
Update in Biomolecular and Genetic Bases of Bicuspid Aortopathy

 ,
,
Abstract
:1. Introduction
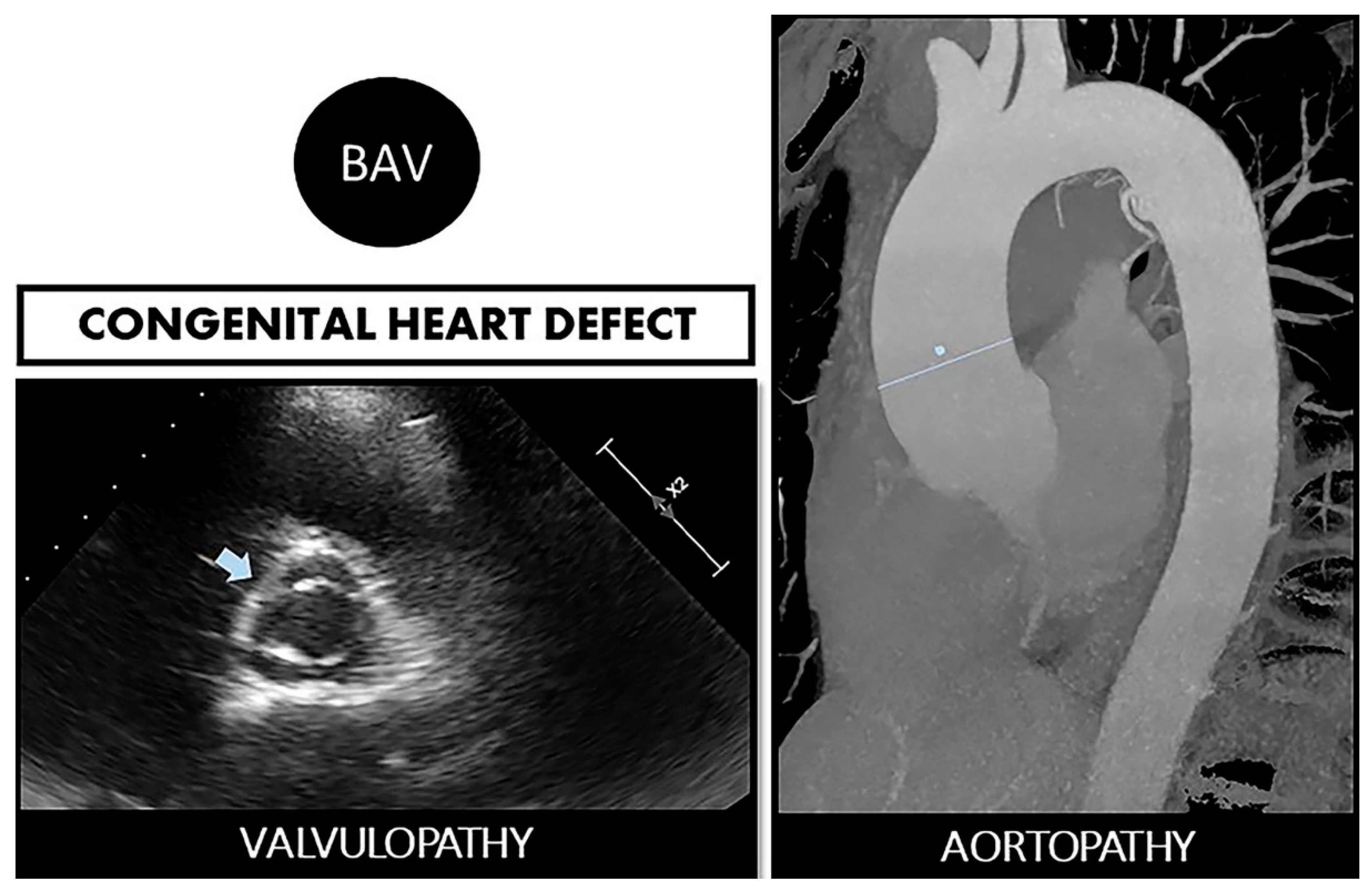
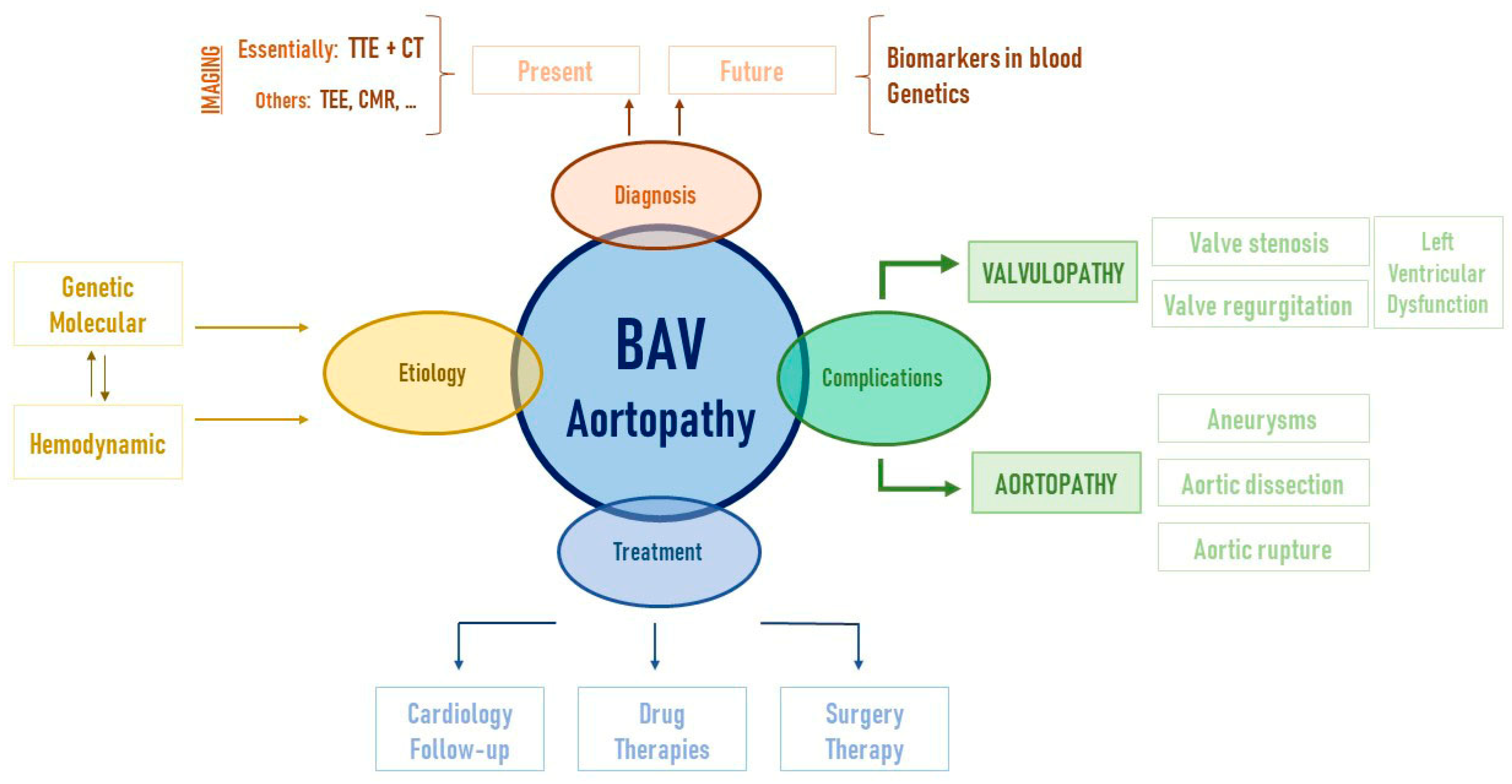
2. BAV and Aortopathy: A Two-Way Road
3. Genetics Insights into Bicuspid Aortopathy
4. Pieces and Findings in Epigenetics
5. Pieces and Findings Related to MicroRNAs and Other Regulatory RNAs
6. Molecular and Cellular Biology of the Aortic Valve and Associated Aortopathy
7. Is There a Biomarker for Bicuspid Aortopathy?
8. Future Directions
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pedersen, M.W.; Groth, K.A.; Mortensen, K.H.; Brodersen, J.; Gravholt, C.H.; Andersen, N.H. Clinical and pathophysiological aspects of bicuspid aortic valve disease. Cardiol. Young 2019, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Siu, S.C.; Silversides, C.K. Bicuspid aortic valve disease. J. Am. Coll. Cardiol. 2010, 55, 2789–2800. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Siu, S.C. Aortic dilatation in patients with bicuspid aortic valve. N. Engl. J. Med. 2014, 370, 1920–1929. [Google Scholar] [CrossRef] [Green Version]
- Michelena, H.I.; Khanna, A.D.; Mahoney, D.; Margaryan, E.; Topilsky, Y.; Suri, R.M.; Eidem, B.; Edwards, W.D.; Sundt, T.M.; Enriquez-Sarano, M. Incidence of Aortic Complications in Patients with Bicuspid Aortic Valves. JAMA 2011, 306, 1104. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, V.L.; Pancyzk, E.; Alpert, J.S. A new disease: Bicuspid aortic valve aortopathy syndrome. Am. J. Med. 2012, 125, 322–323. [Google Scholar] [CrossRef]
- Freeze, S.L.; Landis, B.J.; Ware, S.M.; Helm, B.M. Bicuspid Aortic Valve: A Review with Recommendations for Genetic Counseling. J. Genet. Couns. 2016, 25, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Borger, M.A.; Fedak, P.W.M.; Stephens, E.H.; Gleason, T.G.; Girdauskas, E.; Ikonomidis, J.S.; Khoynezhad, A.; Siu, S.C.; Verma, S.; Hope, M.D.; et al. The American Association for Thoracic Surgery consensus guidelines on bicuspid aortic valve-related aortopathy: Executive summary. J. Thorac. Cardiovasc. Surg. 2018, 156, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Michelena, H.I.; Desjardins, V.A.; Avierinos, J.-F.O.; Russo, A.; Nkomo, V.T.; Sundt, T.M.; Pellikka, P.A.; Tajik, A.J.; Enriquez-Sarano, M. Natural History of Asymptomatic Patients with Normally Functioning or Minimally Dysfunctional Bicuspid Aortic Valve in the Community. Circulation 2008, 117, 2776–2784. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, I.; Agapito, A.F.; de Sousa, L.; Oliveira, J.A.; Branco, L.M.; Galrinho, A.; Abreu, J.; Timoteo, A.T.; Rosa, S.A.; Ferreira, R.C. Bicuspid aortic valve outcomes. Cardiol. Young 2017, 27, 518–529. [Google Scholar] [CrossRef]
- Messner, B.; Bernhard, D. Bicuspid aortic valve-associated aortopathy: Where do we stand? J. Mol. Cell. Cardiol. 2019, 133, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzemos, N.; Therrien, J.; Yip, J.; Thanassoulis, G.; Tremblay, S.; Jamorski, M.T.; Webb, G.D.; Siu, S.C. Outcomes in adults with bicuspid aortic valves. JAMA 2008, 300, 1317–1325. [Google Scholar] [CrossRef] [Green Version]
- Della Corte, A.; Bancone, C.; Quarto, C.; Dialetto, G.; Covino, F.E.; Scardone, M.; Caianiello, G.; Cotrufo, M. Predictors of ascending aortic dilatation with bicuspid aortic valve: A wide spectrum of disease expression. Eur. J. Cardiothorac. Surg. 2007, 31, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.S.; Kloesel, B.; Norris, R.A.; Lindsay, M.; Milan, D.; Body, S.C. Embryonic Development of the Bicuspid Aortic Valve. J. Cardiovasc. Dev. Dis. 2015, 2, 248–272. [Google Scholar] [CrossRef]
- Hope, M.D.; Hope, T.A.; Crook, S.E.; Ordovas, K.G.; Urbania, T.H.; Alley, M.T.; Higgins, C.B. 4D flow CMR in assessment of valve-related ascending aortic disease. JACC Cardiovasc. Imaging 2011, 4, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Mahadevia, R.; Barker, A.J.; Schnell, S.; Entezari, P.; Kansal, P.; Fedak, P.W.; Malaisrie, S.C.; McCarthy, P.; Collins, J.; Carr, J.; et al. Bicuspid aortic cusp fusion morphology alters aortic three-dimensional outflow patterns, wall shear stress, and expression of aortopathy. Circulation 2014, 129, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Ruzmetov, M.; Shah, J.J.; Fortuna, R.S.; Welke, K.F. The