
The Role of NLRP1, NLRP3, and AIM2 Inflammasomes in Psoriasis: Review

 ,
, 
Abstract
:1. Introduction
2. The NLRP3 Inflammasome (in Psoriasis)
3. The NLRP1 Inflammasome (in Psoriasis)
4. The AIM2 Inflammasome (in Psoriasis)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langley, R.G.; Krueger, G.G.; Griffiths, C.E. Psoriasis: Epidemiology, clinical features, and quality of life. Ann. Rheum. Dis. 2005, 64 (Suppl. 2), 18–23. [Google Scholar] [CrossRef] [Green Version]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef]
- Alinaghi, F.; Calov, M.; Kristensen, L.E.; Gladman, D.D.; Coates, L.C.; Jullien, D.; Gottlieb, A.B.; Gisondi, P.; Wu, J.J.; Thyssen, J.P.; et al. Prevalence of psoriatic arthritis in patients with psoriasis: A systematic review and meta-analysis of observational and clinical studies. J. Am. Acad. Dermatol. 2019, 80, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira Mde, F.; Rocha, O.; Duarte, G.V. Psoriasis: Classical and emerging comorbidities. Bras. Dermatol. 2015, 90, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Benhadou, F.; Mintoff, D.; del Marmol, V. Psoriasis: Keratinocytes or Immune Cells–Which Is the Trigger? Dermatology 2019, 235, 91–100. [Google Scholar] [CrossRef]
- McKee, P.H.; Calonje, E.; Granter, S.R. Pathology of the Skin: With Clinical Correlations, 3rd ed.; Mosby Ltd.: Maryland Heights, MO, USA, 2011. [Google Scholar]
- Wagner, E.F.; Schonthaler, H.B.; Guinea–Viniegra, J.; Tschachler, E. Psoriasis: What we have learned from mouse models. Nat. Rev. Rheumatol. 2010, 6, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yang, R.; Wen, C.; Li, H.; Zhao, H. Expression of NLRP3 inflammasome in BALB/c mice with imiquimod-induced psoriasis-like inflammation and therapeutic effect of mustard seed (Sinapis Alba Linn). J. South. Med. Univ. 2013, 33, 1394–1398. [Google Scholar]
- Jiang, W.; Zhu, F.G.; Bhagat, L.; Yu, D.; Tang, J.X.; Kandimalla, E.R.; La Monica, N.; Agrawal, S. A Toll-like receptor 7, 8, and 9 antagonist inhibits Th1 and Th17 responses and inflammasome activation in a model of IL-23-induced psoriasis. J. Investig. Dermatol. 2013, 133, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Goblos, A.; Danis, J.; Vas, K.; Bata-Csorgo, Z.; Kemeny, L.; Szell, M. Keratinocytes express functional CARD18, a negative regulator of inflammasome activation, and its altered expression in psoriasis may contribute to disease pathogenesis. Mol. Immunol. 2016, 73, 10–18. [Google Scholar] [CrossRef]
