
Coaggregation of Asthma and Type 1 Diabetes in Children: A Narrative Review



Abstract
:1. Introduction
2. Divergent Theories Underlying Asthma and Type 1 Diabetes Topic
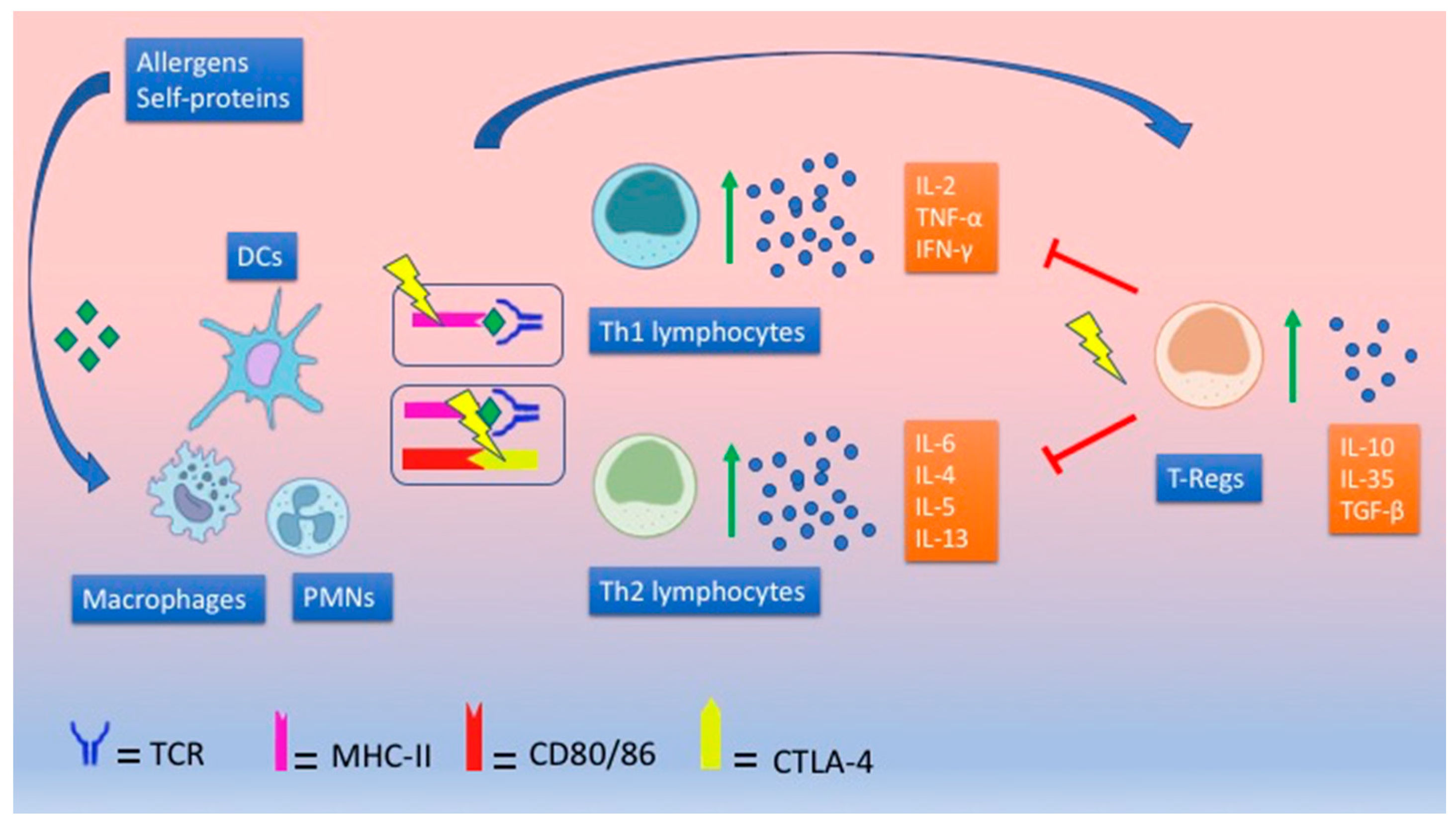
2.1. Th1/Th2 Paradigm: Are Asthma and T1DM Mutually Exclusive?
- Th1 cells defend against infections and tumors but are also involved in the development of autoimmune diseases (T1DM), producing cytokines such as interferon (INF)-γ, Tumor Necrosis Factor (TNF)-α, and interleukin (IL)-2 [11].
