
Proteome Changes Reveal the Protective Roles of Exogenous Citric Acid in Alleviating Cu Toxicity in Brassica napus L.

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
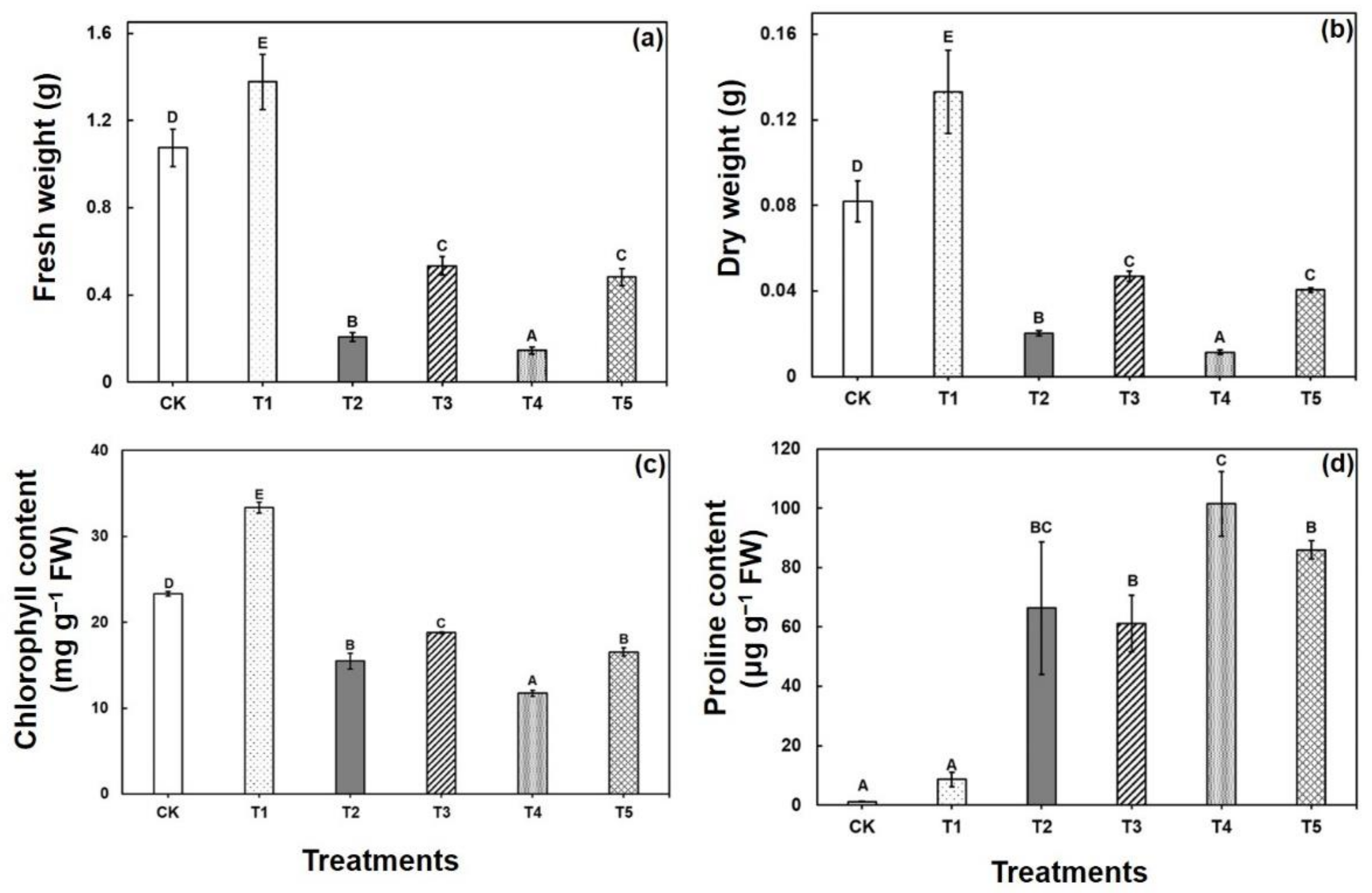
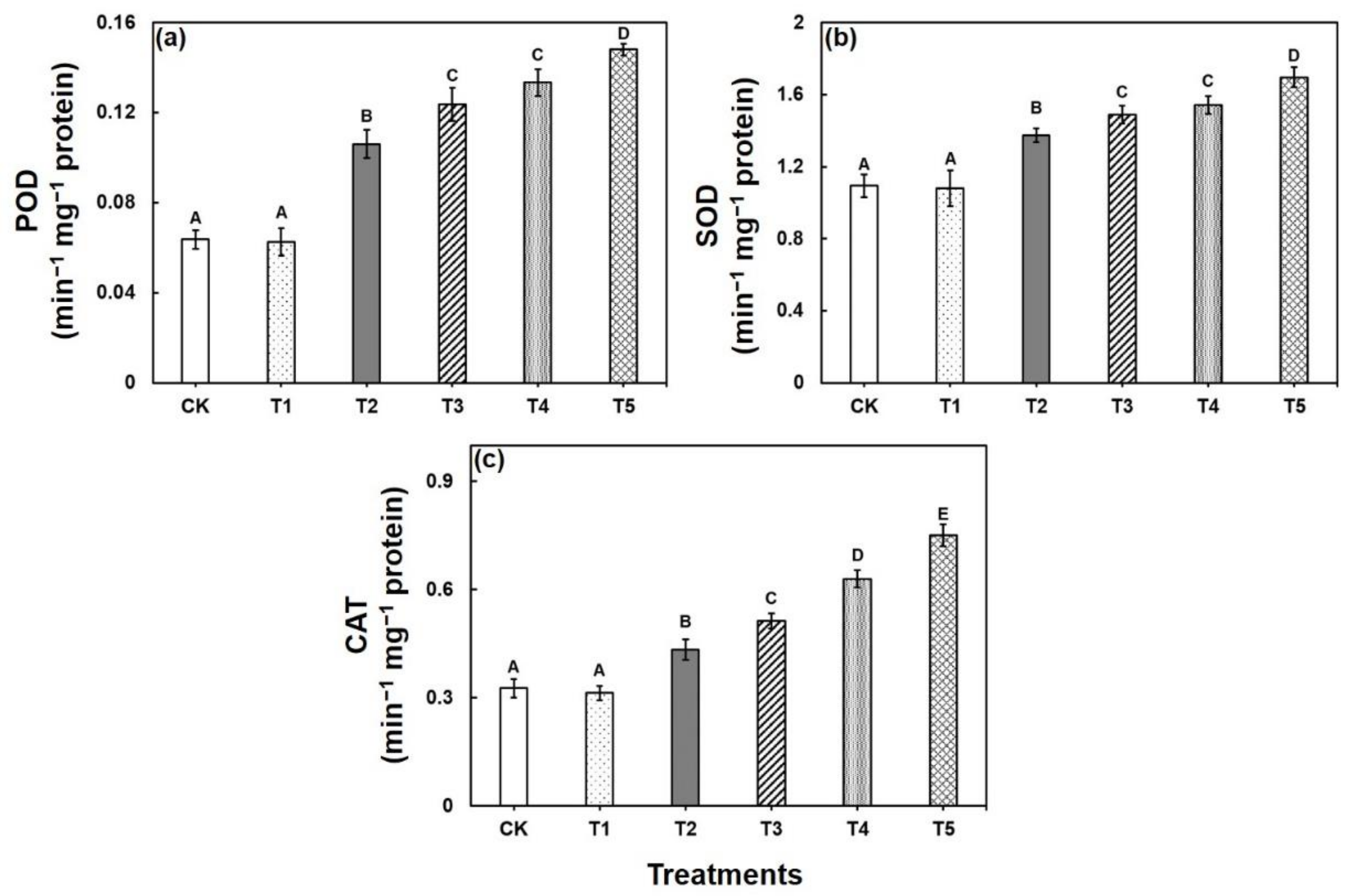
2.1. Responses of Morphological, and Biochemical Responses to CA-Mediated Cu Stress in B. napus
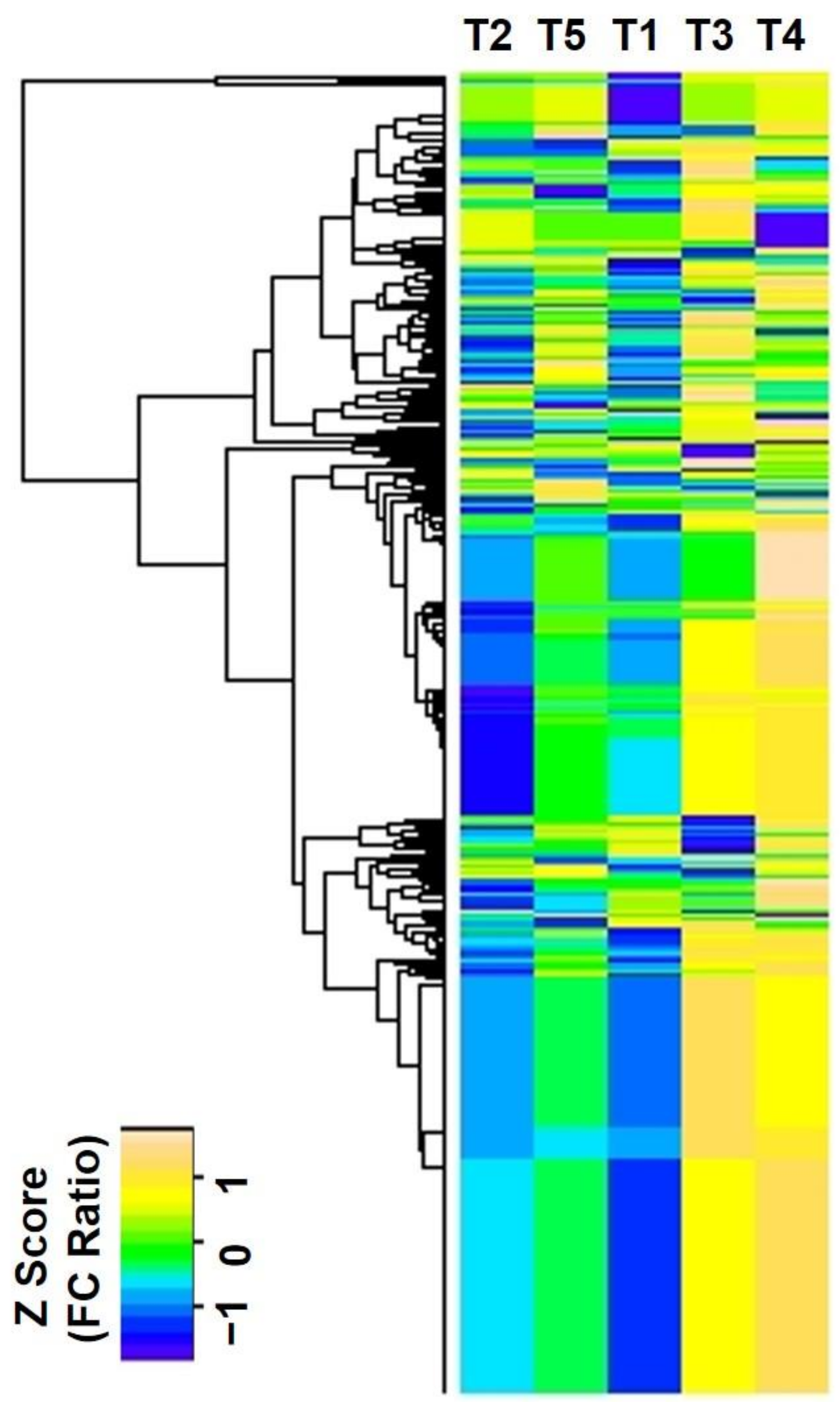
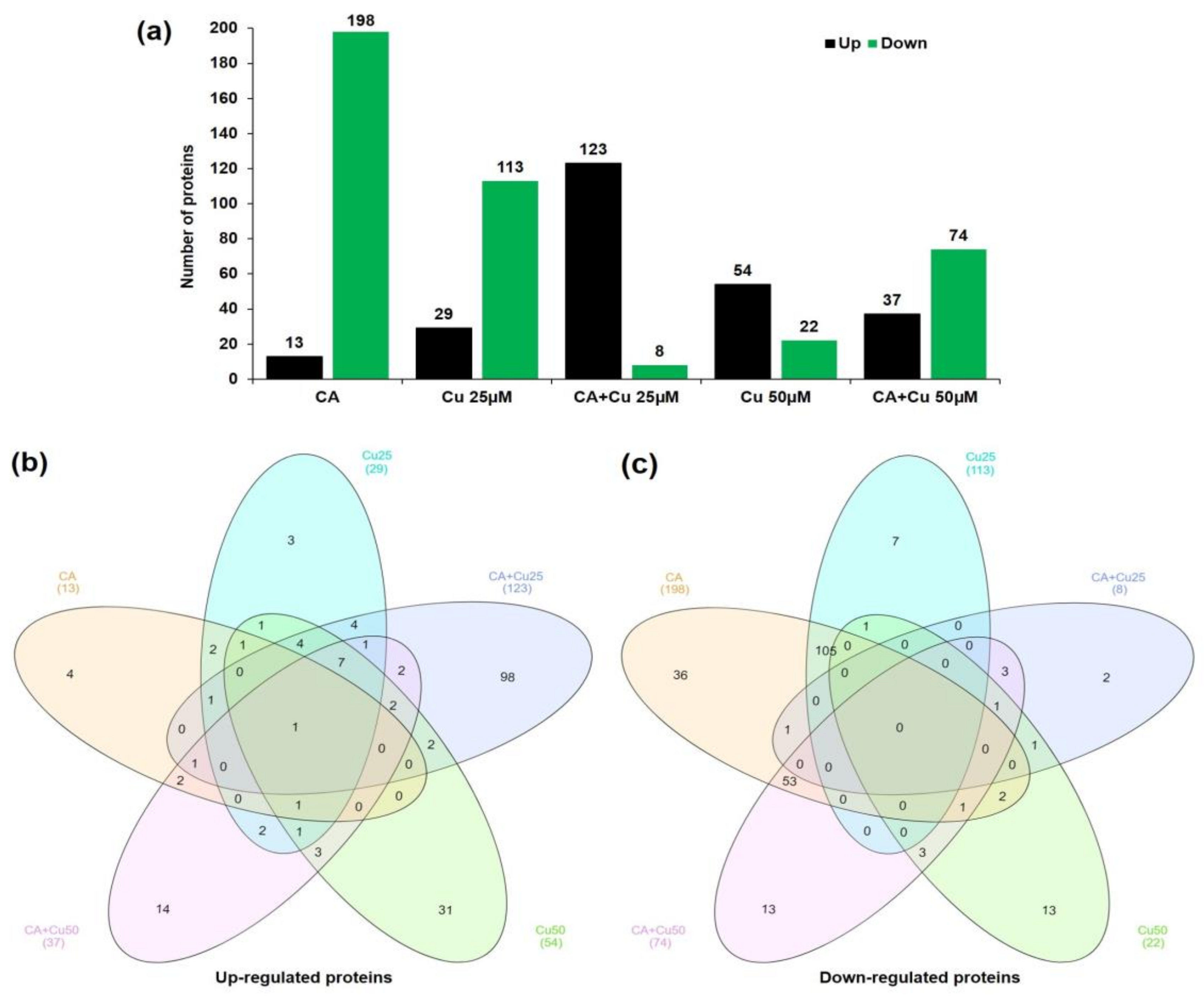
2.2. Changes of CA-Mediated Cu Stress on Proteome
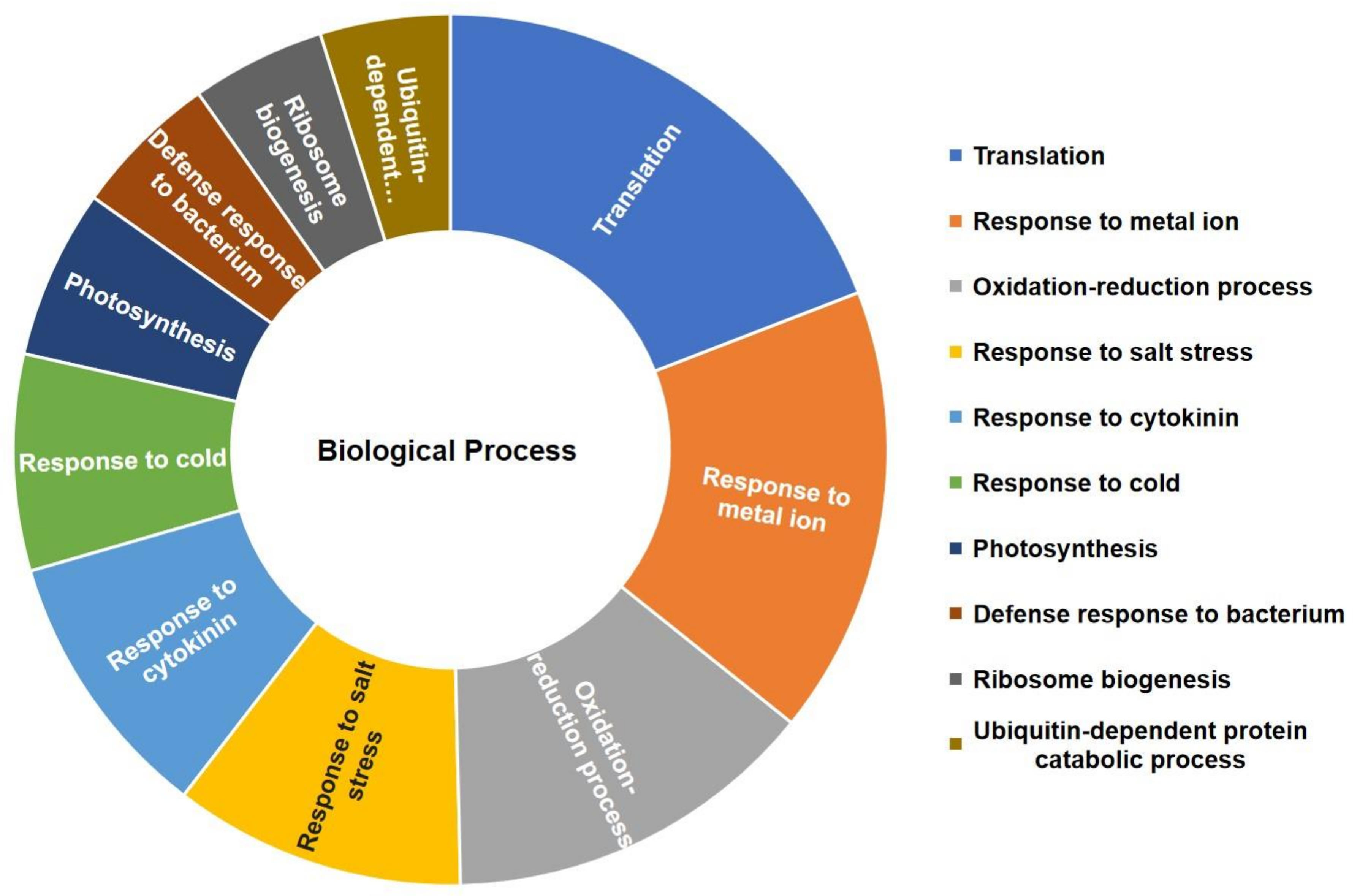
2.3. GO Analysis of the Total Identified Proteins in CA-Mediated Cu Stressed Plants
2.4. GO Analysis of the DAPs in CA-Mediated Cu Stressed Plants
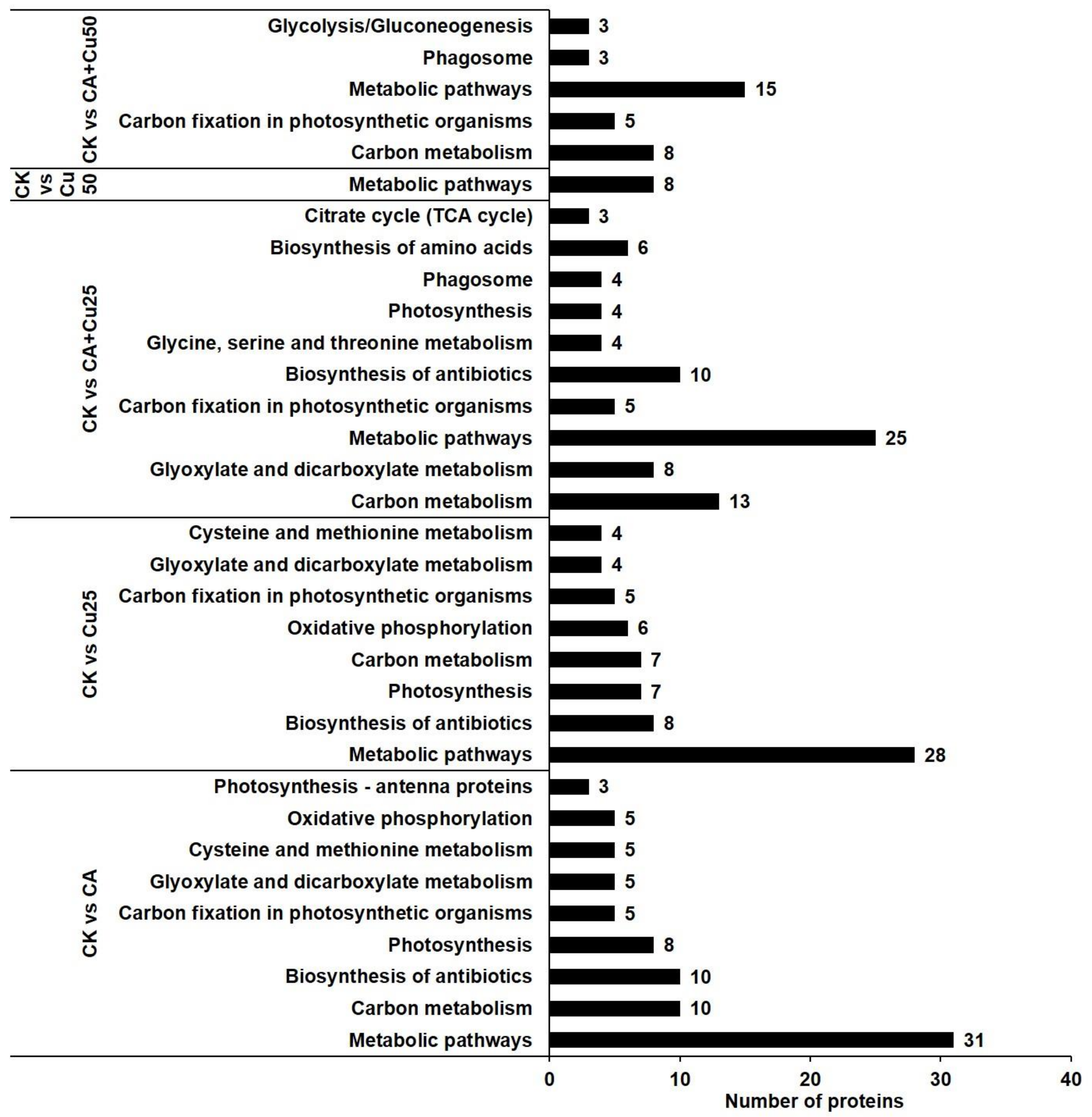
2.5. Potential Metabolic Pathways of the DAPs
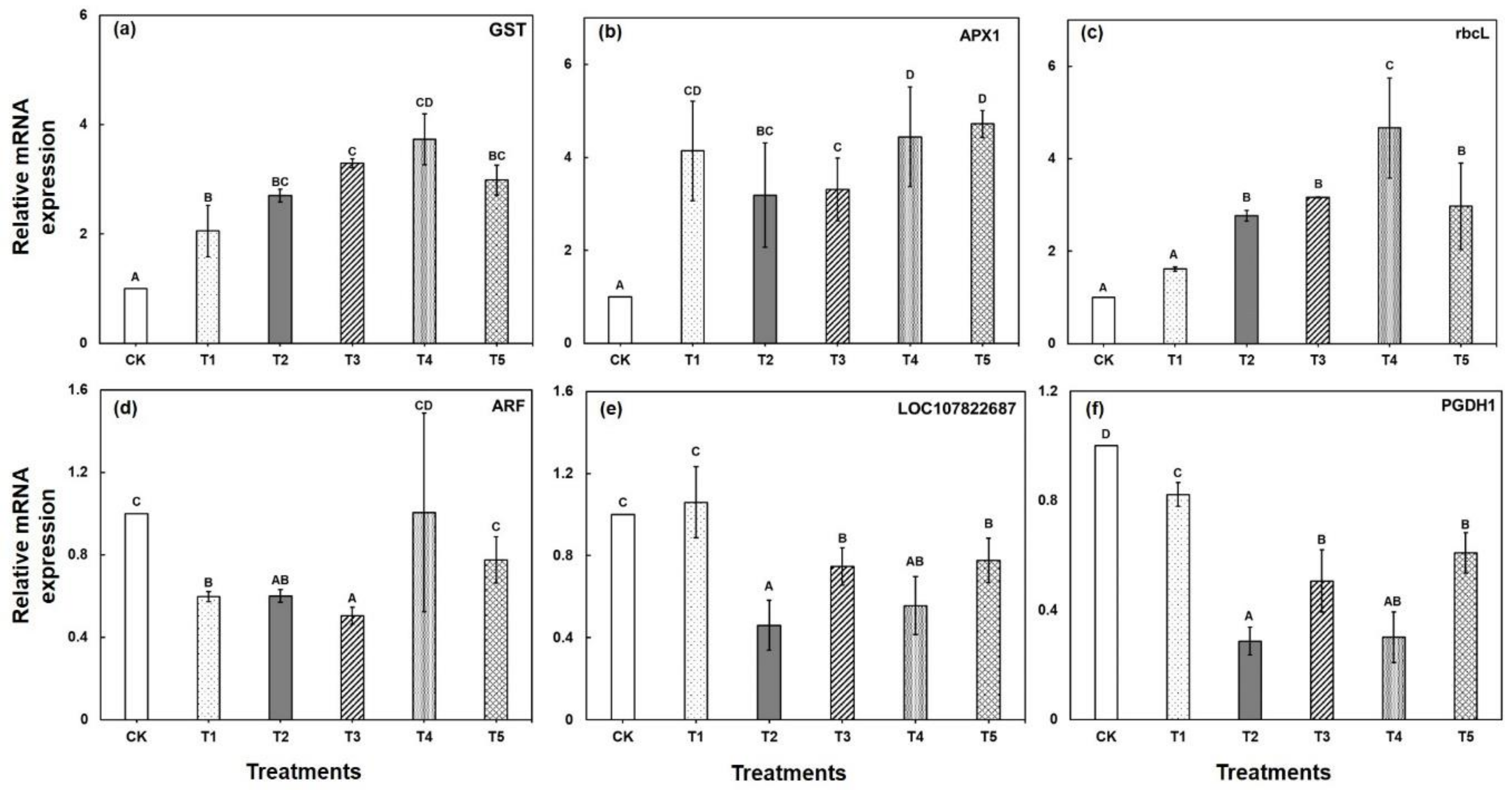
2.6. Validation of the Candidate Proteins by qRT-PCR
3. Discussion
3.1. Cu Stress Altered Energy and Carbohydrate Metabolism
3.2. Cu Stress Impairs the CO2 Assimilation and Photosynthesis
3.3. Cu Stress Induces Changes in Signal Transduction and Protein Metabolism
3.4. Cu Stress Induces Stress-and Defense-Related Proteins
3.5. Proteins Participate in Sulfur Assimilation and Heavy Metal Detoxification
3.6. Proteins Related to Cytoskeleton and Cell Wall-Related Proteins
3.7. Development
4. Materials and Methods
4.1. Plant Materials, Growing Conditions, and Stress Treatments
4.2. Growth Parameters
4.3. Measurement of Biochemical Parameters
4.4. Estimation of Antioxidant Enzyme Activities
4.5. Protein Extraction
4.6. Purification, Digestion of Extracted Proteins
4.7. LC-MS/MS Analysis
4.8. Database Search and Protein Quantification
4.9. Bioinformatics Analysis of the Identified Proteins
4.10. RNA Isolation, cDNA Synthesis, and Gene Expression Analysis by qRT-PCR
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sarwar, N.; Ishaq, W.; Farid, G.; Shaheen, M.R.; Imran, M.; Geng, M.; Hussain, S. Zinc–cadmium interactions: Impact on wheat physiology and mineral acquisition. Ecotoxicol. Environ. Saf. 2015, 122, 528–536. [Google Scholar] [CrossRef]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [Green Version]
- Printz, B.; Lutts, S.; Hausman, J.-F.; Sergeant, K. Copper trafficking in plants and its implication on cell wall dynamics. Front. Plant Sci. 2016, 7, 601. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, J.; Wang, X.; Xia, Y.; Chen, C.; Shen, Z.; Chen, Y. Proteomic analysis on roots of Oenothera glazioviana under copper-stress conditions. Sci. Rep. 2017, 7, 10589. [Google Scholar] [CrossRef] [Green Version]
- Bona, E.; Marsano, F.; Cavaletto, M.; Berta, G. Proteomic characterization of copper stress response in Cannabis sativa roots. Proteomics 2007, 7, 1121–1130. [Google Scholar] [CrossRef]
- Nowicka, B.; Pluciński, B.; Kuczyńska, P.; Kruk, J. Physiological characterization of Chlamydomonas reinhardtii acclimated to chronic stress induced by Ag, Cd, Cr, Cu and Hg ions. Ecotoxicol. Environ. Saf. 2016, 130, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Huang, Y.; Paglia, K.; Vaniya, A.; Wancewicz, B.; Keller, A.A. Metabolomics reveals the molecular mechanisms of copper induced cucumber leaf (Cucumis sativus) senescence. Environ. Sci. Technol. 2018, 52, 7092–7100. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cutright, T. EDTA and HEDTA effects on Cd, Cr, and Ni uptake by Helianthus annuus. Chemosphere 2001, 45, 21–28. [Google Scholar] [CrossRef]
