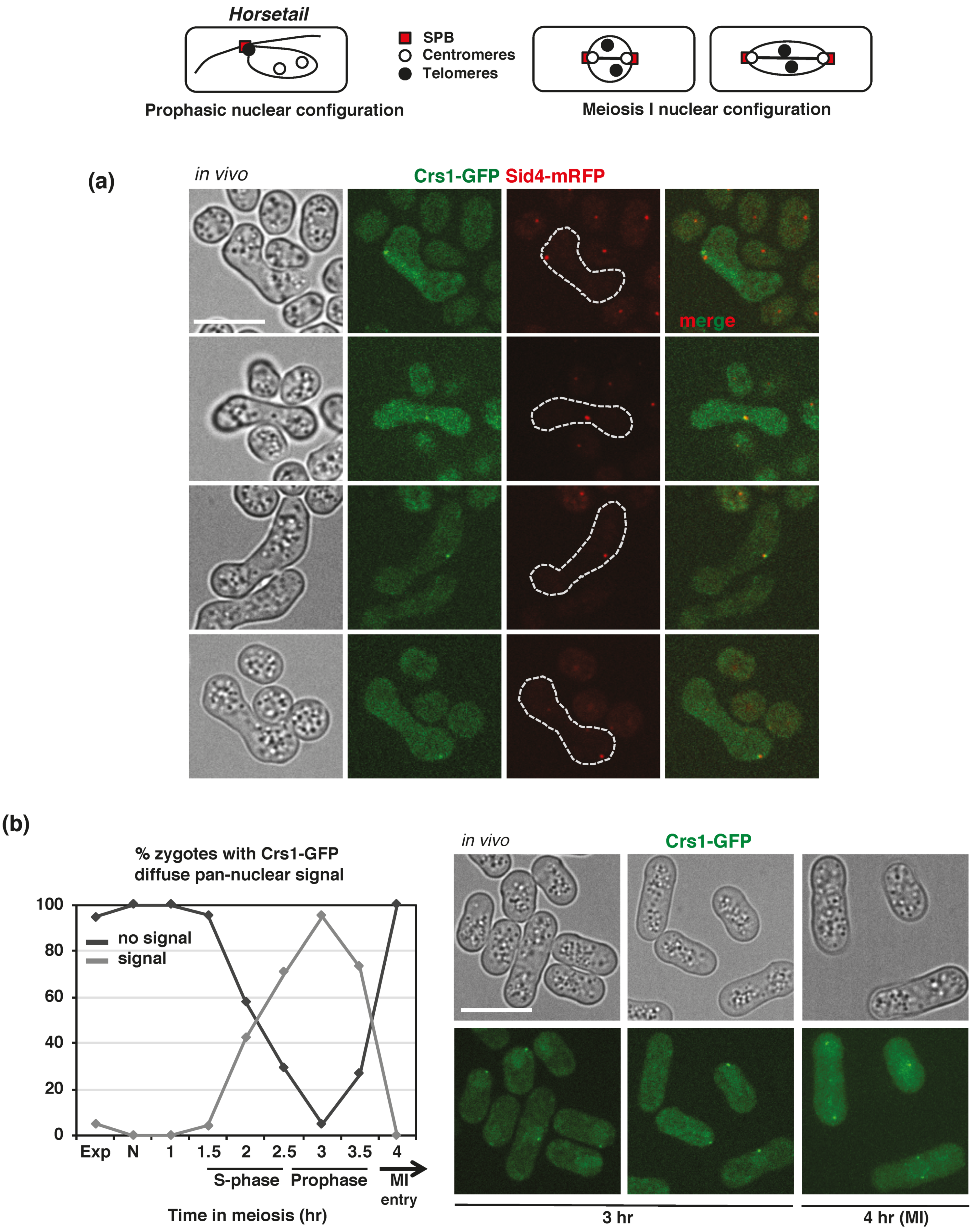{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Yeast Manipulation and General Methods
4.2. Crs1-GFP Fusion
4.3. Kinase Assays
4.4. Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stern, B.; Nurse, P. A quantitative model for the cdc2 control of S phase and mitosis in fission yeast. Trends Genet. 1996, 12, 345–350. [Google Scholar] [CrossRef]
- Uhlmann, F.; Bouchoux, C.; López-Avilés, S. A quantitative model for cyclin-dependent kinase control of the cell cycle: Revisited. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3572–3583. [Google Scholar] [CrossRef] [Green Version]
- Coudreuse, D.; Nurse, P. Driving the cell cycle with a minimal CDK control network. Nat. Cell Biol. 2010, 468, 1074–1079. [Google Scholar] [CrossRef]
- Gutiérrez-Escribano, P.; Nurse, P. A single cyclin—CDK complex is sufficient for both mitotic and meiotic progression in fission yeast. Nat. Commun. 2015, 6, 6871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewald, J.C.; Kuehne, A.; Zamboni, N.; Skotheim, J.M. The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression. Mol. Cell 2016, 62, 532–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurat, C.F.; Wolinski, H.; Petschnigg, J.; Kaluarachchi, S.; Andrews, B.; Natter, K.; Kohlwein, S.D. Cdk1/Cdc28-Dependent Activation of the Major Triacylglycerol Lipase Tgl4 in Yeast Links Lipolysis to Cell-Cycle Progression. Mol. Cell 2009, 33, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Chen, Y.; Carey, L.; Futcher, B. Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae. Mol. Cell 2016, 62, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Trovesi, C.; Manfrini, N.; Falcettoni, M.; Longhese, M.P. Regulation of the DNA Damage Response by Cyclin-Dependent Kinases. J. Mol. Biol. 2013, 425, 4756–4766. [Google Scholar] [CrossRef]
- Hentges, P.; Waller, H.; Reis, C.C.; Ferreira, M.G.; Doherty, A.J. Cdk1 Restrains NHEJ through Phosphorylation of XRCC4-like Factor Xlf1. Cell Rep. 2014, 9, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Dehé, P.-M.; Coulon, S.; Scaglione, S.; Shanahan, P.; Takedachi, A.; A Wohlschlegel, J.; Yates, J.R.; Llorente, B.; Russell, P.; Gaillard, P.-H.L. Regulation of Mus81–Eme1 Holliday junction resolvase in response to DNA damage. Nat. Struct. Mol. Biol. 2013, 20, 598–603. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Niu, H.; Yu, Y.; Wang, J.; Zhu, S.; Zhou, J.; Papusha, A.; Cui, D.; Pan, X.; Kwon, Y.; et al. Enrichment of Cdk1-cyclins at DNA double-strand breaks stimulates Fun30 phosphorylation and DNA end resection. Nucleic Acids Res. 2016, 44, 2742–2753. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, S.; Mielnichuk, N.; Pérez-Martín, J. Programmed cell cycle arrest is required for infection of corn plants by the fungus Ustilago maydis. Development 2014, 141, 4817–4826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Esteban, M.; Garcia-Higuera, I.; Maestre, C.; Moreno, S.; Almeida, A. APC/C-Cdh1 coordinates neurogenesis and cortical size during development. Nat. Commun. 2013, 4, 2879. [Google Scholar] [CrossRef] [Green Version]