- Th2 cells protect against parasites but also promote the development of IgE-mediated atopic diseases (asthma), producing cytokines such as IL-4, IL-5 IL-6, IL-9, IL-10, and IL-13 [11].
2.2. The “Hygiene Hypothesis”: Could Asthma and T1DM Have a Common Environmental Background?
2.2.1. Infections
2.2.2. Gut Microbiome
2.2.3. Parasitic Infections
2.3. Genetic Protective or Risk Factors for Asthma and Type 1 Diabetes
3. Studies Reporting no Association between Asthma and Type 1 Diabetes
4. Inflammatory Pattern of the Patients with Asthma and Type 1 Diabetes
5. Influence of Asthma on T1DM Control and Vice Versa
6. Temporal Relationship between Asthma and Type 1 Diabetes: Asthma to Type 1 Diabetes or Type 1 Diabetes to Asthma?
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CTLA-4 | Cytotoxic T- lymphocytes antigen-4 |
| DCs | Dendritic cells |
| FhHDM | Fasciola hepatica helminth defense molecule |
| GADA | Glutamic acid decarboxylase antibodies |
| GIMAP | GTPases of the immunity associated protein |
| GPR43 | G-protein coupled receptor 43 |
| GWAS | Genome wide association study |
| HbA1c | Glycated hemoglobin |
| HLA | Human leukocyte antigen |
| HbA1c | Glycated hemoglobin |
| IAA | Autoantibodies to insulin |
| IA-2A | Tyrosine phosphatase related insulinoma associated 2 molecule |
| ICA | Islet cell antibodies |
| Ig | Immunoglobulin |
| IL | Interleukin |
| INF | Interferon |
| ISAAC | International study of asthma and allergy in childhood |
| LPS | Lipopolysaccharide |
| MHC | Major histocompatibility complex |
| NOD | Non-obese diabetic |
| PAMPs | Pathogen associated molecular patterns |
| PBMCs | Peripheral blood mononuclear cells |
| SCFAs | Small chain fatty acids |
| SNP | Single nucleotide polymorphism |
| Th | T helper |
| TLR | Toll-like receptors |
| TNF-α | Tumor necrosis factor alpha |
| Tregs | Regulatory T lymphocytes |
| T1DM | Type 1 diabetes mellitus |
References
- Castagnoli, R.; Licari, A.; Manti, S.; Chiappini, E.; Marseglia, G.L. Type-2 inflammatory mediators as targets for precision medicine in children. Pediatr. Allergy Immunol. 2020, 31, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Asthma. Global Strategegy for Asthma Management and Prevention. 2020. Available online: https://ginasthma.org/wp-content/uploads/2020/04/GINA-2020-full-report_-final-_wms.pdf (accessed on 7 February 2021).
- Lai, C.K.W.; Beasley, R.; Crane, J.; Foliaki, S.; Shah, J.; Weiland, S.; Aït-Khaled, N.; Anderson, H.R.; Asher, M.I.; Beasley, R.; et al. Global variation in the prevalence and severity of asthma symptoms: Phase Three of the International Study of Asthma and Allergies in Childhood (ISAAC). Thorax 2009, 64, 476–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dharmage, S.C.; Perret, J.L.; Custovic, A. Epidemiology of Asthma in Children and Adults. Front. Pediatr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Akinbami, L.J.; Simon, A.E.; Rossen, L.M. Changing Trends in Asthma Prevalence among Children. Pediatrics 2016, 137, e20152354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mameli, C.; Ghezzi, M.; Mari, A.; Cammi, G.; Macedoni, M.; Redaelli, F.C.; Calcaterra, V.; Zuccotti, G.; D’Auria, E. The Diabetic Lung: Insights into Pulmonary Changes in Children and Adolescents with Type 1 Diabetes. Metabolites 2021, 11, 69. [Google Scholar] [CrossRef]