- Liphadzi, M.; Kirkham, M. Availability and plant uptake of heavy metals in EDTA-assisted phytoremediation of soil and composted biosolids. S. Afr. J. Bot. 2006, 72, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, I.E.; Ali, S.; Rizwan, M.; Farid, M.; Shakoor, M.B.; Gill, R.A.; Najeeb, U.; Iqbal, N.; Ahmad, R. Citric acid assisted phytoremediation of copper by Brassica napus L. Ecotoxicol. Environ. Saf. 2015, 120, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Nan, Z.; Liu, X.; Li, Y.; Qin, S.; Ding, H. Accumulation and bioavailability of copper and nickel in wheat plants grown in contaminated soils from the oasis, northwest China. Geoderma 2009, 152, 290–295. [Google Scholar] [CrossRef]
- Afshan, S.; Ali, S.; Bharwana, S.A.; Rizwan, M.; Farid, M.; Abbas, F.; Ibrahim, M.; Mehmood, M.A.; Abbasi, G.H. Citric acid enhances the phytoextraction of chromium, plant growth, and photosynthesis by alleviating the oxidative damages in Brassica napus L. Environ. Sci. Pollut. Res. 2015, 22, 11679–11689. [Google Scholar] [CrossRef]
- Freitas, E.V.; Nascimento, C.W.; Souza, A.; Silva, F.B. Citric acid-assisted phytoextraction of lead: A field experiment. Chemosphere 2013, 92, 213–217. [Google Scholar] [CrossRef]
- Gao, Y.; Miao, C.; Mao, L.; Zhou, P.; Jin, Z.; Shi, W. Improvement of phytoextraction and antioxidative defense in Solanum nigrum L. under cadmium stress by application of cadmium-resistant strain and citric acid. J. Hazard. Mater. 2010, 181, 771–777. [Google Scholar] [CrossRef]
- Thounaojam, T.C.; Panda, P.; Mazumdar, P.; Kumar, D.; Sharma, G.D.; Sahoo, L.; Sanjib, P. Excess copper induced oxidative stress and response of antioxidants in rice. Plant Physiol. Biochem. 2012, 53, 33–39. [Google Scholar] [CrossRef]
- Al Mahmud, J.; Hasanuzzaman, M.; Nahar, K.; Bhuyan, M.B.; Fujita, M. Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: Coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol. Environ. Saf. 2018, 147, 990–1001. [Google Scholar] [CrossRef]
- Luo, Z.-B.; He, J.; Polle, A.; Rennenberg, H. Heavy metal accumulation and signal transduction in herbaceous and woody plants: Paving the way for enhancing phytoremediation efficiency. Biotechnol. Adv. 2016, 34, 1131–1148. [Google Scholar] [CrossRef]
- Sarma, R.S.; Prakash, P. Adverse effect of heavy metal toxicity in plants’ metabolic systems and biotechnological approaches for its tolerance mechanism. In New Frontiers in Stress Management for Durable Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 145–168. [Google Scholar]
- Benyó, D.; Horváth, E.; Németh, E.; Leviczky, T.; Takács, K.; Lehotai, N.; Feigl, G.; Kolbert, Z.; Ördög, A.; Gallé, R. Physiological and molecular responses to heavy metal stresses suggest different detoxification mechanism of Populus deltoides and P. x canadensis. J. Plant Physiol. 2016, 201, 62–70. [Google Scholar] [CrossRef]
- Kumar, P.; Srivastava, D.K. Molecular and Biotechnological Interventions for Improving Brassicaceae Crops for Abiotic Stress Tolerance. In The Plant Family Brassicaceae; Springer: Berlin/Heidelberg, Germany, 2020; pp. 437–450. [Google Scholar]
- Han, Y.; Zhang, L.; Gu, J.; Zhao, J.; Fu, J. Citric acid and EDTA on the growth, photosynthetic properties and heavy metal accumulation of Iris halophila Pall. cultivated in Pb mine tailings. Int. Biodeterior. Biodegradation 2018, 128, 15–21. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Trinh, N.N.; Fu, S.-F.; Hsiung, Y.-C.; Chia, L.-C.; Lin, C.-W.; Huang, H.-J. Comparison of early transcriptome responses to copper and cadmium in rice roots. Plant Mol. Biol. 2013, 81, 507–522. [Google Scholar] [CrossRef]