- Henderson, K.A.; Kee, K.; Maleki, S.; Santini, P.A.; Keeney, S. Cyclin-Dependent Kinase Directly Regulates Initiation of Meiotic Recombination. Cell 2006, 125, 1321–1332. [Google Scholar] [CrossRef] [Green Version]
- Chung, G.; Rose, A.M.; Petalcorin, M.I.; Martin, J.S.; Kessler, Z.; Sanchez-Pulido, L.; Ponting, C.P.; Yanowitz, J.L.; Boulton, S.J. REC-1 and HIM-5 distribute meiotic crossovers and function redundantly in meiotic double-strand break formation inCaenorhabditis elegans. Genes Dev. 2015, 29, 1969–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante-Jaramillo, L.F.; Ramos, C.; Alonso, L.; Sesmero, A.; Segurado, M.; Martín-Castellanos, C. CDK contribution to DSB formation and recombination in fission yeast meiosis. PLoS Genet. 2019, 15, e1007876. [Google Scholar] [CrossRef] [PubMed]
- Viera, A.; Alsheimer, M.; Gómez, R.; Berenguer, I.; Ortega, S.; Symonds, C.E.; Santamaría, D.; Benavente, R.; Suja, J.A. CDK2 regulates nuclear envelope protein dynamics and telomere attachment in mouse meiotic prophase. J. Cell Sci. 2015, 128, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Martinerie, L.; Manterola, M.; Chung, S.S.W.; Panigrahi, S.K.; Weisbach, M.; Vasileva, A.; Geng, Y.; Sicinski, P.; Wolgemuth, D.J. Mammalian E-type Cyclins Control Chromosome Pairing, Telomere Stability and CDK2 Localization in Male Meiosis. PLoS Genet. 2014, 10, e1004165. [Google Scholar] [CrossRef] [Green Version]
- Manterola, M.; Sicinski, P.; Wolgemuth, D.J. E-type cyclins modulate telomere integrity in mammalian male meiosis. Chromosoma 2015, 125, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Ashley, T.; Walpita, D.; de Rooij, D. Localization of two mammalian cyclin dependent kinases during mammalian meiosis. J. Cell Sci. 2001, 114, 685–693. [Google Scholar] [CrossRef]
- Reynolds, A.; Qiao, H.; Yang, Y.; Chen, J.K.; Jackson, N.; Biswas, K.; Holloway, J.K.; Baudat, F.; De Massy, B.; Wang, J.; et al. RNF212 is a dosage-sensitive regulator of crossing-over during mammalian meiosis. Nat. Genet. 2013, 45, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Rao, H.P.; Yang, Y.; Fong, J.H.; Cloutier, J.M.; Deacon, D.C.; Nagel, K.E.; Swartz, R.K.; Strong, E.; Holloway, J.K.; et al. Antagonistic roles of ubiquitin ligase HEI10 and SUMO ligase RNF212 regulate meiotic recombination. Nat. Genet. 2014, 46, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Holloway, J.K.; Sun, X.; Yokoo, R.; Villeneuve, A.M.; Cohen, P.E. Mammalian CNTD1 is critical for meiotic crossover maturation and deselection of excess precrossover sites. J. Cell Biol. 2014, 205, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Yokoo, R.; Zawadzki, K.A.; Nabeshima, K.; Drake, M.; Arur, S.; Villeneuve, A.M. COSA-1 Reveals Robust Homeostasis and Separable Licensing and Reinforcement Steps Governing Meiotic Crossovers. Cell 2012, 149, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Wijnker, E.; Harashima, H.; Müller, K.; Parra-Nuñez, P.; de Snoo, C.B.; van de Belt, J.; Dissmeyer, N.; Bayer, M.; Pradillo, M.; Schnittger, A. The Cdk1/Cdk2 homolog CDKA.;1 controls the recombination landscape in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 12534–12539. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.B.; Manova, K.; Capodieci, P.; Lindon, C.; Bottega, S.; Wang, X.-Y.; Refik-Rogers, J.; Pines, J.; Wolgemuth, D.J.; Koff, A. Characterization and Expression of Mammalian Cyclin B3, a Prepachytene Meiotic Cyclin. J. Biol. Chem. 2002, 277, 41960–41969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Averbeck, N.; Sunder, S.; Sample, N.; Wise, J.A.; Leatherwood, J. Negative Control Contributes to an Extensive Program of Meiotic Splicing in Fission Yeast. Mol. Cell 2005, 18, 491–498. [Google Scholar] [CrossRef]
- Malapeira, J.; Moldón, A.; Hidalgo, E.; Smith, G.R.; Nurse, P.; Ayté, J. A Meiosis-Specific Cyclin Regulated by Splicing Is Required for Proper Progression through Meiosis. Mol. Cell. Biol. 2005, 25, 6330–6337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, X.; Song, W.; Wang, W.; Gao, S. Cyclin Cyc2p is required for micronuclear bouquet formation in Tetrahymena thermophila. Sci. China Life Sci. 2019, 62, 668–680. [Google Scholar] [CrossRef]
- Ellenrieder, C.; Bartosch, B.; Lee, G.Y.-C.; Murphy, M.; Sweeney, C.; Hergersberg, M.; Carrington, M.; Jaussi, R.; Hunt, T. The Long Form of CDK2 Arises via Alternative Splicing and Forms an Active Protein Kinase with Cyclins A and E. DNA Cell Biol. 2001, 20, 413–423. [Google Scholar] [CrossRef]
- Tu, Z.; Bayazit, M.B.; Liu, H.; Zhang, J.; Busayavalasa, K.; Risal, S.; Shao, J.; Satyanarayana, A.; Coppola, V.; Tessarollo, L.; et al. Speedy A–Cdk2 binding mediates initial telomere–nuclear envelope attachment during meiotic prophase I independent of Cdk2 activation. Proc. Natl. Acad. Sci. USA 2017, 114, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgne, A.; Murakami, H.; Ayté, J.; Nurse, P. The G1/S Cyclin Cig2p during Meiosis in Fission Yeast. Mol. Biol. Cell 2002, 13, 2080–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decottignies, A.; Zarzov, P.; Nurse, P. In vivo localisation of fission yeast cyclin-dependent kinase cdc2p and cyclin B cdc13p during mitosis and meiosis. J. Cell Sci. 2001, 114, 2627–2640. [Google Scholar] [CrossRef]
- Moiseeva, V.; Amelina, H.; Collopy, L.C.; A Armstrong, C.; Pearson, S.R.; Tomita, K. The telomere bouquet facilitates meiotic prophase progression and exit in fission yeast. Cell Discov. 2017, 3, 17041. [Google Scholar] [CrossRef] [Green Version]
- Bass, H.W. Telomere dynamics unique to meiotic prophase: Formation and significance of the bouquet. Cell. Mol. Life Sci. 2003, 60, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Chikashige, Y.; Ding, D.; Imai, Y.; Yamamoto, M.; Haraguchi, T.; Hiraoka, Y. Meiotic nuclear reorganization: Switching the position of centromeres and telomeres in the fission yeast Schizosaccharomyces pombe. EMBO J. 1997, 16, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikashige, Y.; Tsutsumi, C.; Yamane, M.; Okamasa, K.; Haraguchi, T.; Hiraoka, Y. Meiotic