- Op de Beeck, A.; Eizirik, D.L. Viral infections in type 1 diabetes mellitus—Why the β cells? Nat. Rev. Endocrinol. 2016, 12, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mobasseri, M.; Shirmohammadi, M.; Amiri, T.; Vahed, N.; Fard, H.H.; Ghojazadeh, M. Prevalence and incidence of type 1 diabetes in the world: A systematic review and meta-analysis. Heal. Promot. Perspect. 2020, 10, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Xie, Z.; Huang, G.; Zhou, Z. Incidence and trend of type 1 diabetes and the underlying environmental determinants. Diabetes. Metab. Res. Rev. 2019, 35, e3075. [Google Scholar] [CrossRef] [Green Version]
- Altobelli, E.; Petrocelli, R.; Verrotti, A.; Chiarelli, F.; Marziliano, C. Genetic and environmental factors affect the onset of type 1 diabetes mellitus. Pediatr. Diabetes 2016, 17, 559–566. [Google Scholar] [CrossRef]
- Mukherjee, M.; Nair, P. Autoimmune Responses in Severe Asthma. Allergy Asthma Immunol. Res. 2018, 10, 428. [Google Scholar] [CrossRef]
- Patterson, C.C.; Dahlquist, G.G.; Gyürüs, E.; Green, A.; Soltész, G.; Schober, E.; Waldhoer, T.; Weets, I.; Rooman, R.; Gorus, F.; et al. Incidence trends for childhood type 1 diabetes in Europe during 1989-2003 and predicted new cases 2005-20: A multicentre prospective registration study. Lancet 2009, 373, 2027–2033. [Google Scholar] [CrossRef]
- Meerwaldt, R.; Odink, R.J.; Landaeta, R.; Aarts, F.; Brunekreef, B.; Gerritsen, J.; Van Aalderen, W.M.C.; Hoekstra, M.O. A lower prevalence of atopy symptoms in children with type 1 diabetes mellitus. Clin. Exp. Allergy 2002, 32, 254–255. [Google Scholar] [CrossRef]
- Mattila, P.S.; Tarkkanen, J.; Saxen, H.; Pitkäniemi, J.; Karvonen, M.; Tuomilehto, J. Predisposition to atopic symptoms to inhaled antigens may protect from childhood type 1 diabetes. Diabetes Care 2002, 25, 865–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taleb, N.; Bou Khalil, P.; Zantout, M.S.; Zalloua, P.; Azar, S.T. Prevalence of asthmatic symptoms in Lebanese patients with type 1 diabetes and their unaffected siblings compared to age-matched controls. Acta Diabetol. 2010, 47, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 Cells: Different Patterns of Lymphokine Secretion Lead to Different Functional Properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef] [PubMed]
- Tosca, M.A.; Silòvestri, M.; D’Annunzio, G.; Lorini, R.; Rossi, G.A.; Ciprandi, G. May T1 diabetes mellitus protect from asthma? Allergol. Immunopathol. 2013, 41, 288–291. [Google Scholar] [CrossRef]
- Cardwell, C.R.; Shields, M.D.; Carson, D.J.; Patterson, C.C. A meta-analysis of the association between childhood type 1 diabetes and atopic disease. Diabetes Care 2003, 26, 2568–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirghani, H.O.; Alhazmi, K.; Alghamdi, S.; Alraddadi, M. The Cross-Talk between Atopic Dermatitis and Diabetes Mellitus: A Meta-Analysis. Cureus 2021. [Google Scholar] [CrossRef] [PubMed]
- Pusch, E.; Renz, H.; Skevaki, C. Respiratory virus-induced heterologous immunity. Allergol. J. Int. 2018, 27, 79–96. [Google Scholar] [CrossRef]
- Fsadni, P.; Fsadni, C.; Fava, S.; Montefort, S. Correlation of worldwide incidence of type 1 diabetes (DiaMond) with prevalence of asthma and atopic eczema (ISAAC). Clin. Respir. J. 2012, 6, 18–25. [Google Scholar] [CrossRef]
- Klamt, S.; Vogel, M.; Kapellen, T.M.; Hiemisch, A.; Prenzel, F.; Zachariae, S.; Ceglarek, U.; Thiery, J.; Kiess, W. Association between IgE-mediated allergies and diabetes mellitus type 1 in children and adolescents. Pediatr. Diabetes 2015, 16, 493–503. [Google Scholar] [CrossRef]
