
Changes in Phenotypes and DNA Methylation of In Vitro Aging Sperm in Common Carp Cyprinus carpio

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
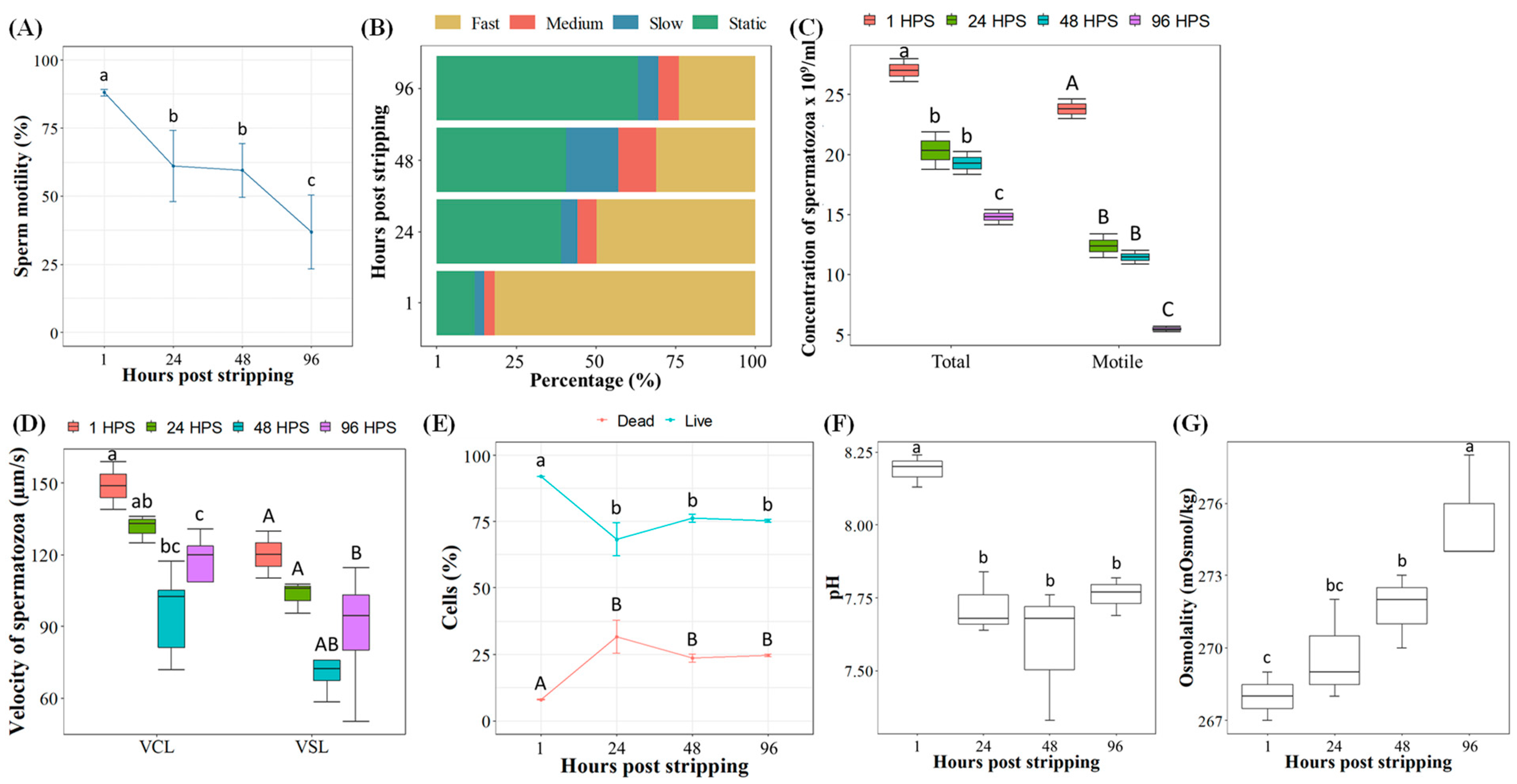
2.1. Effects of Pooled Sperm In Vitro Aging on Different Phenotypic Characteristics
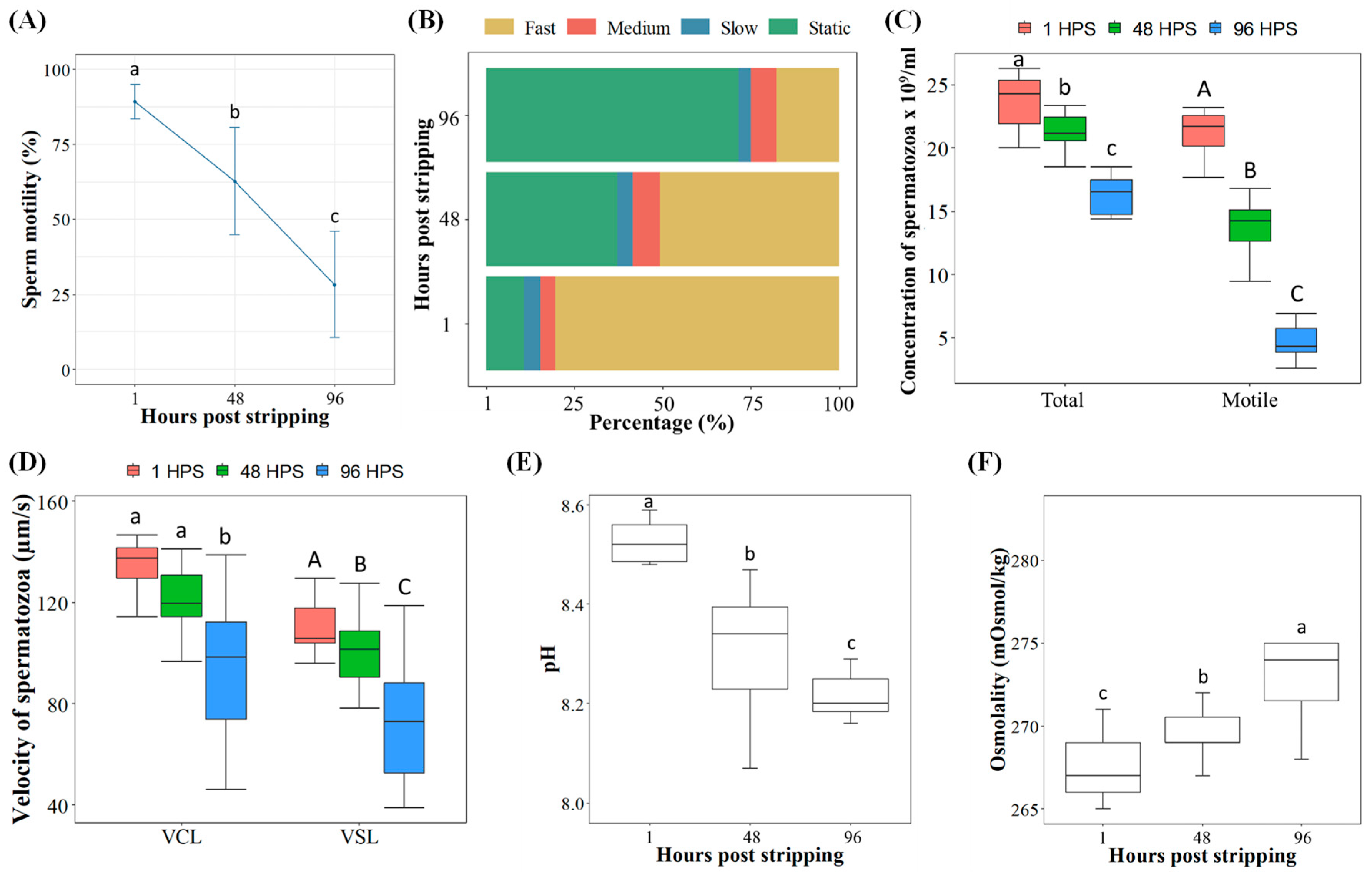
2.2. Effects of Sperm In Vitro Aging from Individual Males on Phenotypic Characteristics
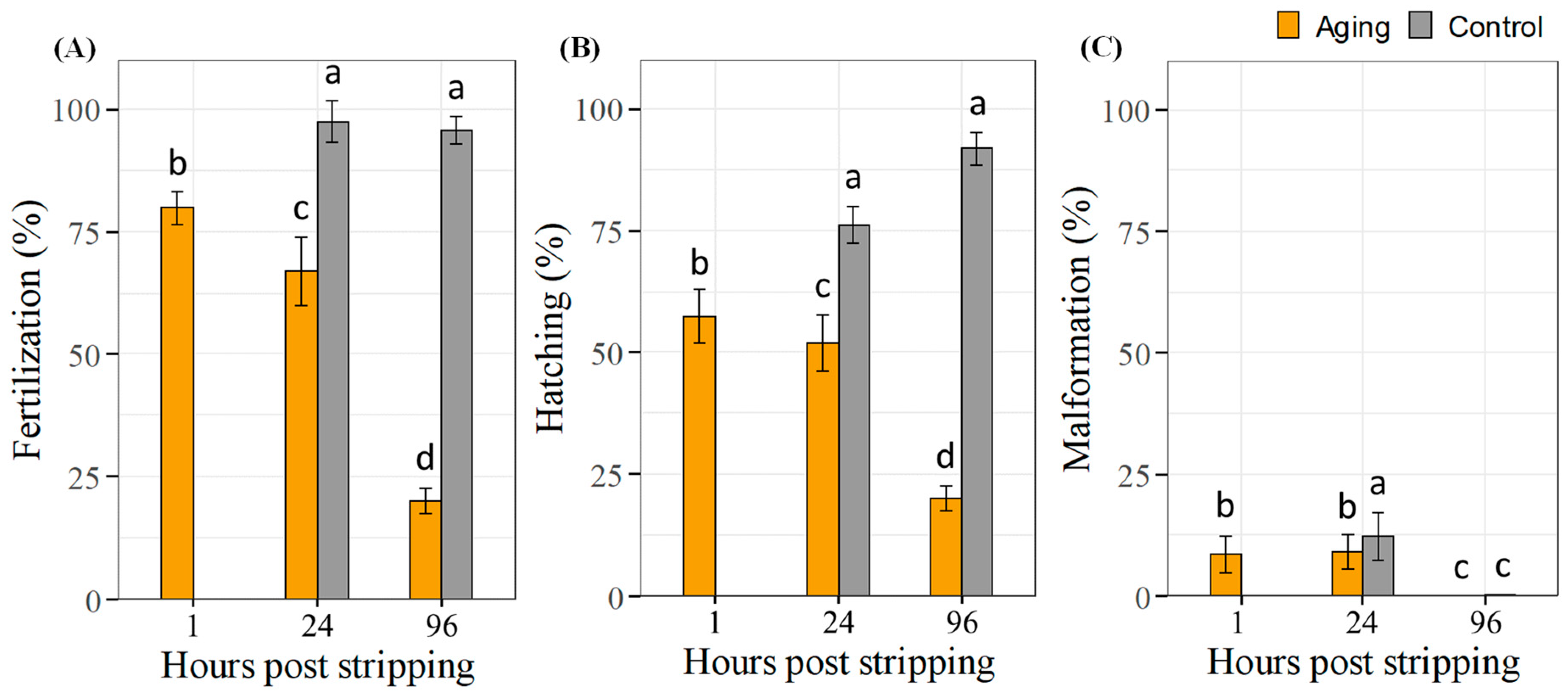
2.3. Effects of Pooled Sperm In Vitro Aging on Fertilization, Hatching and Larval Malformation Rates
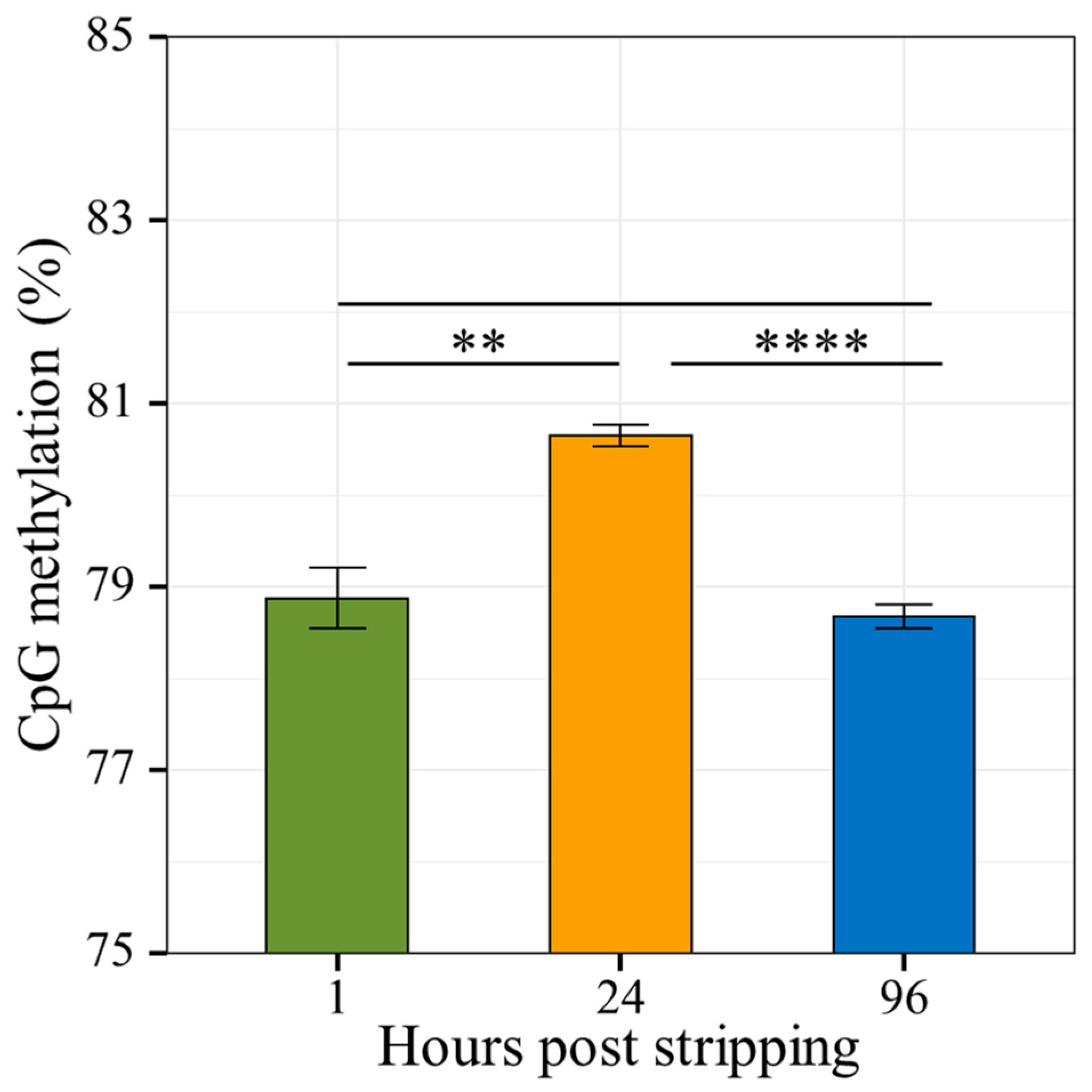
2.4. Bisulfite Sequencing of Carp Spermatozoa Genomic DNA during Aging
2.4.1. Differential Methylation Analysis
2.4.2. Functional Annotation of Differentially Methylated Loci
3. Discussion
4. Material and Methods
4.1. Ethics Statement and Animals
4.2. Experimental Fish
4.3. Experimental Design
4.3.1. Phenotypic Parameters of Pooled Sperm during In Vitro Aging
4.3.2. Phenotypic Parameters of Sperm from Individual Males during In Vitro Aging
4.3.3. Fertilization and Hatching Level with Malformation Embryos Rates from Pooled In Vitro Aged Sperm
4.3.4. Whole Genome Bisulfite Sequencing from Pooled In Vitro Aged Sperm
