
Role Played by Receptors for Advanced Glycosylation End Products in Corneal Endothelial Cells after HSV-1 Infection

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
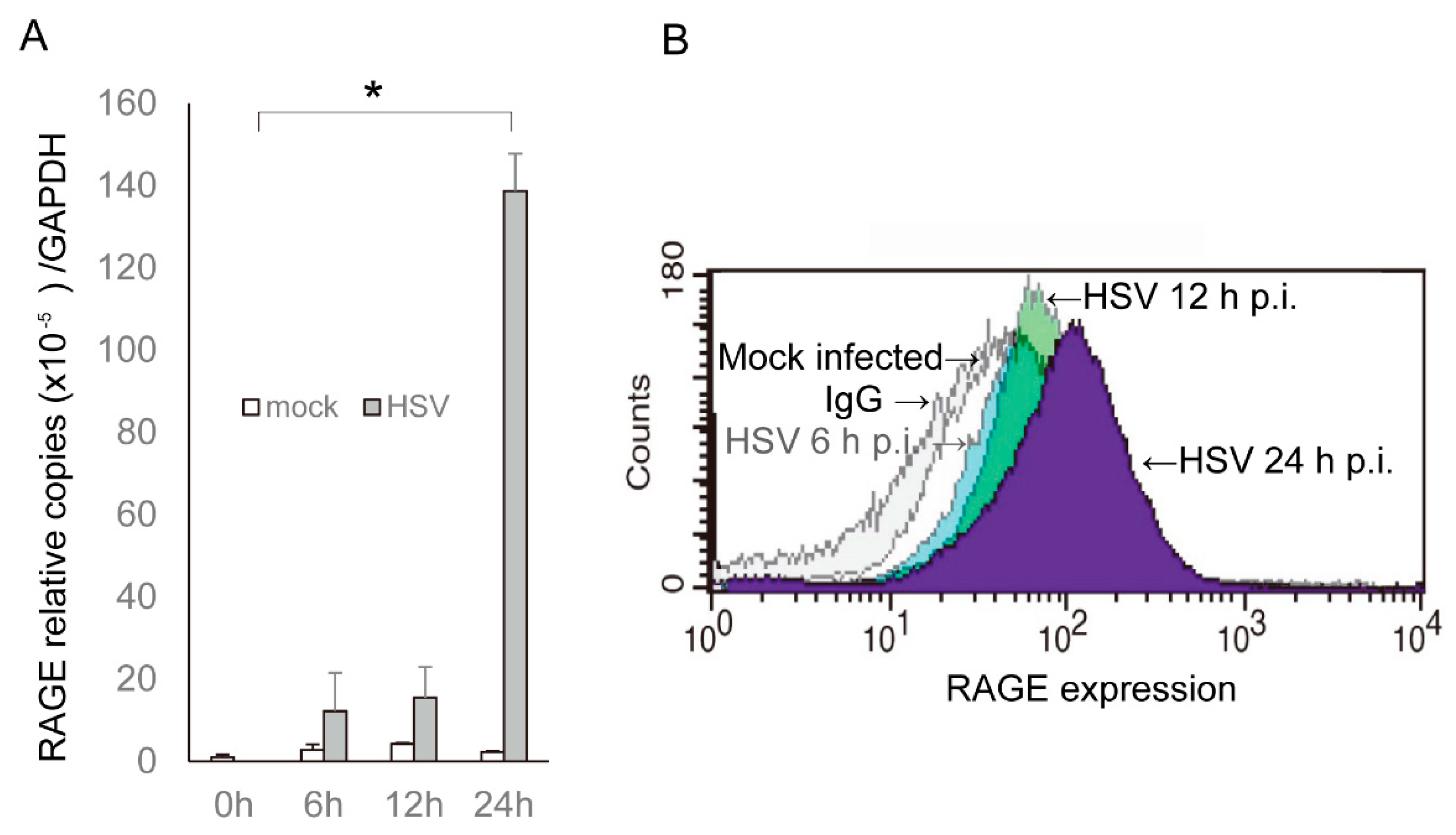
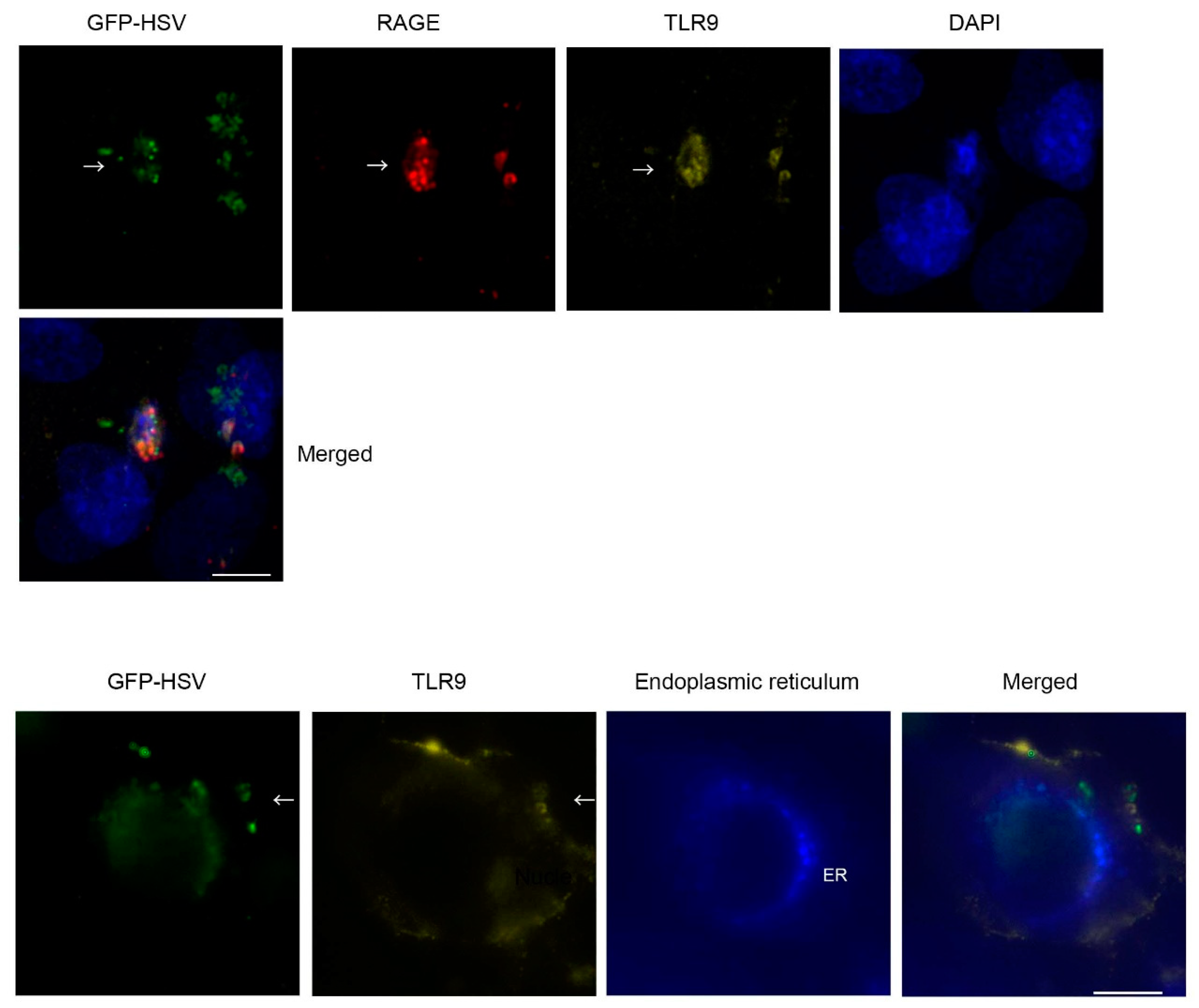
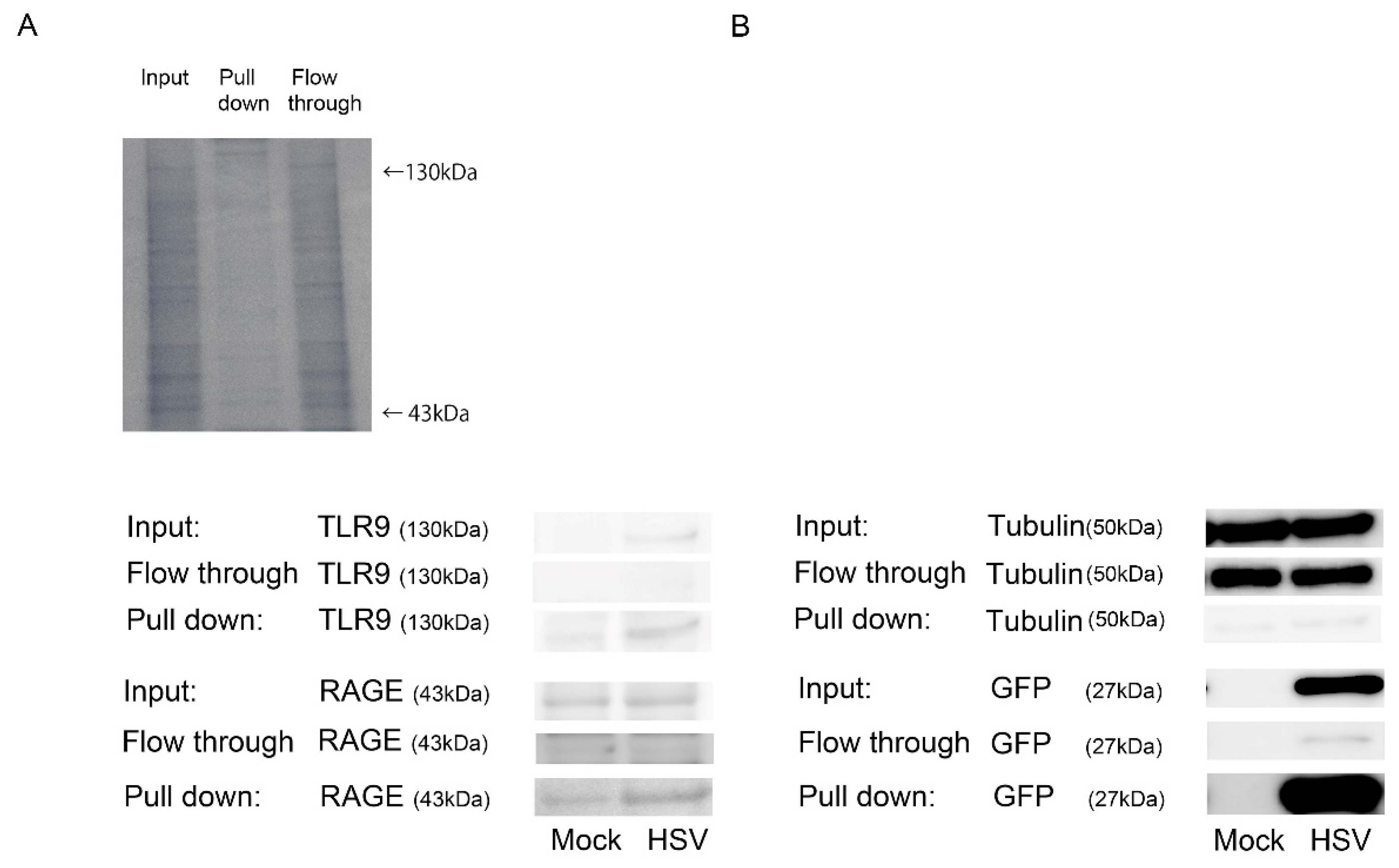
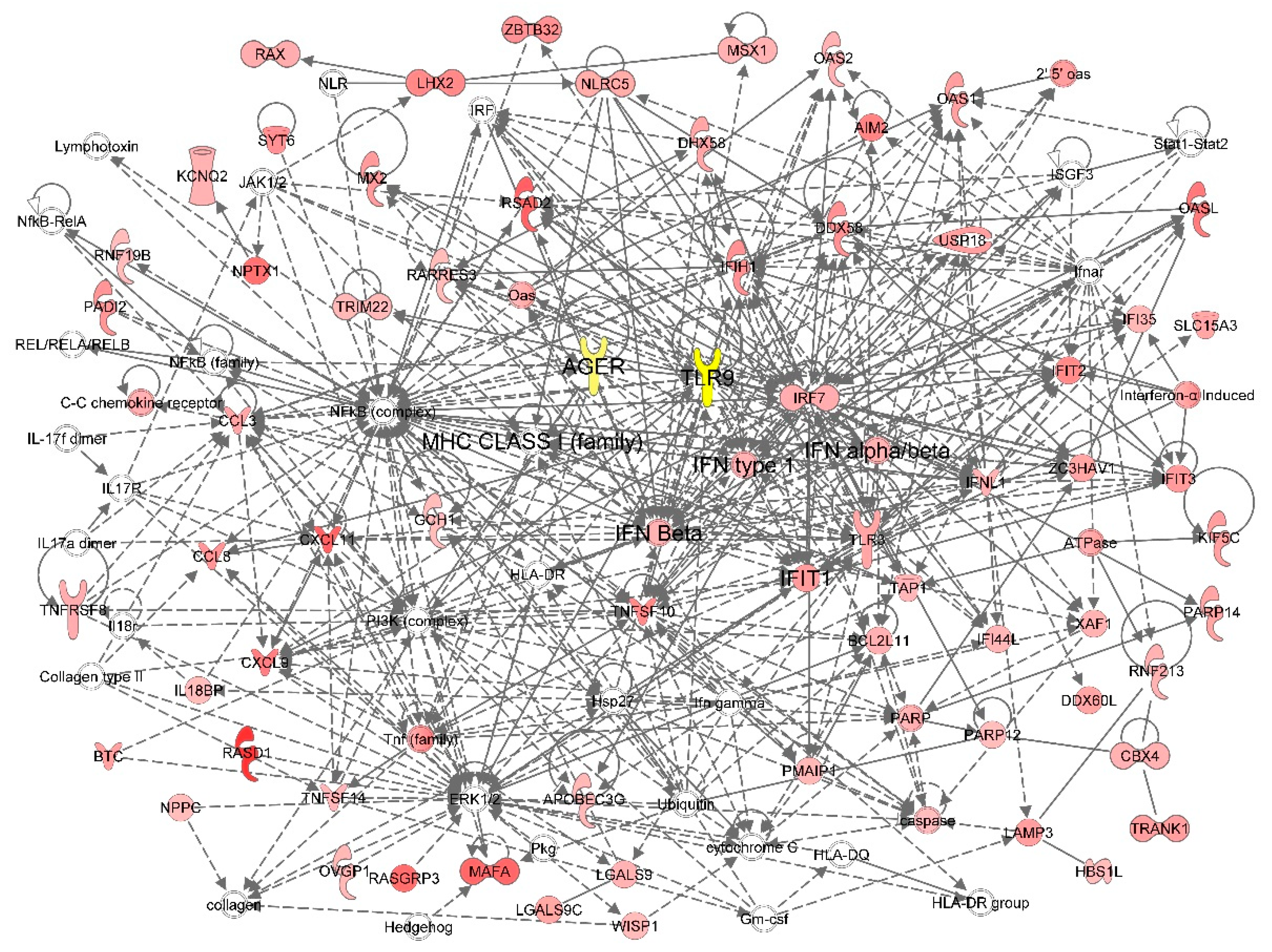
2. Results
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Virus
4.3. ELISA
4.4. Real-Time RT-PCR
4.5. Fluorescence-Activated Cell Sorting (FACS) Analyses
4.6. Immunohistochemistry
4.7. GFP Pull-Down Assay
4.8. Transfection of RAGE Expression Plasmid and Blockade of RAGE
4.9. Network Analysis of Corneal Endothelial Cell Transcriptome after HSV-1 Infection
4.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HSV-1 | herpes simplex virus type 1 |
| PAMP | pathogen-associated molecular patterns |
| RAGE | receptor for advanced glycosylation end products |
| AGE | advanced glycation end products |
| HCEn | human corneal endothelial cell |
| HMGB1 | high mobility group box 1 |
| TLR9 | toll-like receptor 9 |
| SAA | serum amyloid-A |
| PI | post-infection |
References
- Flaxman, S.R.; Bourne, R.R.A.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990–2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef] [Green Version]
- Ung, L.; Bispo, P.J.M.; Shanbhag, S.S.; Gilmore, M.S.; Chodosh, J. The persistent dilemma of microbial keratitis: Global burden, diagnosis, and antimicrobial resistance. Surv. Ophthalmol. 2019, 64, 255–271. [Google Scholar] [CrossRef]
- Garg, P.; Krishna, P.V.; Stratis, A.K.; Gopinathan, U. The value of corneal transplantation in reducing blindness. Eye 2005, 19, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.Y.; Snyder, P.J.; Wu, W.C.; Zhang, M.; Echeverria, A.; Alber, J. Pathophysiologic relationship between Alzheimer’s disease, cerebrovascular disease, and cardiovascular risk: A review and synthesis. Alzheimers Dement. 