- Villa-Nova, H.; Spinola-Castro, A.M.; Garcia, F.E.; Solé, D. Prevalence of allergic diseases and/or allergic sensitisation in children and adolescents with type 1 diabetes mellitus. Allergol. Immunopathol. 2015, 43, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.T.; Cheng, W.C.; Liao, W.C.; Lin, C.L.; Shen, T.C.; Chen, W.C.; Chen, C.H.; Kao, C.H. Type 1 diabetes and increased risk of subsequent asthma: A nationwide population-based cohort study. Medicine 2015, 94, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Black, M.H.; Anderson, A.; Bell, R.A.; Dabelea, D.; Pihoker, C.; Saydah, S.; Seid, M.; Standiford, D.A.; Waitzfelder, B.; Marcovina, S.M.; et al. Prevalence of asthma and its association with glycemic control among youth with diabetes. Pediatrics 2011, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hörtenhuber, T.; Kiess, W.; Fröhlich-Reiterer, E.; Raile, K.; Stachow, R.; Bollow, E.; Rami-Merhar, B.; Holl, R.W. Asthma in children and adolescents with type 1 diabetes in Germany and Austria: Frequency and metabolic control. Pediatr. Diabetes 2018, 19, 727–732. [Google Scholar] [CrossRef]
- Castro-Rodriguez, J.A.; Forno, E.; Rodriguez-Martinez, C.E.; Celedón, J.C. Risk and Protective Factors for Childhood Asthma: What Is the Evidence? J. Allergy Clin. Immunol. Pract. 2016, 4, 1111–1122. [Google Scholar] [CrossRef] [Green Version]
- Karvonen, M.; Maarit, V.-K.; Elena, M.; Ingrid, L.; Ronald, L.; Jaakko, T. Incidence of Childhood Type 1 Diabetes. Diabetes Care 2000, 23, 1516–1526. [Google Scholar] [CrossRef] [Green Version]
- Strachan, D.; Sibbald, B.; Weiland, S.; Aït-Khaled, N.; Anabwani, G.; Anderson, H.R.; Asher, M.I.; Beasley, R.; Björkstén, B.; Burr, M.; et al. Worldwide variations in prevalence of symptoms of allergic rhinoconjunctivitis in children: The International Study of Asthma and Allergies in Childhood (ISAAC). Pediatr. Allergy Immunol. 1997, 8, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, H.; Robertson, C.; Stewart, A.; Aït-Khaled, N.; Anabwani, G.; Anderson, R.; Asher, I.; Beasley, R.; Björkstén, B.; Burr, M.; et al. Worldwide variations in the prevalence of symptoms of atopic eczema in the international study of asthma and allergies in childhood. J. Allergy Clin. Immunol. 1999, 103, 125–138. [Google Scholar] [CrossRef]
- Worldwide variations in the prevalence of asthma symptoms: The International Study of Asthma and Allergies in Childhood (ISAAC). Eur. Respir. J. 1998, 12, 315–335. [CrossRef] [Green Version]
- Stene, L.C.; Nafstad, P. Relation between occurrence of type 1 diabetes and asthma. Lancet 2001, 357, 607–608. [Google Scholar] [CrossRef]
- EURODIAB ACE Study Group. Variation and trends in incidence of childhood diabetes in Europe. Lancet 2000, 355, 873–876. [Google Scholar] [CrossRef]
- Tang, C.; Gao, Y.; Wang, L.; Zhu, Y.; Pan, Q.; Zhang, R.; Xiong, Y. Role of regulatory T cells in Schistosoma-mediated protection against type 1 diabetes. Mol. Cell. Endocrinol. 2019, 491, 110434. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.T.; Ma, Y.; Sun, Y.; Bai, C.Q.; Ling, C.H.; Yuan, F.L. The Protective Effects of Helicobacter pylori Infection on Allergic Asthma. Int. Arch. Allergy Immunol. 2021, 182, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Garn, H.; Renz, H. Epidemiological and immunological evidence for the hygiene hypothesis. Immunobiology 2007, 212, 441–452. [Google Scholar] [CrossRef]