- Shakoor, M.B.; Ali, S.; Hameed, A.; Farid, M.; Hussain, S.; Yasmeen, T.; Najeeb, U.; Bharwana, S.A.; Abbasi, G.H. Citric acid improves lead (Pb) phytoextraction in Brassica napus L. by mitigating Pb-induced morphological and biochemical damages. Ecotoxicol. Environ. Saf. 2014, 109, 38–47. [Google Scholar] [CrossRef]
- Sudo, E.; Itouga, M.; Yoshida-Hatanaka, K.; Ono, Y.; Sakakibara, H. Gene expression and sensitivity in response to copper stress in rice leaves. J. Exp. Bot. 2008, 59, 3465–3474. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Lu, Q.; Cao, Y.; Wang, M.; Cheng, X.; Yan, Q. Comparative transcriptome analysis of the molecular mechanism of the hairy roots of Brassica campestris L. in response to cadmium stress. Int. J. Mol. Sci. 2020, 21, 180. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.-R.; Ikka, T.; Sawaki, Y.; Kobayashi, Y.; Suzuki, Y.; Hibino, T.; Sato, S.; Sakurai, N.; Shibata, D.; Koyama, H. Comparative transcriptomic characterization of aluminum, sodium chloride, cadmium and copper rhizotoxicities in Arabidopsis thaliana. BMC Plant Biol. 2009, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Gong, F.; Cao, D.; Hu, X.; Wang, W. Advances in crop proteomics: PTMs of proteins under abiotic stress. Proteomics 2016, 16, 847–865. [Google Scholar] [CrossRef]
- Farooq, M.A.; Zhang, K.; Islam, F.; Wang, J.; Athar, H.U.; Nawaz, A.; Ullah Zafar, Z.; Xu, J.; Zhou, W. Physiological and iTRAQ-Based Quantitative Proteomics Analysis of Methyl Jasmonate–Induced Tolerance in Brassica napus Under Arsenic Stress. Proteomics 2018, 18, 1700290. [Google Scholar] [CrossRef] [PubMed]
- Jian, M.; Zhang, D.; Wang, X.; Wei, S.; Zhao, Y.; Ding, Q.; Han, Y.; Ma, L. Differential expression pattern of the proteome in response to cadmium stress based on proteomics analysis of wheat roots. BMC Genom. 2020, 21, 343. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, J.; Li, Y.-F.; Xu, X.; Zhang, B.; Liu, Y.; Cui, L.; Li, B.; Gao, Y.; Chai, Z. Comparative metalloproteomic approaches for the investigation proteins involved in the toxicity of inorganic and organic forms of mercury in rice (Oryza sativa L.) roots. Metallomics 2016, 8, 663–671. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, Y.; Wang, Y.; Wang, W.; Gu, C.; Huang, S.; Yuan, H.; Dhankher, O.P. Quantitative proteomic analysis reveals complex regulatory and metabolic response of Iris lactea Pall. var. chinensis to cadmium toxicity. J. Hazard. Mater. 2020, 400, 123165. [Google Scholar] [CrossRef] [PubMed]
- Szuba, A.; Marczak, Ł.; Kozłowski, R. Role of the proteome in providing phenotypic stability in control and ectomycorrhizal poplar plants exposed to chronic mild Pb stress. Environ. Pollut. 2020, 264, 114585. [Google Scholar] [CrossRef]
- Li, F.; Shi, J.; Shen, C.; Chen, G.; Hu, S.; Chen, Y. Proteomic characterization of copper stress response in Elsholtzia splendens roots and leaves. Plant. Mol. Biol. 2009, 71, 251–263. [Google Scholar] [CrossRef]
- Li, G.; Peng, X.; Xuan, H.; Wei, L.; Yang, Y.; Guo, T.; Kang, G. Proteomic analysis of leaves and roots of common wheat (Triticum aestivum L.) under copper-stress conditions. J. Proteome Res. 2013, 12, 4846–4861. [Google Scholar] [CrossRef]