Association Between Aortic Valve Leaflet Morphology and Patterns of Aortic Dilation in Patients with Bicuspid Aortic Valves. Ann. Thorac. Surg. 2015, 99, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Jackson, V.; Petrini, J.; Caidahl, K.; Eriksson, M.J.; Liska, J.; Eriksson, P.; Franco-Cereceda, A. Bicuspid aortic valve leaflet morphology in relation to aortic root morphology: A study of 300 patients undergoing open-heart surgery. Eur. J. Cardiothorac. Surg. 2011, 40, e118–e124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habchi, K.M.; Ashikhmina, E.; Vieira, V.M.; Shahram, J.T.; Isselbacher, E.M.; Sundt, T.M., 3rd; Shekar, P.; Muehlschlegel, J.D.; Bicuspid Aortic Valve, C.; Body, S.C. Association between bicuspid aortic valve morphotype and regional dilatation of the aortic root and trunk. Int. J. Cardiovasc. Imaging 2017, 33, 341–349. [Google Scholar] [CrossRef]
- Bissell, M.M.; Hess, A.T.; Biasiolli, L.; Glaze, S.J.; Loudon, M.; Pitcher, A.; Davis, A.; Prendergast, B.; Markl, M.; Barker, A.J.; et al. Aortic dilation in bicuspid aortic valve disease: Flow pattern is a major contributor and differs with valve fusion type. Circ. Cardiovasc. Imaging 2013, 6, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Guzzardi, D.G.; Barker, A.J.; van Ooij, P.; Malaisrie, S.C.; Puthumana, J.J.; Belke, D.D.; Mewhort, H.E.; Svystonyuk, D.A.; Kang, S.; Verma, S.; et al. Valve-Related Hemodynamics Mediate Human Bicuspid Aortopathy: Insights from Wall Shear Stress Mapping. J. Am. Coll. Cardiol. 2015, 66, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Fedak, P.W.; de Sa, M.P.; Verma, S.; Nili, N.; Kazemian, P.; Butany, J.; Strauss, B.H.; Weisel, R.D.; David, T.E. Vascular matrix remodeling in patients with bicuspid aortic valve malformations: Implications for aortic dilatation. J. Thorac. Cardiovasc. Surg. 2003, 126, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Li, J.; Wang, Y.; Wu, B.; Barker, A.J.; Markl, M.; Wang, C.; Wang, X.; Shu, X. Aortic shear stress in patients with bicuspid aortic valve with stenosis and insufficiency. J. Thorac. Cardiovasc. Surg. 2017, 153, 1263–1272.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollache, E.; Guzzardi, D.G.; Sattari, S.; Olsen, K.E.; Di Martino, E.S.; Malaisrie, S.C.; van Ooij, P.; Collins, J.; Carr, J.; McCarthy, P.M.; et al. Aortic valve-mediated wall shear stress is heterogeneous and predicts regional aortic elastic fiber thinning in bicuspid aortic valve-associated aortopathy. J. Thorac. Cardiovasc. Surg. 2018, 156, 2112–2120.e2. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, A.; Bancone, C.; Dialetto, G.; Covino, F.E.; Manduca, S.; Montibello, M.V.; De Feo, M.; Buonocore, M.; Nappi, G. The ascending aorta with bicuspid aortic valve: A phenotypic classification with potential prognostic significance. Eur. J. Cardiothorac. Surg. 2014, 46, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galian-Gay, L.; Carro Hevia, A.; Teixido-Tura, G.; Rodriguez Palomares, J.; Gutierrez-Moreno, L.; Maldonado, G.; Gonzalez-Alujas, M.T.; Sao-Aviles, A.; Gallego, P.; Calvo-Iglesias, F.; et al. Familial clustering of bicuspid aortic valve and its relationship with aortic dilation in first-degree relatives. Heart 2019, 105, 603–608. [Google Scholar] [CrossRef] [Green Version]
- Girdauskas, E.; Rouman, M.; Disha, K.; Espinoza, A.; Misfeld, M.; Borger, M.A.; Kuntze, T. Aortic Dissection After Previous Aortic Valve Replacement for Bicuspid Aortic Valve Disease. J. Am. Coll. Cardiol. 2015, 66, 1409–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticchi, E.; De Cario, R.; Magi, A.; Giglio, S.; Provenzano, A.; Nistri, S.; Pepe, G.; Giusti, B. Bicuspid Aortic Valve: Role of Multiple Gene Variants in Influencing the Clinical Phenotype. Biomed. Res. Int. 2018, 2018, 8386123. [Google Scholar] [CrossRef]
- Nistri, S.; Porciani, M.C.; Attanasio, M.; Abbate, R.; Gensini, G.F.; Pepe, G. Association of Marfan syndrome and bicuspid aortic valve: Frequency and outcome. Int. J. Cardiol. 2012, 155, 324–325. [Google Scholar] [CrossRef]
- Bondy, C.; Bakalov, V.K.; Cheng, C.; Olivieri, L.; Rosing, D.R.; Arai, A.E. Bicuspid aortic valve and aortic coarctation are linked to deletion of the X chromosome short arm in Turner syndrome. J. Med. Genet. 2013, 50, 662–665. [Google Scholar] [CrossRef] [Green Version]