- Machiels, B.; Dourcy, M.; Xiao, X.; Javaux, J.; Mesnil, C.; Sabatel, C.; Desmecht, D.; Lallemand, F.; Martinive, P.; Hammad, H.; et al. A gammaherpesvirus provides protection against allergic asthma by inducing the replacement of resident alveolar macrophages with regulatory monocytes. Nat. Immunol. 2017, 18, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Granata, M.; Skarmoutsou, E.; Trovato, C.; Rossi, G.A.; Mazzarino, M.C.; D’Amico, F. Obesity, Type 1 Diabetes, and Psoriasis: An Autoimmune Triple Flip. Pathobiology 2017, 84, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D. Family size, infection and atopy: The first decade of the “hygiene hypothesis”. Thorax 2000, 55, S2–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfefferle, P.I.; Keber, C.U.; Cohen, R.M.; Garn, H. The Hygiene Hypothesis—Learning From but Not Living in the Past. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Cardwell, C.R.; Carson, D.J.; Yarnell, J.; Shields, M.D.; Patterson, C.C. Atopy, home environment and the risk of childhood-onset type 1 diabetes: A population-based case-control study. Pediatr. Diabetes 2008, 9, 191–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikkinen, S.M.M.; Pitkäniemi, J.M.; Kilpeläinen, M.L.; Koskenvuo, M.J. Does farm environment protect against type 1 diabetes mellitus? Diabetes Vasc. Dis. Res. 2013, 10, 375–377. [Google Scholar] [CrossRef] [Green Version]
- Vatanen, T.; Kostic, A.D.; D’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Haahtela, T.; Laatikainen, T.; Alenius, H.; Auvinen, P.; Fyhrquist, N.; Hanski, I.; von Hertzen, L.; Jousilahti, P.; Kosunen, T.U.; Markelova, O.; et al. Hunt for the origin of allergy—Comparing the Finnish and Russian Karelia. Clin. Exp. Allergy 2015, 45, 891–901. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Lynch, S.V. Microbiota in Allergy and Asthma and the Emerging Relationship with the Gut Microbiome. Cell Host Microbe 2015, 17, 592–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knip, M.; Honkanen, J. Modulation of Type 1 Diabetes Risk by the Intestinal Microbiome. Curr. Diab. Rep. 2017, 17, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.D.; Gevers, D.; Siljander, H.; Vatanen, T.; Hyötyläinen, T.; Hämäläinen, A.-M.; Peet, A.; Tillmann, V.; Pöhö, P.; Mattila, I.; et al. The Dynamics of the Human Infant Gut Microbiome in Development and in Progression toward Type 1 Diabetes. Cell Host Microbe 2015, 17, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.C.; Ownby, D.R. The infant gut bacterial microbiota and risk of pediatric asthma and allergic diseases. Transl. Res. 2017, 179, 60–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postler, T.S.; Ghosh, S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 26, 110–130. [Google Scholar] [CrossRef] [Green Version]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Kim, C. B cell-helping functions of gut microbial metabolites. Microb. Cell 2016, 3, 529–531. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Qie, Y.; Park, J.; Kim, C.H. Gut Microbial Metabolites Fuel Host Antibody Responses. Cell Host Microbe 2016, 20, 202–214. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Li, Y.; Fang, J.; Liu, G.; Yin, J.; Li, T.; Yin, Y. Gut Microbiota and Type 1 Diabetes. Int. J. Mol. Sci. 2018, 19, 995. [Google Scholar] [CrossRef] [Green Version]
- Van Crevel, R.; van de Vijver, S.; Moore, D.A.J. The global diabetes epidemic: What does it mean for infectious diseases in tropical countries? Lancet Diabetes Endocrinol. 2017, 5, 457–468. [Google Scholar] [CrossRef]
- Stiemsma, L.; Reynolds, L.; Turvey, S.; Finlay, B. The hygiene hypothesis: Current perspectives and future therapies. ImmunoTargets Ther. 2015, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Wang, L.; Tang, Y.; Sun, X. Parasite-Derived Proteins for the Treatment of Allergies and Autoimmune Diseases. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulendran, B.; Tang, H.; Manicassamy, S. Programming dendritic cells to induce TH2 and tolerogenic responses. Nat. Immunol. 2010, 11, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.; Kaisar, M.M.M.; Wiria, A.E.; Hamid, F.; Djuardi, Y.; Sartono, E.; Rosa, B.A.; Mitreva, M.; Supali, T.; Houwing-Duistermaat, J.J.; et al. The Effect of Gut Microbiome Composition on Human Immune Responses: An Exploration of Interference by Helminth Infections. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Berbudi, A.; Ajendra, J.; Wardani, A.P.F.; Hoerauf, A.; Hübner, M.P. Parasitic helminths and their beneficial impact on type 1 and type 2 diabetes. Diabetes. Metab. Res. Rev. 2016, 32, 238–250. [Google Scholar] [CrossRef]