- Roy, S.K.; Kwon, S.J.; Cho, S.-W.; Kamal, A.H.M.; Kim, S.-W.; Sarker, K.; Oh, M.-W.; Lee, M.-S.; Chung, K.-Y.; Xin, Z. Leaf proteome characterization in the context of physiological and morphological changes in response to copper stress in sorghum. BioMetals 2016, 29, 495–513. [Google Scholar] [CrossRef]
- Wu, J.; Hu, J.; Wang, L.; Zhao, L.; Ma, F. Responses of Phragmites australis to copper stress: A combined analysis of plant morphology, physiology and proteomics. Plant. Biol. 2020, 23, 351–362. [Google Scholar] [CrossRef]
- Bandehagh, A.; Salekdeh, G.H.; Toorchi, M.; Mohammadi, A.; Komatsu, S. Comparative proteomic analysis of canola leaves under salinity stress. Proteomics 2011, 11, 1965–1975. [Google Scholar] [CrossRef]
- Chen, S.; Ding, G.; Wang, Z.; Cai, H.; Xu, F. Proteomic and comparative genomic analysis reveals adaptability of Brassica napus to phosphorus-deficient stress. J. Proteom. 2015, 117, 106–119. [Google Scholar] [CrossRef]
- Chu, P.; Yan, G.X.; Yang, Q.; Zhai, L.N.; Zhang, C.; Zhang, F.Q.; Guan, R.Z. iTRAQ-based quantitative proteomics analysis of Brassica napus leaves reveals pathways associated with chlorophyll deficiency. J. Proteom. 2015, 113, 244–259. [Google Scholar] [CrossRef]
- D’Hooghe, P.; Dubousset, L.; Gallardo, K.; Kopriva, S.; Avice, J.-C.; Trouverie, J. Evidence for proteomic and metabolic adaptations associated with alterations of seed yield and quality in sulfur-limited Brassica napus L. Mol. Cell. Proteom. 2014, 13, 1165–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismaili, A.; Salavati, A.; Pour Mohammadi, P. A comparative proteomic analysis of responses to high temperature stress in hypocotyl of canola (Brassica napus L.). Protein Pept. Lett. 2015, 22, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Shao, M.; He, Y.; Guan, R.; Chu, P.; Jiang, H. Proteome dynamics and physiological responses to short-term salt stress in Brassica napus leaves. PLoS ONE 2015, 10, e0144808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maryan, K.E.; Lahiji, H.S.; Farrokhi, N.; Haynes, P.A.; Hamzelou, S.; Komeleh, H.H. Comparative Leaf Proteomics of Brassica napus Genotypes with Distinctive Levels of Early Cold Acclimation. Plant. Mol. Biol. Rep. 2020, 1–18. [Google Scholar] [CrossRef]
- Yang, P.; Li, Y.; He, C.; Yan, J.; Zhang, W.; Li, X.; Xiang, F.; Zuo, Z.; Li, X.; Zhu, Y. Phenotype and TMT-based quantitative proteomics analysis of Brassica napus reveals new insight into chlorophyll synthesis and chloroplast structure. J. Proteom. 2020, 214, 103621. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Simons, B.; Zhu, N.; Oppenheimer, D.G.; Chen, S. Analysis of abscisic acid responsive proteins in Brassica napus guard cells by multiplexed isobaric tagging. J. Proteom. 2010, 73, 790–805. [Google Scholar] [CrossRef]
- Zhu, M.; Zhu, N.; Song, W.y.; Harmon, A.C.; Assmann, S.M.; Chen, S. Thiol-based redox proteins in abscisic acid and methyl jasmonate signaling in Brassica napus guard cells. Plant. J. 2014, 78, 491–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, J.; Chen, G.; Yoo, M.-J.; Zhu, N.; Dufresne, D.; Erickson, J.E.; Shao, H.; Chen, S. Comparative proteomic analysis of Brassica napus in response to drought stress. J. Proteome Res. 2015, 14, 3068–3081. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Luo, Z.; Huang, X.; Zhang, L.; Zhao, P.; Ma, H.; Li, X.; Ban, Z.; Liu, X. Label-free quantitative proteomics to investigate strawberry fruit proteome changes under controlled atmosphere and low temperature storage. J. Proteom. 