- Emanuel, R.; Withers, R.; O’Brien, K.; Ross, P.; Feizi, O. Congenitally bicuspid aortic valves. Clinicogenetic study of 41 families. Br. Heart J. 1978, 40, 1402–1407. [Google Scholar] [CrossRef]
- Huntington, K.; Hunter, A.G.; Chan, K.L. A prospective study to assess the frequency of familial clustering of congenital bicuspid aortic valve. J. Am. Coll. Cardiol. 1997, 30, 1809–1812. [Google Scholar] [CrossRef]
- Cripe, L.; Andelfinger, G.; Martin, L.J.; Shooner, K.; Benson, D.W. Bicuspid aortic valve is heritable. J. Am. Coll. Cardiol. 2004, 44, 138–143. [Google Scholar] [CrossRef]
- Luna-Zurita, L.; Prados, B.; Grego-Bessa, J.; Luxan, G.; del Monte, G.; Benguria, A.; Adams, R.H.; Perez-Pomares, J.M.; de la Pompa, J.L. Integration of a Notch-dependent mesenchymal gene program and Bmp2-driven cell invasiveness regulates murine cardiac valve formation. J. Clin. Investig. 2010, 120, 3493–3507. [Google Scholar] [CrossRef] [Green Version]
- Garg, V.; Muth, A.N.; Ransom, J.F.; Schluterman, M.K.; Barnes, R.; King, I.N.; Grossfeld, P.D.; Srivastava, D. Mutations in NOTCH1 cause aortic valve disease. Nature 2005, 437, 270–274. [Google Scholar] [CrossRef]
- Li, R.G.; Xu, Y.J.; Wang, J.; Liu, X.Y.; Yuan, F.; Huang, R.T.; Xue, S.; Li, L.; Liu, H.; Li, Y.J.; et al. GATA4 Loss-of-Function Mutation and the Congenitally Bicuspid Aortic Valve. Am. J. Cardiol. 2018, 121, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Montes, C.; Martin, M.; Martinez-Arias, L.; Coto, E.; Naves-Diaz, M.; Moris, C.; Cannata-Andia, J.B.; Rodriguez, I. Variants in cardiac GATA genes associated with bicuspid aortic valve. Eur. J. Clin. Investig. 2018, 48, e13027. [Google Scholar] [CrossRef]
- Laforest, B.; Nemer, M. GATA5 interacts with GATA4 and GATA6 in outflow tract development. Dev. Biol. 2011, 358, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.J.; Di, R.M.; Qiao, Q.; Li, X.M.; Huang, R.T.; Xue, S.; Liu, X.Y.; Wang, J.; Yang, Y.Q. GATA6 loss-of-function mutation contributes to congenital bicuspid aortic valve. Gene 2018, 663, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Gharibeh, L.; Komati, H.; Bosse, Y.; Boodhwani, M.; Heydarpour, M.; Fortier, M.; Hassanzadeh, R.; Ngu, J.; Mathieu, P.; Body, S.; et al. GATA6 Regulates Aortic Valve Remodeling, and Its Haploinsufficiency Leads to Right-Left Type Bicuspid Aortic Valve. Circulation 2018, 138, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.F.; Xu, Y.J.; Zhao, C.M.; Wang, X.H.; Qiu, X.B.; Liu, X.; Wu, S.H.; Yang, Y.Q. A novel TBX5 mutation predisposes to familial cardiac septal defects and atrial fibrillation as well as bicuspid aortic valve. Genet. Mol. Biol. 2020, 43, e20200142. [Google Scholar] [CrossRef]
- Gillis, E.; Kumar, A.A.; Luyckx, I.; Preuss, C.; Cannaerts, E.; van de Beek, G.; Wieschendorf, B.; Alaerts, M.; Bolar, N.; Vandeweyer, G.; et al. Candidate Gene Resequencing in a Large Bicuspid Aortic Valve-Associated Thoracic Aortic Aneurysm Cohort: SMAD6 as an Important Contributor. Front. Physiol. 2017, 8, 400. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, I.; MacCarrick, G.; Kempers, M.; Meester, J.; Geryl, C.; Rombouts, O.; Peeters, N.; Claes, C.; Boeckx, N.; Sakalihasan, N.; et al. Confirmation of the role of pathogenic SMAD6 variants in bicuspid aortic valve-related aortopathy. Eur. J. Hum. Genet. 2019, 27, 1044–1053. [Google Scholar] [CrossRef] [PubMed]
- Attanasio, M.; Lapini, I.; Evangelisti, L.; Lucarini, L.; Giusti, B.; Porciani, M.; Fattori, R.; Anichini, C.; Abbate, R.; Gensini, G.; et al. FBN1 mutation screening of patients with Marfan syndrome and related disorders: Detection of 46 novel FBN1 mutations. Clin. Genet. 2008, 74, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.A.; Aziz, H.; Woods, C.E.; Seman-Senderos, M.A.; Sparks, E.; Preuss, C.; Wunnemann, F.; Bedja, D.; Moats, C.R.; McClymont, S.A.; et al. ROBO4 variants predispose individuals to bicuspid aortic valve and thoracic aortic aneurysm. Nat. Genet. 2019, 51, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.C.; Pannu, H.; Tran-Fadulu, V.; Papke, C.L.; Yu, R.K.; Avidan, N.; Bourgeois, S.; Estrera, A.L.; Safi, H.J.; Sparks, E.; et al. Mutations in smooth muscle alpha-actin (ACTA2) lead to thoracic aortic aneurysms and dissections. Nat. Genet. 