- Wang, M.; Wu, L.; Weng, R.; Zheng, W.; Wu, Z.; Lv, Z. Therapeutic potential of helminths in autoimmune diseases: Helminth-derived immune-regulators and immune balance. Parasitol. Res. 2017, 116, 2065–2074. [Google Scholar] [CrossRef]
- Alvarado, R.; O’Brien, B.; Tanaka, A.; Dalton, J.P.; Donnelly, S. A parasitic helminth-derived peptide that targets the macrophage lysosome is a novel therapeutic option for autoimmune disease. Immunobiology 2015, 220, 262–269. [Google Scholar] [CrossRef]
- Eissa, M.M.; Mostafa, D.K.; Ghazy, A.A.; El Azzouni, M.Z.; Boulos, L.M.; Younis, L.K. Anti-Arthritic Activity of Schistosoma mansoni and Trichinella spiralis Derived-Antigens in Adjuvant Arthritis in Rats: Role of FOXP3+ Treg Cells. PLoS ONE 2016, 11, e0165916. [Google Scholar] [CrossRef]
- Bjørnvold, M.; Munthe-Kaas, M.C.; Egeland, T.; Joner, G.; Dahl-Jørgensen, K.; Njølstad, P.R.; Akselsen, H.E.; Gervin, K.; Carlsen, K.C.L.; Carlsen, K.H.; et al. A TLR2 polymorphism is associated with type 1 diabetes and allergic asthma. Genes Immun. 2009, 10, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Moffatt, M.F.; Gut, I.G.; Demenais, F.; Strachan, D.P.; Bouzigon, E.; Heath, S.; von Mutius, E.; Farrall, M.; Lathrop, M.; Cookson, W.O.C.M. A Large-Scale, Consortium-Based Genomewide Association Study of Asthma. N. Engl. J. Med. 2010, 363, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, J.C.; Clayton, D.; Concannon, P.; Akolkar, B.; Cooper, J.D.; Erlich, H.A.; Julier, C.; Morahan, G.; Nerup, J.; Nierras, C.; et al. Genome-wide association study and meta-analysis finds over 40 loci affect risk of type 1 diabetes HHS Public Access Author manuscript. Nat. Genet. 2009, 41, 703–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalloua, P.A.; Abchee, A.; Shbaklo, H.; Zreik, T.G.; Terwedow, H.; Halaby, G.; Azar, S.T. Patients with early onset of type 1 diabetes have significantly higher GG genotype at position 49 of the CTLA4 gene. Hum. Immunol. 2004, 65, 719–724. [Google Scholar] [CrossRef]
- Gao, J.; Lin, Y.; Qiu, C.; Liu, Y.; Ma, Y.; Liu, Y. Association between HLA-DQA1, -DQB1 gene polymorphisms and susceptibility to asthma in northern Chinese subjects. Chin. Med. J. 2003, 116, 1078–1082. [Google Scholar] [PubMed]
- Shadur, B.; Asherie, N.; Kfir-Erenfeld, S.; Dubnikov, T.; NaserEddin, A.; Schejter, Y.D.; Elpeleg, O.; Mor-Shaked, H.; Stepensky, P. A human case of GIMAP6 deficiency: A novel primary immune deficiency. Eur. J. Hum. Genet. 2021, 29, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, M.T.; Laine, A.-P.; Söderhäll, C.; Gruzieva, O.; Rautio, S.; Melén, E.; Pershagen, G.; Lähdesmäki, H.J.; Knip, M.; Ilonen, J.; et al. GIMAP GTPase Family Genes: Potential Modifiers in Autoimmune Diabetes, Asthma, and Allergy. J. Immunol. 2015, 194, 5885–5894. [Google Scholar] [CrossRef]
- Tosca, M.A.; Silvestri, M.; Olcese, R.; D’Annunzio, G.; Pistorio, A.; Lorini, R.; Ciprandi, G.; Rossi, G.A. Allergic sensitization and symptoms, body mass index, and respiratory function in children with type 1 diabetes mellitus. Ann. Allergy Asthma Immunol. 2012, 108, 128–129. [Google Scholar] [CrossRef]