- Vaccari, J.P.; Dietrich, W.D.; Keane, R.W. Activation and regulation of cellular inflammasomes: Gaps in our knowledge for central nervous system injury. J. Cereb. Blood Flow Metab. 2014, 34, 369–375. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Ciążyńska, M.; Bednarski, I.A.; Wódz, K.; Narbutt, J.; Lesiak, A. NLRP1 and NLRP3 inflammasomes as a new approach to skin carcinogenesis (Review). Oncol. Lett. 2020, 19, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Sedimbi, S.K.; Hagglof, T.; Karlsson, M.C. IL-18 in inflammatory andautoimmune disease. Cell Mol. Life Sci. 2013, 70, 4795–4808. [Google Scholar] [CrossRef] [PubMed]
- Ekman, A.K.; Verma, D.; Fredrikson, M.; Bivik, C.; Enerbäck, C. Genetic variations of NLRP1: Susceptibility in psoriasis. Br. J. Dermatol. 2014, 171, 1517–1520, Epub on 2014 October 22. [Google Scholar] [CrossRef] [PubMed]
- Debets, R.; Hegmans, J.P.; Troost, R.J.; Benner, R.; Prens, E.P. Enhanced production of biologically active interleukin-1 alpha and interleukin-1 beta by psoriatic epidermal cells ex vivo: Evidence of increased cytosolic interleukin-1 beta levels and facilitated interleukin-1 release. Eur. J. Immunol. 1995, 25, 1624–1630. [Google Scholar] [CrossRef] [Green Version]
- Rasmy, H.; Mikhael, N.; Ismail, S. Interleukin-18 expression and the response to treatment in patients with psoriasis. Arch. Med. Sci. 2011, 7, 713–719. [Google Scholar] [CrossRef]
- Carlstrom, M.; Ekman, A.K.; Petersson, S.; Soderkvist, P.; Enerback, C. Genetic support for the role of the NLRP3 inflammasome in psoriasis susceptibility. Exp. Derm. 2012, 21, 932–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dombrowski, Y.; Peric, M.; Koglin, S.; Kammerbauer, C.; Göss, C.; Anz, D.; Simanski, M.; Gläser, R.; Harder, J.; Hornung, V.; et al. Cytosolic DNA triggers inflammasome activation in keratinocytes in psoriatic lesions. Sci. Transl. Med. 2011, 3, 82ra38. [Google Scholar] [CrossRef] [Green Version]
- Deng, G.; Chen, W.; Wang, P.; Zhan, T.; Zheng, W.; Gu, Z.; Wang, X.; Ji, X.; Sun, Y. Inhibition of NLRP3 inflammasome-mediated pyroptosis in macrophage by cycloastragenol contributes to amelioration of imiquimod-induced psoriasis-like skin inflammation in mice. Int. Immunopharmacol. 2019, 74, 105682. [Google Scholar] [CrossRef]