2007, 39, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Longobardo, L.; Carerj, M.L.; Pizzino, G.; Bitto, A.; Piccione, M.C.; Zucco, M.; Oreto, L.; Todaro, M.C.; Calabro, M.P.; Squadrito, F.; et al. Impairment of elastic properties of the aorta in bicuspid aortic valve: Relationship between biomolecular and aortic strain patterns. Eur. Heart J. Cardiovasc. Imaging 2018, 19, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Debiec, R.; Hamby, S.E.; Jones, P.D.; Coolman, S.; Asiani, M.; Kharodia, S.; Skinner, G.J.; Samani, N.J.; Webb, T.R.; Bolger, A. Novel loss of function mutation in NOTCH1 in a family with bicuspid aortic valve, ventricular septal defect, thoracic aortic aneurysm, and aortic valve stenosis. Mol. Genet. Genomic. Med. 2020, 8, e1437. [Google Scholar] [CrossRef]
- Kerstjens-Frederikse, W.S.; van de Laar, I.M.; Vos, Y.J.; Verhagen, J.M.; Berger, R.M.; Lichtenbelt, K.D.; Klein Wassink-Ruiter, J.S.; van der Zwaag, P.A.; du Marchie Sarvaas, G.J.; Bergman, K.A.; et al. Cardiovascular malformations caused by NOTCH1 mutations do not keep left: Data on 428 probands with left-sided CHD and their families. Genet. Med. 2016, 18, 914–923. [Google Scholar] [CrossRef]
- Whitcomb, J.; Gharibeh, L.; Nemer, M. From embryogenesis to adulthood: Critical role for GATA factors in heart development and function. IUBMB Life 2020, 72, 53–67. [Google Scholar] [CrossRef]
- Perrucci, G.L.; Rurali, E.; Gowran, A.; Pini, A.; Antona, C.; Chiesa, R.; Pompilio, G.; Nigro, P. Vascular smooth muscle cells in Marfan syndrome aneurysm: The broken bricks in the aortic wall. Cell. Mol. Life Sci. 2017, 74, 267–277. [Google Scholar] [CrossRef]
- Pepe, G.; Nistri, S.; Giusti, B.; Sticchi, E.; Attanasio, M.; Porciani, C.; Abbate, R.; Bonow, R.O.; Yacoub, M.; Gensini, G.F. Identification of fibrillin 1 gene mutations in patients with bicuspid aortic valve (BAV) without Marfan syndrome. BMC Med. Genet. 2014, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- Tortora, G.; Wischmeijer, A.; Berretta, P.; Alfonsi, J.; Di Marco, L.; Barbieri, A.; Marconi, C.; Isidori, F.; Rossi, C.; Leone, O.; et al. Search for genetic factors in bicuspid aortic valve disease: ACTA2 mutations do not play a major role. Interact. Cardiovasc. Thorac. Surg. 2017, 25, 813–817. [Google Scholar] [CrossRef] [Green Version]
- Colpaert, R.M.W.; Calore, M. Epigenetics and microRNAs in cardiovascular diseases. Genomics 2021, 113, 540–551. [Google Scholar] [CrossRef]
- Gu, W.; Cheng, Y.; Wang, S.; Sun, T.; Li, Z. PHD Finger Protein 19 Promotes Cardiac Hypertrophy via Epigenetically Regulating SIRT2. Cardiovasc. Toxicol. 2021. [Google Scholar] [CrossRef]
- Li, Q.; Li, Z.; Fan, Z.; Yang, Y.; Lu, C. Involvement of non-coding RNAs in the pathogenesis of myocardial ischemia/reperfusion injury (Review). Int. J. Mol. Med. 2021, 47, 42. [Google Scholar] [CrossRef]
- Andreassi, M.G.; Della Corte, A. Genetics of bicuspid aortic valve aortopathy. Curr. Opin. Cardiol. 2016, 31, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Girdauskas, E.; Borger, M.A.; Secknus, M.A.; Girdauskas, G.; Kuntze, T. Is aortopathy in bicuspid aortic valve disease a congenital defect or a result of abnormal hemodynamics? A critical reappraisal of a one-sided argument. Eur. J. Cardiothorac. Surg. 2011, 39, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Wei, S.Y.; Chiu, J.J. Mechanical regulation of epigenetics in vascular biology and pathobiology. J. Cell. Mol. Med. 2013, 17, 437–448. [Google Scholar] [CrossRef]
- Pan, S.; Lai, H.; Shen, Y.; Breeze, C.; Beck, S.; Hong, T.; Wang, C.; Teschendorff, A.E. DNA methylome analysis reveals distinct epigenetic patterns of ascending aortic dissection and bicuspid aortic valve. Cardiovasc. Res. 2017, 113, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.A.; Gregory, S.G.; Krupp, D.; Feng, S.; Dorogi, A.; Haynes, C.; Grass, E.; Lin, S.S.; Hauser, E.R.; Kraus, W.E.; et al. Epigenetic profiling identifies novel genes for ascending aortic aneurysm formation with bicuspid aortic valves. Heart Surg. Forum 2015, 18, E134–E139. [Google Scholar] [CrossRef] [Green Version]