2017, 7, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Howes, K.A.; Liu, Y.; Dunaief, J.L.; Milam, A.; Frederick, J.M.; Marks, A.; Baehr, W. Receptor for advanced glycation end products and age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3713–3720. [Google Scholar] [CrossRef]
- Ramasamy, R.; Yan, S.F.; Schmidt, A.M. RAGE: Therapeutic target and biomarker of the inflammatory response--the evidence mounts. J. Leukoc. Biol. 2009, 86, 505–512. [Google Scholar] [CrossRef]
- Kierdorf, K.; Fritz, G. RAGE regulation and signaling in inflammation and beyond. J. Leukoc. Biol. 2013, 94, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Handa, J.T.; Green, W.R.; Stark, W.J.; Weinberg, R.S.; Jun, A.S. Advanced glycation end products and receptors in Fuchs’ dystrophy corneas undergoing Descemet’s stripping with endothelial keratoplasty. Ophthalmology 2007, 114, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandori, M.; Miyazaki, D.; Yakura, K.; Komatsu, N.; Touge, C.; Ishikura, R.; Inoue, Y. Relationship between the number of cytomegalovirus in anterior chamber and severity of anterior segment inflammation. Jpn J. Ophthalmol. 2013, 57, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, N.; Inatomi, T.; Suzuki, T.; Shiraishi, A.; Ohashi, Y.; Kandori, M.; Miyazaki, D.; Inoue, Y.; Soma, T.; Nishida, K.; et al. Clinical features and management of cytomegalovirus corneal endotheliitis:analysis of 106 cases from the Japan corneal endotheliitis study. Br. J. Ophthalmol. 2014, 99, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, D.; Shimizu, D.; Shimizu, Y.; Inoue, Y.; Inoue, T.; Higaki, S.; Ueta, M.; Sugita, S. Diagnostic efficacy of real-time PCR for ocular cytomegalovirus infections. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 2413–2420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Schillinger, J.A.; Sternberg, M.R.; Johnson, R.E.; Lee, F.K.; Nahmias, A.J.; Markowitz, L.E. Seroprevalence and coinfection with herpes simplex virus type 1 and type 2 in the United States, 1988–1994. J. Infect. Dis. 2002, 185, 1019–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, H.E.; Azcuy, A.M.; Varnell, E.D.; Sloop, G.D.; Thompson, H.W.; Hill, J.M. HSV-1 DNA in tears and saliva of normal adults. Investig. Ophthalmol. Vis. Sci. 2005, 46, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, D.; Haruki, T.; Takeda, S.; Sasaki, S.; Yakura, K.; Terasaka, Y.; Komatsu, N.; Yamagami, S.; Touge, H.; Touge, C.; et al. Herpes simplex virus type 1-induced transcriptional networks of corneal endothelial cells indicate antigen presentation function. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4282–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Miyazaki, D.; Sasaki, S.; Yamamoto, Y.; Terasaka, Y.; Yakura, K.; Yamagami, S.; Ebihara, N.; Inoue, Y. Roles played by toll-like receptor-9 in corneal endothelial cells after herpes simplex virus type 1 infection. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6729–6736. [Google Scholar] [CrossRef] [Green Version]
- Moshirfar, M.; Murri, M.S.; Shah, T.J.; Skanchy, D.F.; Tuckfield, J.Q.; Ronquillo, Y.C.; Birdsong, O.C.; Hofstedt, D.; Hoopes, P.C. A Review of Corneal Endotheliitis and Endotheliopathy: Differential Diagnosis, Evaluation, and Treatment. Ophthalmology 2019, 8, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, P.; Welander, P.; Han, X.; Cantin, E. Herpes simplex virus type 1 DNA is immunostimulatory in vitro and in vivo. J. Virol. 