- Su, F.; Xia, Y.; Huang, M.; Zhang, L.; Chen, L. Expression of NLPR3 in Psoriasis Is Associated with Enhancement of Interleukin-1β and Caspase-1. Med. Sci. Monit. 2018, 24, 7909–7913. [Google Scholar] [CrossRef]
- Yun, M.; Seo, G.; Lee, J.Y.; Chae, G.T.; Lee, S.B. Epigallocatechin−3–gallate attenuates the AIM2–induced secretion of IL−1beta in human epidermal keratinocytes. Biochem. Biophys. Res. Commun. 2015, 467, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.C.; Yuan, S.N.; OuYang, C.N.; Hu, S.I.; Lin, H.C.; Huang, K.Y.; Lin, W.-N.; Chuang, Y.-T.; Chen, Y.-J.; Ojcius, D.M.; et al. EFLA 945 restricts AIM2 inflammasome activation by preventing DNA entry for psoriasis treatment. Cytokine 2020, 127, 154951. [Google Scholar] [CrossRef] [PubMed]
- Walter, A.; Schafer, M.; Cecconi, V.; Matter, C.; Urosevic–Maiwald, M.; Belloni, B.; Schönewolf, N.; Dummer, R.; Bloch, W.; Werner, S.; et al. Aldara activates TLR7–independent immune defence. Nat. Commun. 2013, 4, 1560. [Google Scholar] [CrossRef]
- Zhang, C.; Xiao, C.; Dang, E.; Cao, J.; Zhu, Z.; Fu, M.; Yao, X.; Liu, Y.; Jin, B.; Wang, G.; et al. CD100-plexin-B2 promotes the inflammation in psoriasis by activating NF-kappaB and the inflammasome in keratinocytes. J. Investig. Dermatol. 2018, 138, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Zeng, J.; Li, W.; Lin, L.; Zhou, X.; Tian, X.; Liu, W.; Zhang, L.; Zhang, X. Silencing of miR155 suppresses inflammatory responses in psoriasis through inflammasome NLRP3 regulation. Int. J. Mol. Med. 2018, 42, 1086–1095. [Google Scholar] [PubMed] [Green Version]
- Irrera, N.; Vaccaro, M.; Bitto, A.; Pallio, G.; Pizzino, G.; Lentini, M.; Arcoraci, V.; Minutoli, L.; Scuruchi, M.; Cutroneo, G.; et al. BAY 11–7082 inhibits the NF–kappaB and NLRP3 inflammasome pathways and protects against IMQ–induced psoriasis. Clin. Sci. 2017, 131, 487–498. [Google Scholar] [CrossRef]
- Rabeony, H.; Pohin, M.; Vasseur, P.; Petit-Paris, I.; Jégou, J.; Favot, L.; Frouin, E.; Boutet, M.; Blanchard, F.; Togbe, D.; et al. IMQ-induced skin inflammation in mice is dependent on IL-1R1 and MyD88 signaling but independent of the NLRP3 inflammasome. Eur. J. Immunol. 2015, 45, 2847–2857. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Perez, J.A.; Killeen, M.E.; Yang, Y.; Carey, C.D.; Falo, L.D.; Mathers, A.R. Extracellular ATP and IL-23 form a local inflammatory circuit leading to the development of a neutrophildependent psoriasiform dermatitis. J. Investig. Dermatol. 