- Bjorck, H.M.; Du, L.; Pulignani, S.; Paloschi, V.; Lundstromer, K.; Kostina, A.S.; Osterholm, C.; Malashicheva, A.; Kostareva, A.; Evangelista, A.; et al. Altered DNA methylation indicates an oscillatory flow mediated epithelial-to-mesenchymal transition signature in ascending aorta of patients with bicuspid aortic valve. Sci. Rep. 2018, 8, 2777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, D.; Coyet, A.; Ollivier, V.; Jeunemaitre, X.; Jondeau, G.; Michel, J.B.; Vranckx, R. Epigenetic control of vascular smooth muscle cells in Marfan and non-Marfan thoracic aortic aneurysms. Cardiovasc. Res. 2011, 89, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Forte, A.; Galderisi, U.; Cipollaro, M.; De Feo, M.; Della Corte, A. Epigenetic regulation of TGF-beta1 signalling in dilative aortopathy of the thoracic ascending aorta. Clin. Sci. (Lond) 2016, 130, 1389–1405. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Balistreri, C.R.; Forte, M.; Greco, E.; Paneni, F.; Cavarretta, E.; Frati, G.; Sciarretta, S. An overview of the molecular mechanisms underlying development and progression of bicuspid aortic valve disease. J. Mol. Cell. Cardiol. 2019, 132, 146–153. [Google Scholar] [CrossRef]
- Vecoli, C.; Borghini, A.; Turchi, S.; Mercuri, A.; Andreassi, M.G. Genetic polymorphisms of miRNA machinery genes in bicuspid aortic valve and associated aortopathy. Per. Med. 2021, 18, 21–29. [Google Scholar] [CrossRef]
- Yanagawa, B.; Lovren, F.; Pan, Y.; Garg, V.; Quan, A.; Tang, G.; Singh, K.K.; Shukla, P.C.; Kalra, N.P.; Peterson, M.D.; et al. miRNA-141 is a novel regulator of BMP-2-mediated calcification in aortic stenosis. J. Thorac. Cardiovasc. Surg. 2012, 144, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Pulignani, S.; Borghini, A.; Andreassi, M.G. microRNAs in bicuspid aortic valve associated aortopathy: Recent advances and future perspectives. J. Cardiol. 2019, 74, 297–303. [Google Scholar] [CrossRef]
- Nigam, V.; Sievers, H.H.; Jensen, B.C.; Sier, H.A.; Simpson, P.C.; Srivastava, D.; Mohamed, S.A. Altered microRNAs in bicuspid aortic valve: A comparison between stenotic and insufficient valves. J. Heart Valve. Dis. 2010, 19, 459–465. [Google Scholar] [PubMed]
- Naito, S.; Petersen, J.; Sequeira-Gross, T.; Neumann, N.; Duque Escobar, J.; Zeller, T.; Reichenspurner, H.; Girdauskas, E. Bicuspid aortopathy—Molecular involvement of microRNAs and MMP-TIMP. Biomarkers 2020, 25, 711–718. [Google Scholar] [CrossRef]
- Martinez-Micaelo, N.; Beltran-Debon, R.; Baiges, I.; Faiges, M.; Alegret, J.M. Specific circulating microRNA signature of bicuspid aortic valve disease. J. Transl. Med. 2017, 15, 76. [Google Scholar] [CrossRef] [Green Version]
- Pulignani, S.; Borghini, A.; Foffa, I.; Vecoli, C.; Ait-Ali, L.; Andreassi, M.G. Functional characterization and circulating expression profile of dysregulated microRNAs in BAV-associated aortopathy. Heart Vessel. 2020, 35, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Maegdefessel, L. Non-coding RNA Contribution to Thoracic and Abdominal Aortic Aneurysm Disease Development and Progression. Front. Physiol. 2017, 8, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrion, K.; Dyo, J.; Patel, V.; Sasik, R.; Mohamed, S.A.; Hardiman, G.; Nigam, V. The long non-coding HOTAIR is modulated by cyclic stretch and WNT/beta-CATENIN in human aortic valve cells and is a novel repressor of calcification genes. PLoS ONE 2014, 9, e96577. [Google Scholar] [CrossRef] [Green Version]
- Leon-Mimila, P.; Wang, J.; Huertas-Vazquez, A. Relevance of Multi-Omics Studies in Cardiovascular Diseases. Front. Cardiovasc. Med. 2019, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Malashicheva, A.; Kostina, A.; Kostareva, A.; Irtyuga, O.; Gordeev, M.; Uspensky, V. Notch signaling in the pathogenesis of thoracic aortic aneurysms: A bridge between embryonic and adult states. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165631. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Crapanzano, F.; Schirone, L.; Allegra, A.; Pisano, C.; Ruvolo, G.; Forte, M.; Greco, E.; Cavarretta, E.; Marullo, A.G.M.; et al. Deregulation of Notch1 pathway and circulating endothelial progenitor cell (EPC) number in patients with bicuspid aortic valve with and without ascending aorta aneurysm. Sci. Rep. 2018, 8, 13834. [Google Scholar] [CrossRef] [PubMed]