2003, 77, 11158–11169. [Google Scholar] [CrossRef] [Green Version]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of herpesviruses by the innate immune system. Nat. Rev. Immunol. 2011, 11, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Neeper, M.; Schmidt, A.M.; Brett, J.; Yan, S.D.; Wang, F.; Pan, Y.C.; Elliston, K.; Stern, D.; Shaw, A. Cloning and expression of a cell surface receptor for advanced glycosylation end products of proteins. J. Biol. Chem. 1992, 267, 14998–15004. [Google Scholar] [CrossRef]
- Inoue, Y. Review of clinical and basic approaches to corneal endotheliitis. Cornea 2014, 33 (Suppl. 11), S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Inata, K.; Miyazaki, D.; Uotani, R.; Shimizu, D.; Miyake, A.; Shimizu, Y.; Inoue, Y. Effectiveness of real-time PCR for diagnosis and prognosis of varicella-zoster virus keratitis. Jpn. J. Ophthalmol. 2018, 62, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Danby, F.W. Nutrition and aging skin: Sugar and glycation. Clin Derm. 2010, 28, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Kaji, Y.; Amano, S.; Usui, T.; Oshika, T.; Yamashiro, K.; Ishida, S.; Suzuki, K.; Tanaka, S.; Adamis, A.P.; Nagai, R.; et al. Expression and function of receptors for advanced glycation end products in bovine corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 521–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, P.; Chaudhry, N.; Mittal, R.; Mukherjee, T.K. Role of receptor for advanced glycation end products in the complication and progression of various types of cancers. Biochim. Biophys. Acta 2015, 1850, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, Z.A.; Armour, C.L.; Phipps, S.; Sukkar, M.B. RAGE and TLRs: Relatives, friends or neighbours? Mol. Immunol. 2013, 56, 739–744. [Google Scholar] [CrossRef]