2018, 138, 2595–2605. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Yang, B.Y.; Cheng, Y.G.; Liu, Y.; Liu, Y.; Tan, J.Y.; Guan, W.; Guo, S.; Kuang, H.-X.; Liu, Y. Datura Metel L. Ameliorates imiquimod–induced psoriasis–like dermatitis and inhibits inflammatory cytokines production through TLR7/8–MyD88–NF– kappaB–NLRP3 inflammasome pathway. Molecules 2019, 24, 2157. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Duncan, B.; Li, X.; Shi, J. The role of NLRP3 inflammasome in infection-related, immune-mediated and autoimmune skin diseases. J. Dermatol. Sci. 2020, 98, 146–151, Epub on 2020 March 6. [Google Scholar] [CrossRef] [PubMed]
- Primiano, M.J.; Lefker, B.A.; Bowman, M.R.; Bree, A.G.; Hubeau, C.; Bonin, P.D.; Mangan, M.; Dower, K.; Monks, B.G.; Cushing, L.; et al. Efficacy and pharmacology of the NLRP3 inflammasome inhibitor CP-456,773 (CRID3) in murine models of dermal and pulmonary inflammation. J. Immunol. 2016, 197, 2421–2433. [Google Scholar] [CrossRef] [Green Version]
- Forouzandeh, M.; Besen, J.; Keane, R.W.; Vaccari, J.P. The Inflammasome Signaling Proteins ASC and IL-18 as Biomarkers of Psoriasis. Front. Pharmacol. 2020, 11, 1238, PMCID: PMC7438850. [Google Scholar] [CrossRef] [PubMed]
- Day, T.G.; Ramanan, A.V.; Hinks, A.; Lamb, R.; Packham, J.; Wise, C.; Punaro, M.; Donn, R.P. Autoinflammatory genes and susceptibility to psoriatic juvenile idiopathic arthritis. Arthritis Rheum. 2008, 58, 2142–2146. [Google Scholar] [CrossRef] [PubMed]
- Kastbom, A.; Verma, D.; Eriksson, P.; Skogh, T.; Wingren, G.; Söderkvist, P. Genetic variation in proteins of the cryopyrin inflammasome influences susceptibility and severity of rheumatoid arthritis (The Swedish TIRA project). Rheumatology 2008, 47, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Hao, S.; Zheng, H.; Zhao, X.; Li, Y. Association of NLRP1 and NLRP3 Polymorphisms with Psoriasis Vulgaris Risk in the Chinese Han Population. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Burian, M.; Yazdi, A.S. NLRP1 Is the Key Inflammasome in Primary Human Keratinocytes. J. Investig. Dermatol. 2018, 138, 2507–2510. [Google Scholar] [CrossRef] [Green Version]
- Fenini, G.; Karakaya, T.; Hennig, P.; Di Filippo, M.; Beer, H.-D. The NLRP1 Inflammasome in Human Skin and Beyond. Int. J. Mol. Sci. 2020, 21, 4788. [Google Scholar] [CrossRef]