- Sciacca, S.; Pilato, M.; Mazzoccoli, G.; Pazienza, V.; Vinciguerra, M. Anti-correlation between longevity gene SirT1 and Notch signaling in ascending aorta biopsies from patients with bicuspid aortic valve disease. Heart Vessel. 2013, 28, 268–275. [Google Scholar] [CrossRef]
- Kostina, A.; Semenova, D.; Kostina, D.; Uspensky, V.; Kostareva, A.; Malashicheva, A. Human aortic endothelial cells have osteogenic Notch-dependent properties in co-culture with aortic smooth muscle cells. Biochem. Biophys. Res. Commun. 2019, 514, 462–468. [Google Scholar] [CrossRef]
- Antequera-Gonzalez, B.; Martinez-Micaelo, N.; Alegret, J.M. Bicuspid Aortic Valve and Endothelial Dysfunction: Current Evidence and Potential Therapeutic Targets. Front. Physiol. 2020, 11, 1015. [Google Scholar] [CrossRef]
- Peterson, J.C.; Wisse, L.J.; Wirokromo, V.; van Herwaarden, T.; Smits, A.M.; Gittenberger-de Groot, A.C.; Goumans, M.T.H.; VanMunsteren, J.C.; Jongbloed, M.R.M.; DeRuiter, M.C. Disturbed nitric oxide signalling gives rise to congenital bicuspid aortic valve and aortopathy. Dis. Model Mech. 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Gauer, S.; Balint, B.; Kollmann, C.; Federspiel, J.M.; Henn, D.; Bandner-Risch, D.; Schmied, W.; Schafers, H.J. Dysregulation of Endothelial Nitric Oxide Synthase Does Not Depend on Hemodynamic Alterations in Bicuspid Aortic Valve Aortopathy. J. Am. Heart Assoc. 2020, 9, e016471. [Google Scholar] [CrossRef] [PubMed]
- Hirata, Y.; Aoki, H.; Shojima, T.; Takagi, K.; Takaseya, T.; Akasu, K.; Tobinaga, S.; Fukumoto, Y.; Tanaka, H. Activation of the AKT Pathway in the Ascending Aorta with Bicuspid Aortic Valve. Circ. J. 2018, 82, 2485–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skeffington, K.L.; Bond, A.R.; Bigotti, M.G.; AbdulGhani, S.; Iacobazzi, D.; Kang, S.L.; Heesom, K.J.; Wilson, M.C.; Stoica, S.; Martin, R.; et al. Changes in inflammation and oxidative stress signalling pathways in coarcted aorta triggered by bicuspid aortic valve and growth in young children. Exp. Ther. Med. 2020, 20, 48. [Google Scholar] [CrossRef]
- Forte, A.; Yin, X.; Fava, M.; Bancone, C.; Cipollaro, M.; De Feo, M.; Mayr, M.; Jahangiri, M.; Della Corte, A. Locally different proteome in aortas from patients with stenotic tricuspid and bicuspid aortic valvesdagger. Eur. J. Cardiothorac. Surg. 2019, 56, 458–469. [Google Scholar] [CrossRef]
- Surendran, A.; Edel, A.; Chandran, M.; Bogaert, P.; Hassan-Tash, P.; Kumar Asokan, A.; Hiebert, B.; Solati, Z.; Sandhawalia, S.; Raabe, M.; et al. Metabolomic Signature of Human Aortic Valve Stenosis. JACC Basic Transl. Sci. 2020, 5, 1163–1177. [Google Scholar] [CrossRef]
- MacGrogan, D.; Martinez-Poveda, B.; Desvignes, J.P.; Fernandez-Friera, L.; Gomez, M.J.; Gil Vilarino, E.; Callejas Alejano, S.; Garcia-Pavia, P.; Solis, J.; Lucena, J.; et al. Identification of a peripheral blood gene signature predicting aortic valve calcification. Physiol. Genom. 2020, 52, 563–574. [Google Scholar] [CrossRef]
- Balint, B.; Yin, H.; Nong, Z.; Arpino, J.M.; O’Neil, C.; Rogers, S.R.; Randhawa, V.K.; Fox, S.A.; Chevalier, J.; Lee, J.J.; et al. Seno-destructive smooth muscle cells in the ascending aorta of patients with bicuspid aortic valve disease. EBioMedicine 2019, 43, 54–66. [Google Scholar] [CrossRef] [Green Version]
- Crawford, E.E.; McCarthy, P.M.; Malaisrie, S.C.; Puthumana, J.J.; Robinson, J.D.; Markl, M.; Liu, M.; Andrei, A.C.; Guzzardi, D.G.; Kruse, J.; et al. Applications of a Specialty Bicuspid Aortic Valve Program: Clinical Continuity and Translational Collaboration. J. Clin. Med. 2020, 9, 1354. [Google Scholar] [CrossRef]
- Bons, L.R.; Geenen, L.W.; van den Hoven, A.T.; Dik, W.A.; van den Bosch, A.E.; Duijnhouwer, A.L.; Siebelink, H.J.; Budde, R.P.J.; Boersma, E.; Wessels, M.W.; et al. Blood biomarkers in patients with bicuspid aortic valve disease. J. Cardiol. 2020, 76, 287–294. [Google Scholar] [CrossRef]
- Lo Presti, F.; Guzzardi, D.G.; Bancone, C.; Fedak, P.W.M.; Della Corte, A. The science of BAV aortopathy. Prog. Cardiovasc. Dis. 2020, 63, 465–474. [Google Scholar] [CrossRef]
- Della Corte, A.; Michelena, H.I.; Citarella, A.; Votta, E.; Piatti, F.; Lo Presti, F.; Ashurov, R.; Cipollaro, M.; Forte, A. Risk Stratification in Bicuspid Aortic Valve Aortopathy: Emerging Evidence and Future Perspectives. Curr. Probl. Cardiol. 2021, 46, 100428. [Google Scholar] [CrossRef]