2015, 120, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-S.; Zhang, Z. Proteomic changes in the xylem sap of Brassica napus under cadmium stress and functional validation. BMC Plant. Biol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhuyan, M.B.; Oku, H.; Fujita, M. Exogenous nitric oxide pretreatment protects Brassica napus L. seedlings from paraquat toxicity through the modulation of antioxidant defense and glyoxalase systems. Plant. Physiol. Biochem. 2018, 126, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Lacalle, R.G.; Gómez-Sagasti, M.T.; Artetxe, U.; Garbisu, C.; Becerril, J.M. Brassica napus has a key role in the recovery of the health of soils contaminated with metals and diesel by rhizoremediation. Sci. Total Environ. 2018, 618, 347–356. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, Z.; Xiang, Z.; Yang, Z. Exogenous application of citric acid ameliorates the adverse effect of heat stress in tall fescue (Lolium arundinaceum). Front. Plant. Sci. 2016, 7, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parveen, A.; Saleem, M.H.; Kamran, M.; Haider, M.Z.; Chen, J.-T.; Malik, Z.; Rana, M.S.; Hassan, A.; Hur, G.; Javed, M.T.; et al. Effect of citric acid on growth, ecophysiology, chloroplast ultrastructure, and phytoremediation potential of jute (Corchorus capsularis L.) seedlings exposed to copper stress. Biomolecules 2020, 10, 592. [Google Scholar] [CrossRef]
- Choudhary, S.P.; Oral, H.V.; Bhardwaj, R.; Yu, J.-Q.; Tran, L.-S.P. Interaction of brassinosteroids and polyamines enhances copper stress tolerance in Raphanus sativus. J. Exp. Bot. 2012, 63, 5659–5675. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.; Filgueiras, L.; Santos, B.; Coelho, M.; Silva, M.; Estrada-Bonilla, G.; Vidal, M.; Baldani, J.I.; Meneses, C. Gluconacetobacter diazotrophicus changes the molecular mechanisms of root development in Oryza sativa L. growing under water stress. Int. J. Mol. Sci. 2020, 21, 333. [Google Scholar] [CrossRef] [Green Version]
- Cornu, J.Y.; Huguenot, D.; Jézéquel, K.; Lollier, M.; Lebeau, T. Bioremediation of copper-contaminated soils by bacteria. World J. Microbiol. Biotechnol. 2017, 33, 26. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Seraj, Z.I.; Fujita, M. Interactive effects of nitric oxide and glutathione in mitigating copper toxicity of rice (Oryza sativa L.) seedlings. Plant. Signal. Behav. 2015, 10, e991570. [Google Scholar] [CrossRef] [Green Version]
- Drążkiewicz, M.; Skórzyńska-Polit, E.; Krupa, Z. Response of the ascorbate–glutathione cycle to excess copper in Arabidopsis thaliana (L.). Plant. Sci. 2003, 164, 195–202. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Sami, F.; Siddiqui, H.; Hayat, S. Supplementation of salicylic acid and citric acid for alleviation of cadmium toxicity to Brassica juncea. J. Plant. Growth Regul. 2020, 39, 641–655. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Xu, X.Y.; Gong, Q.Q.; Xie, C.; Fan, W.; Yang, J.L.; Lin, Q.S.; Zheng, S.J. Root proteome of rice studied by iTRAQ provides integrated insight into aluminum stress tolerance mechanisms in plants. J. Proteom. 2014, 98, 189–205. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Y.; Zhang, F.; Tang, M.; Chen, Y.; Wang, J.; Karanja, B.K.; Luo, X.; Zhang, W.; Liu, L. Dissecting root proteome changes reveals new insight into cadmium stress response in radish (Raphanus sativus L.). Plant. Cell Physiol. 2017, 58, 1901–1913. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Jwa, N.