- Fenini, G.; Grossi, S.; Contassot, E.; Biedermann, T.; Reichmann, E.; French, L.E.; Beer, H.-D. Genome editing of human primary keratinocytes by CRISPR/Cas9 reveals an essential role of the NLRP1 inflammasome in UVB sensing. J. Investig. Dermatol. 2018, 138, 2644–2652. [Google Scholar] [CrossRef] [Green Version]
- Juneblad, K.; Kastbom, A.; Johansson, L.; Rantapää-Dahlqvist, S.; Söderkvist, P.; Alenius, G.M. Association between inflammasome-related polymorphisms and psoriatic arthritis. Scand. J. Rheumatol. 2021, 50, 206–212. [Google Scholar] [CrossRef]
- Sharma, B.R.; Karki, R.; Kanneganti, T.D. Role of AIM2 inflammasome in inflammatory diseases, cancer and infection. Eur. J. Immunol. 2019, 49, 1998–2011. [Google Scholar] [CrossRef] [Green Version]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Peric, M.; Koglin, S.; Dombrowski, Y.; Gross, K.; Bradac, E.; Büchau, A.; Steinmeyer, A.; Zügel, U.; Ruzicka, T.; Schauber, J. Vitamin D analogs differentially control antimicrobial peptide/“alarmin” expression in psoriasis. PLoS ONE 2009, 4, e6340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vähävihu, K.; Ala-Houhala, M.; Peric, M.; Karisola, P.; Kautiainen, H.; Hasan, T.; Snellman, E.; Alenius, H.; Schauber, J.; Reunala, T. Narrowband ultraviolet B treatment improves vitamin D balance and alters antimicrobial peptide expression in skin lesions of psoriasis and atopic dermatitis. Br. J. Dermatol. 2010, 163, 321–328. [Google Scholar] [CrossRef]
- Feldmeyer, L.; Werner, S.; French, L.E.; Beer, H.D. Interleukin−1, inflammasomes and the skin. Eur. J. Cell Biol. 2010, 89, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, Y.; Schauber, J. Cathelicidin LL−37, a defense molecule with a potential role in psoriasis pathogenesis. Exp. Dermatol. 2012, 21, 327–330. [Google Scholar] [CrossRef]
- Chan, J.R.; Blumenschein, W.; Murphy, E.; Diveu, C.; Wiekowski, M.; Abbondanzo, S.; Lucian, L.; Geissler, R.; Brodie, S.; Kimball, A.B.; et al. IL-23 stimulates epidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasis pathogenesis. J. Exp. Med. 2006, 203, 2577–2587. [Google Scholar] [CrossRef]
- Zheng, Y.; Danilenko, D.M.; Valdez, P.; Kasman, I.; Eastham-Anderson, J.; Wu, J.; Ouyang, W. Interleukin-22, a T(H)17 cytokine, mediates IL-23-induced dermal inflammation and acanthosis. Nature 2007, 445, 648–651. [Google Scholar] [CrossRef]