- Ikonomidis, J.S.; Ruddy, J.M.; Benton, S.M., Jr.; Arroyo, J.; Brinsa, T.A.; Stroud, R.E.; Zeeshan, A.; Bavaria, J.E.; Gorman, J.H., 3rd; Gorman, R.C.; et al. Aortic dilatation with bicuspid aortic valves: Cusp fusion correlates to matrix metalloproteinases and inhibitors. Ann. Thorac. Surg. 2012, 93, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, B.; Dong, L.; Wang, C.; Wang, X.; Shu, X. Circulating matrix metalloproteinase patterns in association with aortic dilatation in bicuspid aortic valve patients with isolated severe aortic stenosis. Heart Vessel. 2016, 31, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Drapisz, S.; Goralczyk, T.; Jamka-Miszalski, T.; Olszowska, M.; Undas, A. Nonstenotic bicuspid aortic valve is associated with elevated plasma asymmetric dimethylarginine. J. Cardiovasc. Med. (Hagerstown) 2013, 14, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Gavriliuk, N.D.; Druzhkova, T.A.; Irtyuga, O.B.; Zhloba, A.A.; Subbotina, T.F.; Uspenskiy, V.E.; Alexeyeva, N.P.; Moiseeva, O.M. Asymmetric Dimethylarginine in Patients with Ascending Aortic Aneurysms. Aorta (Stamford) 2016, 4, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Branchetti, E.; Bavaria, J.E.; Grau, J.B.; Shaw, R.E.; Poggio, P.; Lai, E.K.; Desai, N.D.; Gorman, J.H.; Gorman, R.C.; Ferrari, G. Circulating soluble receptor for advanced glycation end product identifies patients with bicuspid aortic valve and associated aortopathies. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2349–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, A.; Bancone, C.; Cobellis, G.; Buonocore, M.; Santarpino, G.; Fischlein, T.J.M.; Cipollaro, M.; De Feo, M.; Della Corte, A. A Possible Early Biomarker for Bicuspid Aortopathy: Circulating Transforming Growth Factor beta-1 to Soluble Endoglin Ratio. Circ. Res. 2017, 120, 1800–1811. [Google Scholar] [CrossRef]
- Martinez-Micaelo, N.; Ligero, C.; Antequera-Gonzalez, B.; Junza, A.; Yanes, O.; Alegret, J.M. Plasma Metabolomic Profiling Associates Bicuspid Aortic Valve Disease and Ascending Aortic Dilation with a Decrease in Antioxidant Capacity. J. Clin. Med. 2020, 9, 2215. [Google Scholar] [CrossRef]
- Xiong, T.Y.; Liu, C.; Liao, Y.B.; Zheng, W.; Li, Y.J.; Li, X.; Ou, Y.; Wang, Z.J.; Wang, X.; Li, C.M.; et al. Differences in metabolic profiles between bicuspid and tricuspid aortic stenosis in the setting of transcatheter aortic valve replacement. BMC Cardiovasc. Disord. 2020, 20, 229. [Google Scholar] [CrossRef]
- Simsek, E.C.; Yakar Tuluce, S.; Tuluce, K.; Emren, S.V.; Cuhadar, S.; Nazli, C. The relationship between serum apelin levels and aortic dilatation in bicuspid aortic valve patients. Congenit. Heart Dis. 2019, 14, 256–263. [Google Scholar] [CrossRef]
- Hillebrand, M.; Millot, N.; Sheikhzadeh, S.; Rybczynski, M.; Gerth, S.; Kolbel, T.; Keyser, B.; Kutsche, K.; Robinson, P.N.; Berger, J.; et al. Total serum transforming growth factor-beta1 is elevated in the entire spectrum of genetic aortic syndromes. Clin. Cardiol. 2014, 37, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Agnese, V.; Coronnello, C.; Raffa, G.M.; Bellavia, D.; Conaldi, P.G.; Pilato, M.; Pasta, S. On the prospect of serum exosomal miRNA profiling and protein biomarkers for the diagnosis of ascending aortic dilatation in patients with bicuspid and tricuspid aortic valve. Int. J. Cardiol. 2018, 273, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Martinez, C.; Lamas, O.; Carrasco-Chinchilla, F.; Robledo-Carmona, J.; Porras, C.; Sanchez-Espin, G.; Navarro, M.J.; Fernandez, B. Increased blood levels of transforming growth factor beta in patients with aortic dilatation. Interact. Cardiovasc. Thorac. Surg. 2017, 25, 571–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, D.; Monteagudo, J.; Suarez-Farinas, M.; Bander, J.; Varshney, R.; Gonzalez, J.; Coller, B.S.; Ahamed, J. New methodologies to accurately assess circulating active transforming growth factor-beta1 levels: Implications for evaluating heart failure and the impact of left ventricular assist devices. Transl. Res. 2018, 192, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Leeper, N.J.; Tedesco, M.M.; Kojima, Y.; Schultz, G.M.; Kundu, R.K.; Ashley, E.A.; Tsao, P.S.; Dalman, R.L.; Quertermous, T. Apelin prevents aortic aneurysm formation by inhibiting macrophage inflammation. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1329–H1335. [Google Scholar] [CrossRef] [Green Version]