Proteins Bqt1 and Bqt2 Tether Telomeres to Form the Bouquet Arrangement of Chromosomes. Cell 2006, 125, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Jin, Y.; Cande, W.Z. Bqt2p is essential for initiating telomere clustering upon pheromone sensing in fission yeast. J. Cell Biol. 2006, 173, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-Q.; Hiraoka, Y. Nuclear movement enforcing chromosome alignment in fission yeast—Meiosis without homolog synapsis. In Recombination and Meiosis; Springer: Berlin/Heidelberg, Germany, 2007; pp. 231–247. [Google Scholar]
- Ruan, K.; Yamamoto, T.G.; Asakawa, H.; Chikashige, Y.; Masukata, H.; Haraguchi, T.; Hiraoka, Y. Meiotic nuclear movements in fission yeast are regulated by the transcription factor Mei4 downstream of a Cds1-dependent replication checkpoint pathway. Genes Cells 2014, 20, 160–172. [Google Scholar] [CrossRef]
- Ding, D.-Q.; Yamamoto, A.; Haraguchi, T.; Hiraoka, Y. Dynamics of Homologous Chromosome Pairing during Meiotic Prophase in Fission Yeast. Dev. Cell 2004, 6, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; Smith, G.R. The Meiotic Bouquet Promotes Homolog Interactions and Restricts Ectopic Recombination in Schizosaccharomyces pombe. Genetics 2006, 174, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, K.; Cooper, J.P. The Telomere Bouquet Controls the Meiotic Spindle. Cell 2007, 130, 113–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klutstein, M.; Fennell, A.; Fernández-Álvarez, A.; Cooper, J.P. The telomere bouquet regulates meiotic centromere assembly. Nat. Cell Biol. 2015, 17, 458–469. [Google Scholar] [CrossRef] [PubMed]
- McPheeters, D.S.; Cremona, N.; Sunder, S.; Chen, H.-M.; Averbeck, N.; Leatherwood, J.; Wise, J.A. A complex gene regulatory mechanism that operates at the nexus of multiple RNA processing decisions. Nat. Struct. Mol. Biol. 2009, 16, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-M.; Futcher, B.; Leatherwood, J. The Fission Yeast RNA Binding Protein Mmi1 Regulates Meiotic Genes by Controlling Intron Specific Splicing and Polyadenylation Coupled RNA Turnover. PLoS ONE 2011, 6, e26804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folco, H.D.; Chalamcharla, V.R.; Sugiyama, T.; Thillainadesan, G.; Zofall, M.; Balachandran, V.; Dhakshnamoorthy, J.; Mizuguchi, T.; Grewal, H. Untimely expression of gametogenic genes in vegetative cells causes uniparental disomy. Nat. Cell Biol. 2017, 543, 126–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iino, Y.; Yamamoto, M. Negative control for the initiation of meiosis in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1985, 82, 2447–2451. [Google Scholar] [CrossRef] [Green Version]
- Lock, A.; Rutherford, K.; A Harris, M.; Hayles, J.; Oliver, S.G.; Bähler, J.; Wood, V. PomBase 2018: User-driven reimplementation of the fission yeast database provides rapid and intuitive access to diverse, interconnected information. Nucleic Acids Res. 2019, 47, D821–D827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booher, R.N.; Alfa, C.E.; Hyams, J.S.; Beach, D.H. The fission yeast cdc2/cdc13/suc1 protein kinase: Regulation of catalytic activity and nuclear localization. Cell 1989, 58, 485–497. [Google Scholar] [CrossRef]