- Tosca, M.A.; Villa, E.; Silvestri, M.; D’Annunzio, G.; Pistorio, A.; Aicardi, M.; Minicucci, L.; Lorini, R.; Rossi, G.A. Discrepancy between sensitization to inhaled allergens and respiratory symptoms in pediatric patients with type 1 diabetes mellitus. Pediatr. Allergy Immunol. 2009, 20, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Jasser-Nitsche, H.; Varga, E.-M.; Borkenstein, H.M.; Höntzsch, J.; Suppan, E.; Weinhandl, G.; Pieringer, L.; Avian, A.; Fröhlich-Reiterer, E. Type 1 diabetes in children and adolescents is not associated with a reduced prevalence of atopy and allergic diseases. Pediatr. Diabetes 2017, 18, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Schlaud, M.; Atzpodien, K.; Thierfelder, W. Allergische Erkrankungen: Ergebnisse aus dem Kinder- und Jugendgesundheitssurvey (KiGGS). Bundesgesundheitsblatt Gesundheitsforsch. Gesundh. 2007, 50, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Asher, M.; Montefort, S.; Bjorksten, B.; Lai, C.; Strachan, D.; Weiland, S.; Williams, H.; Phase, I.I. Worldwide time trends in the prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and eczema in childhood. Lancet 2006, 368, 733–743. [Google Scholar] [CrossRef]
- Proskocil, B.J.; Calco, G.N.; Nie, Z. Insulin acutely increases agonist-induced airway smooth muscle contraction in humans and rats. Am. J. Physiol. Cell Mol. Physiol. 2021, 320, L545–L556. [Google Scholar] [CrossRef] [PubMed]
- Kolahian, S.; Leiss, V.; Nürnberg, B. Diabetic lung disease: Fact or fiction? Rev. Endocr. Metab. Disord. 2019, 20, 303–319. [Google Scholar] [CrossRef]
- Sansone, F.; Attanasi, M.; Di Pillo, S.; Chiarelli, F. Asthma and Obesity in Children. Biomedicines 2020, 8, 231. [Google Scholar] [CrossRef]
- Nie, Z.; Jacoby, D.B.; Fryer, A.D. Hyperinsulinemia potentiates airway responsiveness to parasympathetic nerve stimulation in obese rats. Am. J. Respir. Cell Mol. Biol. 2014, 51, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Rachmiel, M.; Bloch, O.; Shaul, A.A.; Ben-Yehudah, G.; Bistritzer, Z.; Weintrob, N.; Ofan, R.; Rapoport, M.J. Young patients with both type 1 diabetes mellitus and asthma have a unique IL-12 and IL-18 secretory pattern. Pediatr. Diabetes 2011, 12, 596–603. [Google Scholar] [CrossRef]
- Kainonen, E.; Rautava, S.; Korkeamäki, M.; Isolauri, E. Unique cytokine secretion profile in children with both type I diabetes and asthma distinct from that of solely diabetic or asthmatic children. Cytokine 2006, 34, 198–205. [Google Scholar] [CrossRef]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 649. [Google Scholar] [CrossRef] [Green Version]
- Behzadi, P.; Behzadi, E.; Ranjbar, R. IL-12 Family Cytokines: General Characteristics, Pathogenic Microorganisms, Receptors, and Signalling Pathways. Acta Microbiol. Immunol. Hung. 2016, 63, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.W.; Peters, L.D.; Fuhrman, C.A.; Seay, H.R.; Posgai, A.L.; Stimpson, S.E.; Brusko, M.A.; Perry, D.J.; Yeh, W.-I.; Newby, B.N.; et al. Innate inflammation drives NK cell activation to impair Treg activity. J. Autoimmun. 2020, 108, 102417. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Saraiva, M.; Vieira, P.; Vieira, P.; Vieira, P.; O’Garra, A.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coomes, S.M.; Kannan, Y.; Pelly, V.S.; Entwistle, L.J.; Guidi, R.; Perez-Lloret, J.; Nikolov, N.; Müller, W.; Wilson, M.S. CD4+ Th2 cells are directly regulated by IL-10 during allergic airway inflammation. Mucosal Immunol. 2017, 10, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Ahmadizar, F.; Souverein, P.C.; Arets, H.G.M.; de Boer, A.; Maitland-van der Zee, A.H. Asthma related medication use and exacerbations in children and adolescents with type 1 diabetes. Pediatr. Pulmonol. 2016, 51, 1113–1121. [Google Scholar] [CrossRef]