- Fatehi Hassanabad, A.; Garcia, J.; Verma, S.; White, J.A.; Fedak, P.W.M. Utilizing wall shear stress as a clinical biomarker for bicuspid valve-associated aortopathy. Curr. Opin. Cardiol. 2019, 34, 124–131. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | OMIM | Pathway/Role | Mutation Consequence | Reference |
|---|---|---|---|---|
| NOTCH1 | 190198 | Development of valve and endocardial cushions | Defect on the aortic valve and de-repression of calcium deposition | [33,34] |
| GATA4 | 600576 | Cardiovascular embryogenesis | Enhanced susceptibility to BAV | [35,36] |
| GATA5 | 611496 | Cardiovascular embryogenesis | BAV development | [36,37] |
| GATA6 | 601656 | Cardiovascular embryogenesis | Enhanced susceptibility to familial BAV | [36,38,39] |
| TBX5 | 601620 | Cardiac development | Atrial fibrillation. Segregates with BAV | [40] |
| SMAD6 | 602931 | TGF-β signaling pathway | Related with BAV + aortic aneurysm | [41,42] |
| FBN1 | 134797 | Integrity of the aortic media | Related with isolated BAV and Marfan syndrome with aortic root dilation | [43] |
| ROBO4 | 607528 | Transmembrane receptor | Segregating with ascending aortic aneurysm in BAV | [44] |
| ACTA2 | 102620 | Smooth muscle α-actin | Related with familial ascending aortic aneurysm in BAV | [45] |
| ELN | 130160 | Arterial development | Correlated with ascending aorta diameter in BAV | [46] |
| miRNAs | Regulation | Target | BAV-Aortopathy | Reference |
|---|---|---|---|---|
| 34 out of 1583 | 8↑ 26↓ | − | BAV | [67] |
| miR-141 | ↓ | BMP2-dependent calcification-related genes | BAV + aortic stenosis | [67] |
| miR-424 | ↓ | SMAD7 | BAV + aortic aneurysm | [68] |
| miR-16, miR-26a, miR-27a miR-30b, miR-130, miR-195, miR-497 | ↓ | − | BAV | [69] |
| miR-130, miR-195, miR-497 | ↓ | Pro calcifying genes ALPL and BMP2 | BAV + aortic stenosis/aortic regurgitation | [69] |
| miR-133a miR-143 | ↓ | TIMP2 | BAV + aortic aneurysm | [70] |
| miR-122 miR-486 | ↓ | TGF-β pathway-related genes | BAV | [71] |
| miR-130a | ↑ | TGF-β pathway-related genes | BAV | [71] |
| miR-718 | ↓ | Blood vessel remodeling and focal adhesion pathways-related genes | BAV + aortic aneurysm | [71] |
| Biomarker | Levels | BAV Associated-Aortopathy | Reference |
|---|---|---|---|
| MMPs/TIMPs | ↓TIMPs | BAV + aortic aneurysm | [93] |
| ↑MMPs | Levels of MMP-2 correlate with aortic dimensions in BAV + aortic aneurysm | [94] | |
| ADMA | ↑ | Non-stenotic BAV + aortic aneurysm | [95] |
| ↑ | Aortic aneurysm independently of valve morphology | [96] | |
| sRAGE | − | Levels in plasma indicative of BAV status | [97] |
| NT-ProBNP hsTnT hsCRP | ↑ ↑ ↑ | BAV + aortic stenosis and regurgitation BAV + aortic regurgitation BAV without any correlation with aortic diameter. | [90] |
| TGF-β | ↑ | Higher serum TGF-β/sENG ratio in BAV patients undergoing valve replacement due to aortic stenosis | [98] |
| ↓ | BAV independently of aortic dilation | [90] | |
| Alpha-tocopherol CRP, EMPs | ↑CRP, EMPs ↓Alpha-tocopherol | BAV | [99] |
| Pro/Arg pathways | ↓ | BAV + aortic stenosis | [100] |
| Apelin | ↓ | BAV + aortic aneurysm compared to BAV without ascending aorta dilation and control subjects | [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Junco-Vicente, A.; del Río-García, Á.; Martín, M.; Rodríguez, I. Update in Biomolecular and Genetic Bases of Bicuspid Aortopathy. Int. J. Mol. Sci. 2021, 22, 5694. https://doi.org/10.3390/ijms22115694
Junco-Vicente A, del Río-García Á, Martín M, Rodríguez I. Update in Biomolecular and Genetic Bases of Bicuspid Aortopathy. International Journal of Molecular Sciences. 2021; 22(11):5694. https://doi.org/10.3390/ijms22115694
Chicago/Turabian StyleJunco-Vicente, Alejandro, Álvaro del Río-García, María Martín, and Isabel Rodríguez. 2021. "Update in Biomolecular and Genetic Bases of Bicuspid Aortopathy" International Journal of Molecular Sciences 22, no. 11: 5694. https://doi.org/10.3390/ijms22115694
APA StyleJunco-Vicente, A., del Río-García, Á., Martín, M., & Rodríguez, I. (2021). Update in Biomolecular and Genetic Bases of Bicuspid Aortopathy. International Journal of Molecular Sciences, 22(11), 5694. https://doi.org/10.3390/ijms22115694