- Tian, J.; Avalos, A.M.; Mao, S.Y.; Chen, B.; Senthil, K.; Wu, H.; Parroche, P.; Drabic, S.; Golenbock, D.; Sirois, C.; et al. Toll-like receptor 9-dependent activation by DNA-containing immune complexes is mediated by HMGB1 and RAGE. Nat. Immunol. 2007, 8, 487–496. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, C.D.; Shukla, D. The Importance of Heparan Sulfate in Herpesvirus Infection. Virol. Sin. 2008, 23, 383–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Young, J.; Song, D.; Esko, J.D. Heparan sulfate is essential for high mobility group protein 1 (HMGB1) signaling by the receptor for advanced glycation end products (RAGE). J. Biol. Chem. 2011, 286, 41736–41744. [Google Scholar] [CrossRef] [Green Version]
- Sirois, C.M.; Jin, T.; Miller, A.L.; Bertheloot, D.; Nakamura, H.; Horvath, G.L.; Mian, A.; Jiang, J.; Schrum, J.; Bossaller, L.; et al. RAGE is a nucleic acid receptor that promotes inflammatory responses to DNA. J. Exp. Med. 2013, 210, 2447–2463. [Google Scholar] [CrossRef] [Green Version]
- Bertheloot, D.; Naumovski, A.L.; Langhoff, P.; Horvath, G.L.; Jin, T.; Xiao, T.S.; Garbi, N.; Agrawal, S.; Kolbeck, R.; Latz, E. RAGE Enhances TLR Responses through Binding and Internalization of RNA. J. Immunol. 2016, 197, 4118–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, D.; Uotani, R.; Inoue, M.; Haruki, T.; Shimizu, Y.; Yakura, K.; Yamagami, S.; Suzutani, T.; Hosogai, M.; Isomura, H.; et al. Corneal endothelial cells activate innate and acquired arm of anti-viral responses after cytomegalovirus infection. Exp. Eye Res. 2017, 161, 143–152. [Google Scholar] [CrossRef]
- Yu, Q.; Katlinskaya, Y.V.; Carbone, C.J.; Zhao, B.; Katlinski, K.V.; Zheng, H.; Guha, M.; Li, N.; Chen, Q.; Yang, T.; et al. DNA-damage-induced type I interferon promotes senescence and inhibits stem cell function. Cell Rep. 2015, 11, 