4.4. Examining Phenotypic Characteristics of Spermatozoa during In Vitro Aging
4.4.1. Sperm Motility, Velocity and Concentration
4.4.2. Osmolality and pH of Seminal Plasma
4.4.3. Sperm Viability Analysis
4.5. Examining Fertilization, Hatching and Larval Malformation Rates during Spermatozoa In Vitro Aging
4.6. Whole Genome Bisulfite Sequencing of Sperm
4.6.1. DNA Isolation from Carp Sperm Samples
4.6.2. Functional Annotation of Differentially Methylated Regions
4.7. Data Evaluation and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HPS | Hours post stripping |
| PSA | Post sperm activation |
| CpG | Cytosine phosphate guanine dinucleotide |
| KIS | Kurokura immobilizing solution |
| PI | Propidium iodide |
| DMR | Differentially methylated region |
| MBD-seq | Methyl-CpG binding domain sequencing |
| DNMT | DNA methyltransferase |
| RAS | Recirculation Aquaculture System |
| VCL | Curvilinear velocity |
| VSL | Straight-line velocity |
| WGBS | whole-genome bisulfite sequencing |
References
- Babiak, I.; Ottesen, O.; Rudolfsen, G.; Johnsen, S. Quantitative characteristics of Atlantic halibut, Hippoglossus hippoglossus L., semen throughout the reproductive season. Theriogenology 2006, 65, 1587–1604. [Google Scholar] [CrossRef]