- Hedrick, M.N.; Lonsdorf, A.S.; Shirakawa, A.K.; Richard Lee, C.C.; Liao, F.; Singh, S.P.; Zhang, H.H.; Grinberg, A.; Love, P.E.; Hwang, S.T.; et al. CCR6 is required for IL-23-induced psoriasis-like inflammation in mice. J. Clin. Investig. 2009, 119, 2317–2329. [Google Scholar] [CrossRef] [Green Version]



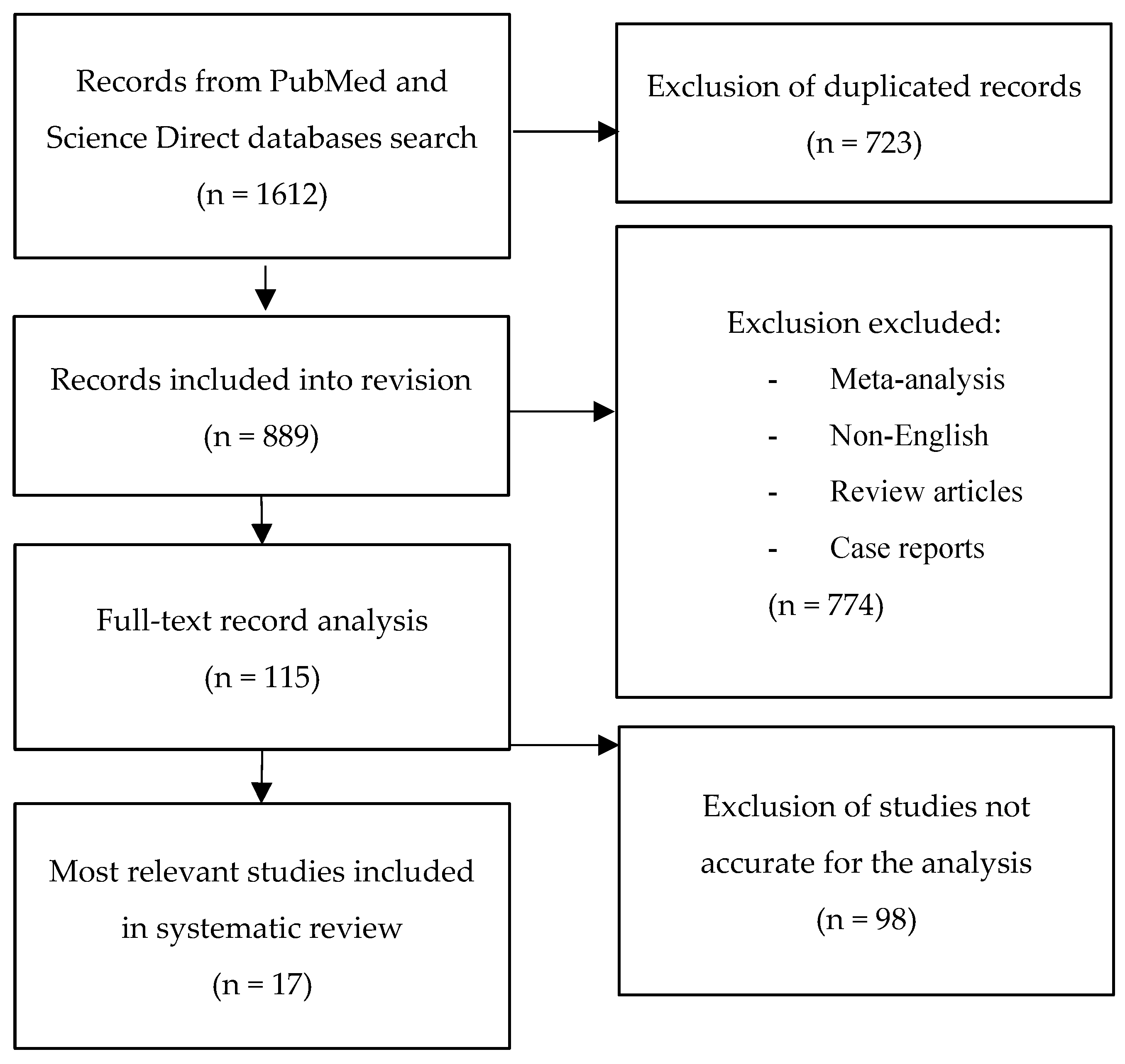{kind=link}
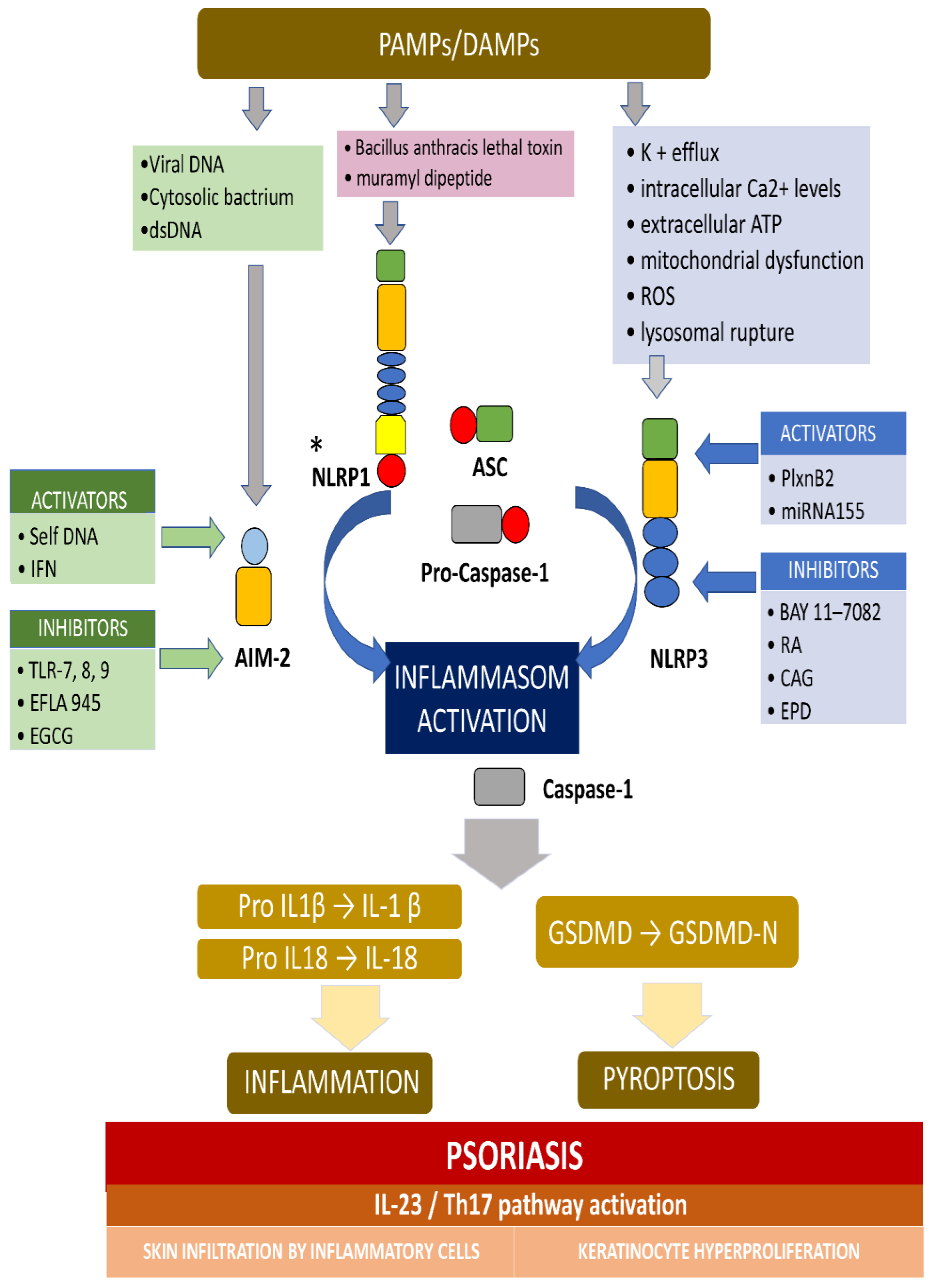{kind=link}
| Author | Inflammasome | Effector Signal | Cell Type | Mouse/Human | Main Finding |
|---|---|---|---|---|---|
| Dombrowski et al. [20] | AIM2 | IL-1β | Keratinocytes | Human | Cytosolic DNA can trigger AIM2 inflammasome IL-1β activation in psoriasis; cathelicidin LL-37 interfered with DNA-sensing inflammasomes. |
| Ekman et al. [16] | NLRP1 | IL-1β, IL-18 | n/a | Human | NLRP1 inflammasome complex genetic variations are related to elevated vulnerability to psoriasis. |
| Deng et al. [21] | NLRP3 | IL-1β | Dendritic cells, Neutrophils, T lymphocytes | Mouse | CAG significantly reduced imiquimod-triggered NLRP3 inflammasome activation and gasdermin D (GSDMD)-mediated pyroptosis. |
| Su et al. [22] | NLRP3 | IL-1β, caspase 1 | Keratinocytes | Human | The expression of NLRP3 in psoriatic samples was 3.5 to 4.3 times higher than the expression of NLRP3 in normal skin biopsy samples; NLRP3 expression is upregulated in psoriasis and is associated with concomitant accumulation of IL-1b and caspase 1. |
| Yun et al. [23] | AIM2 | IL-1β | HEKn | Human | EGCG reduces AIM2-induced IL-1b secretion by suppressing IL 1b-mediated priming and poly-induced ASC oligomerization. |
| Chung et al. [24] | AIM2 | IL-1β, IL-18 | Macrophages | Human | EFLA 945 attenuates IMQ-induced psoriasis-related proinflammatory responses; |
| Jiang et al. [9] | AIM2 | IL-1β, IL-18 | n/a | Mouse | the inflammasome expression components NLRP3 and AIM2 are reduced by antagonist treatment. |
| Walter et al. [25] | NLRP1 | IL-1β, IL-18 | Neutrophils, Keratinocytes | Mouse | NLRP1 inflammasome activation is promoted by isostearic acid in cultured keratinocytes. |
| Zhang et al. [26] | NLRP3 | IL-1β | Keratinocytes | Human | NLRP3 inflammasome is activated by CD100 in keratinocytes through binding to PLXNB2. |
| Luo et al. [27] | NLRP3 | IL-1β, IL-18 | Keratinocytes | Mouse | miR-155 activates the NLRP3 inflammasome; miR-155 does not impact the TLR4/NF-kB signaling pathway. |
| Yang et al. [26] | NLRP3 | IL-1β | Keratinocytes | Mouse | EPD inhibits the production of imiquimod-induced inflammatory cytokines |