785–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Yamada, Y.; Ma, J.; Kezuka, T.; Hamada, H.; Usui, T.; Mochizuki, M.; et al. Human corneal endothelial cells expressing programmed death-ligand 1 (PD-L1) suppress PD-1+ T helper 1 cells by a contact-dependent mechanism. Investig. Ophthalmol. Vis. Sci. 2009, 50, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Sugita, S.; Horie, S.; Yamagami, S.; Mochizuki, M. Mechanisms of immune suppression for CD8+ T cells by human corneal endothelial cells via membrane-bound TGFbeta. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2548–2557. [Google Scholar] [CrossRef] [Green Version]
- Ballas, Z.K.; Rasmussen, W.L.; Krieg, A.M. Induction of NK activity in murine and human cells by CpG motifs in oligodeoxynucleotides and bacterial DNA. J. Immunol. 1996, 157, 1840–1845. [Google Scholar] [PubMed]
- Krug, A.; Rothenfusser, S.; Hornung, V.; Jahrsdorfer, B.; Blackwell, S.; Ballas, Z.K.; Endres, S.; Krieg, A.M.; Hartmann, G. Identification of CpG oligonucleotide sequences with high induction of IFN-alpha/beta in plasmacytoid dendritic cells. Eur. J. Immunol. 2001, 31, 2154–2163. [Google Scholar] [CrossRef]
- Vollmer, J.; Weeratna, R.; Payette, P.; Jurk, M.; Schetter, C.; Laucht, M.; Wader, T.; Tluk, S.; Liu, M.; Davis, H.L.; et al. Characterization of three CpG oligodeoxynucleotide classes with distinct immunostimulatory activities. Eur. J. Immunol. 2004, 34, 251–262. [Google Scholar] [CrossRef]
- Rothenfusser, S.; Hornung, V.; Ayyoub, M.; Britsch, S.; Towarowski, A.; Krug, A.; Sarris, A.; Lubenow, N.; Speiser, D.; Endres, S.; et al. CpG-A and CpG-B oligonucleotides differentially enhance human peptide-specific primary and memory CD8+ T-cell responses in vitro. Blood 2004, 103, 2162–2169. [Google Scholar] [CrossRef]