- Risopatrón, J.; Merino, O.; Cheuquemán, C.; Figueroa, E.; Sánchez, R.; Farías, J.G.; Valdebenito, I. Effect of the age of broodstock males on sperm function during cold storage in the trout (Oncorhynchus mykiss). Andrologia 2018, 50, e12857. [Google Scholar] [CrossRef]
- Gasparini, C.; Dosselli, R.; Evans, J.P. Sperm storage by males causes changes in sperm phenotype and influences the reproductive fitness of males and their sons. Evol. Lett. 2017, 1, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajitschek, S.; Hotzy, C.; Zajitschek, F.; Immler, S. Short-Term variation in sperm competition causes sperm-mediated epigenetic effects on early offspring performance in the zebrafish. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140422. [Google Scholar] [CrossRef] [Green Version]
- Shaliutina, A.; Hulak, M.; Dzuyba, B.; Linhart, O. Spermatozoa motility and variation in the seminal plasma proteome of Eurasian perch (Perca fluviatilis) during the reproductive season. Mol. Reprod. Dev. 2012, 79, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Shaliutina, A.; Hulak, M.; Gazo, I.; Linhartova, P.; Linhart, O. Effect of short-term storage on quality parameters, DNA integrity, and oxidative stress in Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) sturgeon sperm. Anim. Reprod. Sci. 2013, 139, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Trigo, P.; Merino, O.; Figueroa, E.; Valdebenito, I.; Sánchez, R.; Risopatrón, J. Effect of short-term semen storage in salmon (Oncorhynchus mykiss) on sperm functional parameters evaluated by flow cytometry. Andrologia 2015, 47, 407–411. [Google Scholar] [CrossRef]
- Contreras, P.; Ulloa, P.; Merino, O.P.; Valdebenito, I.; Figueroa, E.; Farías, J.G.; Risopatrón, J. Effect of short-term storage on sperm function in Patagonian blenny (Eleginops maclovinus) sperm. Aquaculture 2017, 481, 58–63. [Google Scholar] [CrossRef]
- Xin, M.M.; Sterba, J.; Shaliutina-Kolesova, A.; Dzyuba, B.; Lieskovska, J.; Boryshpolets, S.; Siddique, M.A.M.; Kholodnyy, V.; Lebeda, I.; Linhart, O. Protective role of antifreeze proteins on sterlet (Acipenser ruthenus) sperm during cryopreservation. Fish Physiol. Biochem. 2018, 44, 1527–1533. [Google Scholar] [CrossRef]