- Smew, A.I.; Lundholm, C.; Sävendahl, L.; Lichtenstein, P.; Almqvist, C. Familial Coaggregation of Asthma and Type 1 Diabetes in Children. JAMA Netw. Open 2020, 3, e200834. [Google Scholar] [CrossRef]
- Metsälä, J.; Lundqvist, A.; Virta, L.J.; Kaila, M.; Gissler, M.; Virtanen, S.M.; Nevalainen, J. The association between asthma and type 1 diabetes: A paediatric case-cohort study in Finland, years 1981–2009. Int. J. Epidemiol. 2018, 47, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Krischer, J.P.; Cuthbertson, D.; Couluris, M.; Knip, M.; Virtanen, S.M. Association of diabetes-related autoantibodies with the incidence of asthma, eczema and allergic rhinitis in the TRIGR randomised clinical trial. Diabetologia 2020, 63, 1796–1807. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, J.; Vaarala, O.; Ludvigsson, J. Asthma and allergic symptoms and type 1 diabetes-related autoantibodies in 2.5-yr-old children. Pediatr. Diabetes 2011, 12, 604–610. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Protein | Function | T1DM-Associated | Asthma-Associated |
|---|---|---|---|---|
| FLG | Profilaggrin | Precursor of filaggrin, necessary for epidermal structure | No | Yes |
| SEPS1 | Selenoprotein S | Translocation of misfolded proteins from the endoplasmic reticulum to the cytosol | No | Possible |
| IL18 | Interleukin 18 | Proinflammatory cytokine of the IL-1 family | No | Possible |
| IL12RB1 | IL-12 receptor β1 | Subunit of the IL-12 receptor | No | No |
| IL12RB2 | IL-12 receptor β2 | Subunit of the IL-12 receptor | No | No |
| TLR2 | Toll Like Receptor 2 | Recognition of PAMPs and activation of the innate immune response | Yes | Yes |
| TLR4 | Toll Like Receptor 4 | Recognition of PAMPs and activation of the innate immune response | No | No |
| CD14 | Cluster of Differentiation 14 | Recognition of LPS and PAMPs by macrophages | No | No |
| GSDMB | Gasdermin B | Regulation of apoptosis | Yes | Yes |
| HLA-DRB1 | Human Leukocyte Antigen DR β1 | Subunit of HLA class II receptor | Yes | Yes |
| HLA-DQB1 | Human Leukocyte Antigen DQ β1 | Subunit of HLA class II receptor | Yes | Yes |
| CTLA4 | Cytotoxic T-lymphocyte-associated protein 4 | Downregulation of immune responses, constitutively expressed by Tregs | Possible | Possible |
| GIMAP | GTPase of the immunity-associated protein | Regulators of immune cells homeostasis | Yes | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sgrazzutti, L.; Sansone, F.; Attanasi, M.; Di Pillo, S.; Chiarelli, F. Coaggregation of Asthma and Type 1 Diabetes in Children: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 5757. https://doi.org/10.3390/ijms22115757
Sgrazzutti L, Sansone F, Attanasi M, Di Pillo S, Chiarelli F. Coaggregation of Asthma and Type 1 Diabetes in Children: A Narrative Review. International Journal of Molecular Sciences. 2021; 22(11):5757. https://doi.org/10.3390/ijms22115757
Chicago/Turabian StyleSgrazzutti, Laura, Francesco Sansone, Marina Attanasi, Sabrina Di Pillo, and Francesco Chiarelli. 2021. "Coaggregation of Asthma and Type 1 Diabetes in Children: A Narrative Review" International Journal of Molecular Sciences 22, no. 11: 5757. https://doi.org/10.3390/ijms22115757
APA StyleSgrazzutti, L., Sansone, F., Attanasi, M., Di Pillo, S., & Chiarelli, F. (2021). Coaggregation of Asthma and Type 1 Diabetes in Children: A Narrative Review. International Journal of Molecular Sciences, 22(11), 5757. https://doi.org/10.3390/ijms22115757