-S. Understanding the responses of rice to environmental stress using proteomics. J. Proteome Res. 2013, 12, 4652–4669. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Taamalli, M.; Gevi, F.; Timperio, A.M.; Zolla, L.; Ghnaya, T. Cadmium stress responses in Brassica juncea: Hints from proteomics and metabolomics. J. Proteome Res. 2013, 12, 4979–4997. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, S.; Wang, J.; Su, X.; Suo, B.; Qin, F.; Zhao, H. Exogenous application of 5-aminolevulinic acid on wheat seedlings under drought stress enhances the transcription of psbA and psbD genes and improves photosynthesis. Braz. J. Bot. 2018, 41, 275–285. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant. Growth Regul. 2019, 1–23. [Google Scholar] [CrossRef]
- Huang, J.; Wu, X.; Tian, F.; Chen, Q.; Luo, P.; Zhang, F.; Wan, X.; Zhong, Y.; Liu, Q.; Lin, T. Changes in proteome and protein phosphorylation reveal the protective roles of exogenous nitrogen in alleviating cadmium toxicity in poplar plants. Int. J. Mol. Sci. 2020, 21, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Okay, S.; Yildirim, V.; Büttner, K.; Becher, D.; Özcengiz, G. Dynamic proteomic analysis of Phanerochaete chrysosporium under copper stress. Ecotoxicol. Environ. Saf. 2020, 198, 110694. [Google Scholar] [CrossRef]
- Sobhanian, H.; Razavizadeh, R.; Nanjo, Y.; Ehsanpour, A.A.; Jazii, F.R.; Motamed, N.; Komatsu, S. Proteome analysis of soybean leaves, hypocotyls and roots under salt stress. Proteome Sci. 2010, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Cui, J.; Zhang, H.; Wang, G.; Zhao, F.-J.; Shen, Z. Proteomic analysis of copper stress responses in the roots of two rice (Oryza sativa L.) varieties differing in Cu tolerance. Plant. Soil 2013, 366, 647–658. [Google Scholar] [CrossRef]
- Mendoza-Cozatl, D.; Loza-Tavera, H.; Hernández-Navarro, A.; Moreno-Sánchez, R. Sulfur assimilation and glutathione metabolism under cadmium stress in yeast, protists and plants. FEMS Microbiol. Rev. 2005, 29, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Ritter, A.; Ubertini, M.; Romac, S.; Gaillard, F.; Delage, L.; Mann, A.; Cock, J.M.; Tonon, T.; Correa, J.A.; Potin, P. Copper stress proteomics highlights local adaptation of two strains of the model brown alga Ectocarpus siliculosus. Proteomics 2010, 10, 2074–2088. [Google Scholar] [CrossRef]
- Gutsch, A.; Zouaghi, S.; Renaut, J.; Cuypers, A.; Hausman, J.-F.; Sergeant, K. Changes in the proteome of Medicago sativa leaves in response to long-term cadmium exposure using a cell-wall targeted approach. Int. J. Mol. Sci. 2018, 19, 2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtaszek, P.; Stobiecki, M.; Bolwell, G.P. Changes in the composition of exocellular proteins of suspension-cultured Lupinus albus cells in response to fungal elicitors or CuCl2. J. Exp. Bot. 1997, 48, 2015–2021. [Google Scholar] [CrossRef] [Green Version]
- Sauer, D.B.; Burroughs, R. Disinfection of seed surfaces with sodium hypochlorite. Phytopathology 1986, 76, 745–749. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants Without Soil; Circular, California Agricultural Experiment Station; College of Agriculture, University of California: Berkeley, CA, USA, 1950; p. 347. [Google Scholar]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zhang, X. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. In Crop Physiology Research Methods; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef]