- Moreno, S.; Hayles, J.; Nurse, P. Regulation of p34cdc2 protein kinase during mitosis. Cell 1989, 58, 361–372. [Google Scholar] [CrossRef]
- Nurse, P.; Thuriaux, P.; Nasmyth, K. Genetic control of the cell division cycle in the fission yeast Schizosaccharomyces pombe. Mol. Genet. Genom. 1976, 146, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Moreno, A.; Alves-Rodrigues, I.; Hidalgo, E.; Ayté, J. Chemical genetic induction of meiosis in Schizosaccharomyces pombe. Cell Cycle 2012, 11, 1621–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Castellanos, C.; Labib, K.; Moreno, S. B-type cyclins regulate G1 progression in fission yeast in opposition to the p25rum1 cdk inhibitor. EMBO J. 1996, 15, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Martín-Castellanos, C. Regulación de la progresión por la fase G1 del ciclo celular en Schizosaccharomyces pombe. Ph.D. Thesis, Department of Microbiology and Genetics, University of Salamanca, Salamanca, Spain, 1997. [Google Scholar]
- Obara-Ishihara, T.; Okayama, H. A B-type cyclin negatively regulates conjugation via interacting with cell cycle ‘start’ genes in fission yeast. EMBO J. 1994, 13, 1863–1872. [Google Scholar] [CrossRef]
- Hayashi, A.; Asakawa, H.; Haraguchi, T.; Hiraoka, Y. Reconstruction of the Kinetochore during Meiosis in Fission Yeast Schizosaccharomyces pombe. Mol. Biol. Cell 2006, 17, 5173–5184. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Tabarés, I.; Grallert, A.; Ortiz, J.-M.; Hagan, I.M. Schizosaccharomyces pombe protein phosphatase 1 in mitosis, endocytosis and a partnership with Wsh3/Tea4 to control polarised growth. J. Cell Sci. 2007, 120, 3589–3601. [Google Scholar] [CrossRef] [Green Version]
- Chikashige, Y.; Ding, D.Q.; Funabiki, H.; Haraguchi, T.; Mashiko, S.; Yanagida, M.; Hiraoka, Y. Telomere-led premeiotic chromosome movement in fission yeast. Science 1994, 264, 270–273. [Google Scholar] [CrossRef]
- Chikashige, Y.; Hiraoka, Y. Telomere binding of the Rap1 protein is required for meiosis in fission yeast. Curr. Biol. 2001, 11, 1618–1623. [Google Scholar] [CrossRef] [Green Version]
- West, R.R.; Vaisberg, E.V.; Ding, R.; Nurse, P.; McIntosh, J.R. cut11+: A Gene Required for Cell Cycle-dependent Spindle Pole Body Anchoring in the Nuclear Envelope and Bipolar Spindle Formation in Schizosaccharomyces pombe. Mol. Biol. Cell 1998, 9, 2839–2855. [Google Scholar] [CrossRef] [Green Version]
- Varberg, J.M.; Gardner, J.M.; McCroskey, S.; Saravanan, S.; Bradford, W.D.; Jaspersen, S.L. High-Throughput Identification of Nuclear Envelope Protein Interactions in Schizosaccharomyces pombe Using an Arrayed Membrane Yeast-Two Hybrid Library. G3 Genes Genomes Genet. 2020, 10, 4649–4663. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M. Cyclin-dependent kinases. Genome Biol. 2014, 15, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harashima, H.; Dissmeyer, N.; Schnittger, A. Cell cycle control across the eukaryotic kingdom. Trends Cell Biol. 2013, 23, 345–356. [Google Scholar] [CrossRef]