| Irrera et al. [28] | NLRP3 | IL-1β, IL-18 | Keratinocytes | Mouse | BAY 11-7082 alleviates the NLRP3 and dual NF-kB. |
| Rabeony et al. [29] | NLRP3 | n/a | n/a | Mouse | IMQ-induced skin inflammation is independent on the NLRP3 inflammasome. |
| Diaz–Perez et al. [30] | NLRP3 | IL-1β | Macrophages, Granulocytes, Neutrophils | Mouse | P2 × 7R-induced inflammation is largely dependent on the IL-1b/NLRP3 inflammasome pathway and neutrophils. |
| Shi et al. [31] | NLRP3 | IL-1β | Keratinocytes | Mouse | Caspase-11 and caspase-1 can cleave GSDMD, revealing a pyroptosis-inducing fragment |
| Yang et al. [32] | NLRP3 | IL-1β | Keratinocytes | Mouse | EPD exhibited a protective effect on an imiquimod-induced psoriasis mice model by inhibiting the inflammatory response, which might be ascribed to the inhibition of the TLR7/8–MyD88–NF-κB–NLRP3 inflammasome pathway |
| Forouzandeh et al. [32] | NLRP3 | ASC, IL-1β, IL-18 | Keratinocytes | Human | ASC and IL-18 play a significant role in the inflammatory response associated with the pathology of psoriasis. These inflammasome proteins appear to be key biomarkers in determining diagnoses in this patient population. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciążyńska, M.; Olejniczak-Staruch, I.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibińska, M.; Lesiak, A. The Role of NLRP1, NLRP3, and AIM2 Inflammasomes in Psoriasis: Review. Int. J. Mol. Sci. 2021, 22, 5898. https://doi.org/10.3390/ijms22115898
Ciążyńska M, Olejniczak-Staruch I, Sobolewska-Sztychny D, Narbutt J, Skibińska M, Lesiak A. The Role of NLRP1, NLRP3, and AIM2 Inflammasomes in Psoriasis: Review. International Journal of Molecular Sciences. 2021; 22(11):5898. https://doi.org/10.3390/ijms22115898
Chicago/Turabian StyleCiążyńska, Magdalena, Irmina Olejniczak-Staruch, Dorota Sobolewska-Sztychny, Joanna Narbutt, Małgorzata Skibińska, and Aleksandra Lesiak. 2021. "The Role of NLRP1, NLRP3, and AIM2 Inflammasomes in Psoriasis: Review" International Journal of Molecular Sciences 22, no. 11: 5898. https://doi.org/10.3390/ijms22115898
APA StyleCiążyńska, M., Olejniczak-Staruch, I., Sobolewska-Sztychny, D., Narbutt, J., Skibińska, M., & Lesiak, A. (2021). The Role of NLRP1, NLRP3, and AIM2 Inflammasomes in Psoriasis: Review. International Journal of Molecular Sciences, 22(11), 5898. https://doi.org/10.3390/ijms22115898