- Shinoda, K.; Tomita, M.; Ishihama, Y. emPAI Calc—For the estimation of protein abundance from large-scale identification data by liquid chromatography-tandem mass spectrometry. Bioinformatics 2010, 26, 576–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Protein Description | Gene Name | Coverage | Peptides | Fold Change | ||||
|---|---|---|---|---|---|---|---|---|---|
| T1/CK | T2/CK | T3/CK | T4/CK | T5/CK | |||||
| Energy and Carbohydrate Metabolism | |||||||||
| O82662 | Succinate—CoA ligase [ADP-forming] subunit beta | At2g20420 | 15% | 5 | 1.464 | 1.946 | 3.206 | 1.853 | 1.605 |
| P93338 | NADP-dependent glyceraldehyde-3-phosphate dehydrogenase | GAPN | 12% | 6 | 0.903 | 0.960 | 1.000 | 0.965 | 0.915 |
| P80261 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 3 | NAD9 | 21% | 3 | 1.098 | 1.401 | 1.634 | 1.193 | 1.342 |
| P57106 | Malate dehydrogenase 2 | MDH2 | 28% | 10 | 0.942 | 1.023 | 1.324 | 1.039 | 1.170 |
| P93819 | Malate dehydrogenase 1 | MDH1 | 37% | 11 | 0.985 | 1.044 | 1.379 | 1.097 | 1.179 |
| P16096 | Fructose-bisphosphate aldolase | N/A | 16% | 6 | 0.774 | 0.774 | 0.950 | 0.950 | 0.802 |
| O65581 | Fructose-bisphosphate aldolase 5 | FBA5 | 13% | 3 | 0.673 | 0.501 | 0.830 | 0.602 | 0.776 |
| P83483 | ATP synthase subunit beta-1 | At5g08670 | 48% | 24 | 1.212 | 1.271 | 1.295 | 1.323 | 1.034 |
| A4QL04 | ATP synthase subunit alpha | atpA | 29% | 15 | 1.164 | 0.874 | 1.026 | 1.561 | 1.078 |
| P12858 | Glyceraldehyde-3-phosphate dehydrogenase A | GAPA | 19% | 14 | 0.892 | 0.850 | 0.985 | 1.035 | 0.963 |
| O49485 | D-3-phosphoglycerate dehydrogenase 1 | PGDH1 | 14% | 7 | 0.628 | 0.734 | 1.004 | 1.090 | 0.744 |
| CO2 assimilation and photosynthesis | |||||||||
| P36491 | Photosystem II protein D1 | psbA | 16% | 6 | 0.692 | 0.744 | 1.233 | 0.904 | 0.793 |
| P69561 | Photosystem II protein D1 | psbA | 16% | 6 | 0.692 | 0.744 | 1.099 | 0.789 | 0.793 |
| A0A331 | Photosystem II CP43 reaction center protein | psbC | 21% | 8 | 1.481 | 1.535 | 2.054 | 1.855 | 0.642 |
| A0ZZ31 | Photosystem II CP43 reaction center protein | psbC | 18% | 6 | 0.627 | 0.733 | 1.004 | 0.910 | 0.744 |
| A4QJB4 | Photosystem I P700 chlorophyll a apoprotein A2 | psaB | 13% | 6 | 0.842 | 0.619 | 0.766 | 0.974 | 0.708 |
| A1EA38 | Cytochrome b6 | petB | 24% | 4 | 0.872 | 0.854 | 0.901 | 0.863 | 0.903 |
| Q01667 | Chlorophyll a-b binding protein 6 | LHCA1 | 19% | 9 | 0.991 | 0.945 | 1.102 | 0.950 | 0.998 |
| P27521 | Chlorophyll a-b binding protein 4 | LHCA4 | 17% | 5 | 0.904 | 0.862 | 1.004 | 1.048 | 0.910 |
| P27141 | Carbonic anhydrase | LOC107822687 | 13% | 4 | 0.657 | 0.783 | 1.004 | 0.860 | 0.738 |
| A1E9T2 | Ribulose bisphosphate carboxylase large chain | rbcL | 19% | 13 | 1.004 | 1.099 | 1.233 | 1.048 | 1.089 |
| P29409 | Phosphoglycerate kinase | N/A | 29% | 16 | 0.859 | 0.818 | 1.001 | 1.048 | 0.916 |
| P51062 | Phosphoenolpyruvate carboxylase | N/A | 8% | 7 | 1.211 | 1.110 | 0.775 | 0.731 | 0.673 |
| P23321 | Oxygen-evolving enhancer protein 1-1 | PSBO1 | 47% | 19 | 0.911 | 0.709 | 1.001 | 1.055 | 0.899 |
| Signal transduction and Protein Metabolism | |||||||||
| P34791 | Peptidyl-prolyl cis-trans isomerase CYP20-3 | CYP20-3 | 27% | 5 | 1.004 | 0.907 | 1.004 | 1.130 | 0.987 |
| P38547 | GTP-binding nuclear protein Ran2 | RAN2 | 18% | 4 | 0.972 | 1.474 | 1.476 | 1.536 | 1.560 |
| P38546 | GTP-binding nuclear protein Ran1 | RAN1 | 18% | 4 | 0.972 | 1.474 | 0.759 | 1.536 | 0.414 |