- Li, J.; Qian, W.-P.; Sun, Q.-Y. Cyclins regulating oocyte meiotic cell cycle progression†. Biol. Reprod. 2019, 101, 878–881. [Google Scholar] [CrossRef]
- Palmer, N.; Talib, S.Z.A.; Kaldis, P. Diverse roles for CDK-associated activity during spermatogenesis. FEBS Lett. 2019, 593, 2925–2949. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, A.M.; Lacefield, S. CDK Regulation of Meiosis: Lessons from S. cerevisiae and S. pombe. Genes 2020, 11, 723. [Google Scholar] [CrossRef]
- Murakami, H.; Nurse, P. Meiotic DNA replication checkpoint control in fission yeast. Genes Dev. 1999, 13, 2581–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlile, T.M.; Amon, A. Meiosis I Is Established through Division-Specific Translational Control of a Cyclin. Cell 2008, 133, 280–291. [Google Scholar] [CrossRef] [Green Version]
- Futcher, B. Cyclins in Meiosis: Lost in Translation. Dev. Cell 2008, 14, 644–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeney, S.; Lange, J.; Mohibullah, N. Self-Organization of Meiotic Recombination Initiation: General Principles and Molecular Pathways. Annu. Rev. Genet. 2014, 48, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Ortega, S.; Prieto, I.; Odajima, J.; Martin, A.; Dubus, P.; Sotillo, R.; Barbero, J.L.; Malumbres, M.; Barbacid, M. Cyclin-dependent kinase 2 is essential for meiosis but not for mitotic cell division in mice. Nat. Genet. 2003, 35, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Viera, A.; Rufas, J.S.; Martínez, I.; Barbero, J.L.; Ortega, S.; Suja, J.A. CDK2 is required for proper homologous pairing, recombination and sex-body formation during male mouse meiosis. J. Cell Sci. 2009, 122, 2149–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikolcevic, P.; Isoda, M.; Shibuya, H.; Barrantes, I.D.B.; Igea, A.; Suja, J.A.; Shackleton, S.; Watanabe, Y.; Nebreda, A.R. Essential role of the Cdk2 activator RingoA in meiotic telomere tethering to the nuclear envelope. Nat. Commun. 2016, 7, 11084. [Google Scholar] [CrossRef] [Green Version]
- Amelina, H.; Subramaniam, S.; Moiseeva, V.; Armstrong, C.A.; Pearson, S.R.; Tomita, K. Telomere protein Rap1 is a charge resistant scaffolding protein in chromosomal bouquet formation. BMC Biol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Fujita, I.; Nishihara, Y.; Tanaka, M.; Tsujii, H.; Chikashige, Y.; Watanabe, Y.; Saito, M.; Ishikawa, F.; Hiraoka, Y.; Kanoh, J. Telomere-Nuclear Envelope Dissociation Promoted by Rap1 Phosphorylation Ensures Faithful Chromosome Segregation. Curr. Biol. 2012, 22, 1932–1937. [Google Scholar] [CrossRef] [Green Version]