- Ayash-Rashkovsky, M.; Borkow, G.; Davis, H.L.; Moss, R.B.; Bartholomew, R.; Bentwich, Z. Enhanced HIV-1 specific immune response by CpG ODN and HIV-1 immunogen-pulsed dendritic cells confers protection in the Trimera murine model of HIV-1 infection. Faseb J. 2005, 19, 1152–1154. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, Y.; Miyazaki, D.; Yakura, K.; Haruki, T.; Inoue, Y. Induction of IL-6 in transcriptional networks in corneal epithelial cells after herpes simplex virus type 1 infection. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2441–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyazaki, D.; Kandori-Inoue, M.; Shimizu, Y.; Ohtani, F.; Chono, I.; Inoue, Y.; Yamagami, S. Role Played by Receptors for Advanced Glycosylation End Products in Corneal Endothelial Cells after HSV-1 Infection. Int. J. Mol. Sci. 2021, 22, 5833. https://doi.org/10.3390/ijms22115833
Miyazaki D, Kandori-Inoue M, Shimizu Y, Ohtani F, Chono I, Inoue Y, Yamagami S. Role Played by Receptors for Advanced Glycosylation End Products in Corneal Endothelial Cells after HSV-1 Infection. International Journal of Molecular Sciences. 2021; 22(11):5833. https://doi.org/10.3390/ijms22115833
Chicago/Turabian StyleMiyazaki, Dai, Michiko Kandori-Inoue, Yumiko Shimizu, Fumie Ohtani, Ikuyo Chono, Yoshitsugu Inoue, and Satoru Yamagami. 2021. "Role Played by Receptors for Advanced Glycosylation End Products in Corneal Endothelial Cells after HSV-1 Infection" International Journal of Molecular Sciences 22, no. 11: 5833. https://doi.org/10.3390/ijms22115833
APA StyleMiyazaki, D., Kandori-Inoue, M., Shimizu, Y., Ohtani, F., Chono, I., Inoue, Y., & Yamagami, S. (2021). Role Played by Receptors for Advanced Glycosylation End Products in Corneal Endothelial Cells after HSV-1 Infection. International Journal of Molecular Sciences, 22(11), 5833. https://doi.org/10.3390/ijms22115833