| P35480 | Chaperonin CPN60, mitochondrial | LOC106437789 | 18% | 10 | 1.786 | 1.823 | 2.514 | 1.699 | 1.614 |
| P36210 | 50S ribosomal protein L12-1 | RPL12A | 16% | 3 | 0.904 | 0.862 | 1.004 | 0.935 | 0.910 |
| O49203 | Nucleoside diphosphate kinase III | NDPK3 | 9% | 3 | 0.904 | 0.862 | 1.004 | 1.048 | 0.910 |
| Stress-and Defense-related proteins | |||||||||
| O78310 | Superoxide dismutase [Cu-Zn] 2 | CSD2 | 13% | 6 | 1.950 | 0.628 | 1.476 | 1.536 | 1.560 |
| P09678 | Superoxide dismutase [Cu-Zn] | SODCC | 18% | 3 | 0.875 | 1.475 | 1.255 | 1.048 | 0.910 |
| Q02028 | Stromal 70 kDa heat shock-related protein | HSP70 | 10% | 7 | 0.911 | 1.046 | 1.511 | 1.363 | 1.210 |
| Q05431 | L-ascorbate peroxidase 1 | APX1 | 14% | 4 | 1.096 | 1.753 | 1.186 | 2.735 | 1.038 |
| Sulfur assimilation and heavy metal detoxification | |||||||||
| P47999 | Cysteine synthase | OASB | 20% | 7 | 1.233 | 1.240 | 1.002 | 1.148 | 0.970 |
| Q8L7C9 | Glutathione S-transferase U20 | GSTU20 | 9.70% | 3 | 1.904 | 1.321 | 1.643 | 1.048 | 1.400 |
| Q42624 | Glutamine synthetase | GLN2 | 25% | 14 | 0.941 | 0.952 | 1.008 | 1.252 | 1.176 |
| O50008 | 5-methyltetrahydropteroyltriglutamate—homocysteine methyltransferase 1 | MS1 | 29% | 26 | 1.046 | 0.991 | 1.370 | 1.373 | 1.074 |
| Cytoskeleton and cell wall-related proteins | |||||||||
| P49236 | Glucan endo-1,3-beta-glucosidase | BGL | 8% | 3 | 1.111 | 0.821 | 3.215 | 9.134 | 1.093 |
| P45854 | Germin-like protein 1 | GLP1 | 25% | 5 | 1.093 | 1.042 | 1.215 | 1.268 | 1.101 |
| O22349 | Tubulin alpha-3 chain | TUBA3 | 14% | 8 | 0.952 | 0.884 | 0.794 | 0.862 | 1.004 |
| Development | |||||||||
| P51823 | ADP-ribosylation factor 2 | ARF | 46% | 7 | 0.404 | 0.379 | 1.273 | 0.085 | 1.302 |
| P51821 | ADP-ribosylation factor 1 | ARF1 | 46% | 7 | 0.239 | 0.240 | 0.929 | 0.052 | 0.235 |
| P49076 | ADP-ribosylation factor | ARF1 | 46% | 7 | 0.703 | 0.463 | 0.109 | 0.109 | 0.419 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, Y.-H.; Roy, S.K.; Roy Choudhury, A.; Kwon, S.-J.; Choi, J.-Y.; Rahman, M.A.; Katsube-Tanaka, T.; Shiraiwa, T.; Lee, M.-S.; Cho, K.; et al. Proteome Changes Reveal the Protective Roles of Exogenous Citric Acid in Alleviating Cu Toxicity in Brassica napus L. Int. J. Mol. Sci. 2021, 22, 5879. https://doi.org/10.3390/ijms22115879
Ju Y-H, Roy SK, Roy Choudhury A, Kwon S-J, Choi J-Y, Rahman MA, Katsube-Tanaka T, Shiraiwa T, Lee M-S, Cho K, et al. Proteome Changes Reveal the Protective Roles of Exogenous Citric Acid in Alleviating Cu Toxicity in Brassica napus L. International Journal of Molecular Sciences. 2021; 22(11):5879. https://doi.org/10.3390/ijms22115879
Chicago/Turabian StyleJu, Young-Hwan, Swapan Kumar Roy, Aritra Roy Choudhury, Soo-Jeong Kwon, Ju-Young Choi, Md Atikur Rahman, Tomoyuki Katsube-Tanaka, Tatsuhiko Shiraiwa, Moon-Soon Lee, Kun Cho, and et al. 2021. "Proteome Changes Reveal the Protective Roles of Exogenous Citric Acid in Alleviating Cu Toxicity in Brassica napus L." International Journal of Molecular Sciences 22, no. 11: 5879. https://doi.org/10.3390/ijms22115879
APA StyleJu, Y. -H., Roy, S. K., Roy Choudhury, A., Kwon, S. -J., Choi, J. -Y., Rahman, M. A., Katsube-Tanaka, T., Shiraiwa, T., Lee, M. -S., Cho, K., & Woo, S. -H. (2021). Proteome Changes Reveal the Protective Roles of Exogenous Citric Acid in Alleviating Cu Toxicity in Brassica napus L. International Journal of Molecular Sciences, 22(11), 5879. https://doi.org/10.3390/ijms22115879