- Fennell, A.; Fernández-Álvarez, A.; Tomita, K.; Cooper, J.P. Telomeres and centromeres have interchangeable roles in promoting meiotic spindle formation. J. Cell Biol. 2015, 208, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Álvarez, A.; Bez, C.; O’Toole, E.T.; Morphew, M.; Cooper, J.P. Mitotic Nuclear Envelope Breakdown and Spindle Nucleation Are Controlled by Interphase Contacts between Centromeres and the Nuclear Envelope. Dev. Cell 2016, 39, 544–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, I.M.; Grallert, A. Spatial control of mitotic commitment in fission yeast. Biochem. Soc. Trans. 2013, 41, 1766–1771. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M.; Ferenczy, L. Protoplast fusion of Schizosaccharomyces pombe auxotrophic mutants of identical mating-type. Mol. Genet. Genom. 1977, 151, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.; Rozalén, A.E.; Moreno, S.; Smith, G.R.; Martín-Castellanos, C. Rec25 and Rec27, Novel Linear-Element Components, Link Cohesin to Meiotic DNA Breakage and Recombination. Curr. Biol. 2008, 18, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Foiani, M.; Marini, F.; Gamba, D.; Lucchini, G.; Plevani, P. The B subunit of the DNA polymerase alpha-primase complex in Saccharomyces cerevisiae executes an essential function at the initial stage of DNA replication. Mol. Cell. Biol. 1994, 14, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Bahler, J.; Wu, J.Q.; Longtine, M.S.; Shah, N.G.; McKenzie, A., 3rd; Steever, A.B.; Wach, A.; Philippsen, P.; Pringle, J.R. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 1998, 14, 943–951. [Google Scholar] [CrossRef]
- Benito, J.; Martín-Castellanos, C.; Moreno, S. Regulation of the G1 phase of the cell cycle by periodic stabilization and degradation of the p25rum1 CDK inhibitor. EMBO J. 1998, 17, 482–497. [Google Scholar] [CrossRef] [PubMed]
- Rincon, S.A.; Lamson, A.; Blackwell, R.; Syrovatkina, V.; Fraisier, V.; Paoletti, A.; Betterton, M.D.; Tran, P.T. Kinesin-5-independent mitotic spindle assembly requires the antiparallel microtubule crosslinker Ase1 in fission yeast. Nat. Commun. 2017, 8, 15286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante-Jaramillo, L.F.; Ramos, C.; Martín-Castellanos, C. The Meiosis-Specific Crs1 Cyclin Is Required for Efficient S-Phase Progression and Stable Nuclear Architecture. Int. J. Mol. Sci. 2021, 22, 5483. https://doi.org/10.3390/ijms22115483
Bustamante-Jaramillo LF, Ramos C, Martín-Castellanos C. The Meiosis-Specific Crs1 Cyclin Is Required for Efficient S-Phase Progression and Stable Nuclear Architecture. International Journal of Molecular Sciences. 2021; 22(11):5483. https://doi.org/10.3390/ijms22115483
Chicago/Turabian StyleBustamante-Jaramillo, Luisa F., Celia Ramos, and Cristina Martín-Castellanos. 2021. "The Meiosis-Specific Crs1 Cyclin Is Required for Efficient S-Phase Progression and Stable Nuclear Architecture" International Journal of Molecular Sciences 22, no. 11: 5483. https://doi.org/10.3390/ijms22115483
APA StyleBustamante-Jaramillo, L. F., Ramos, C., & Martín-Castellanos, C. (2021). The Meiosis-Specific Crs1 Cyclin Is Required for Efficient S-Phase Progression and Stable Nuclear Architecture. International Journal of Molecular Sciences, 22(11), 5483. https://doi.org/10.3390/ijms22115483