- Xin, M.M.; Cheng, Y.; Rodina, M.; Tučková, V.; Shelton, W.L.; Linhart, O. Improving motility and fertilization capacity of low-quality sperm of sterlet Acipenser ruthenus during storage. Theriogenology 2020, 156, 90–96. [Google Scholar] [CrossRef]
- Harman, D. The free radical theory of aging. Antioxid. Redox Signal. 2003, 5, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Sabanegh, E.; Kim, T.; Agarwal, A. Free radical theory of aging: Implications in male infertility. Urology 2010, 75, 14–19. [Google Scholar] [CrossRef]
- Samarin, A.M.; Ostbye, T.K.K.; Ruyter, B.; Sampels, S.; Burkina, V.; Blecha, M.; Gela, D.; Policar, T. Alteration of mRNA abundance, oxidation products and antioxidant enzyme activities during oocyte ageing in common carp Cyprinus carpio. PLoS ONE 2019, 14, e0212694. [Google Scholar] [CrossRef]
- Crary-Dooley, F.K.; Tam, M.E.; Dunaway, K.W.; Hertz-Picciotto, I.; Schmidt, R.J.; LaSalle, J.M. A comparison of existing global DNA methylation assays to low-coverage whole-genome bisulfite sequencing for epidemiological studies. Epigenetics 2017, 12, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.F.; Emperle, M.; Guo, Y.R.; Grimm, S.; Ren, W.D.; Adam, S.; Uryu, H.; Zhang, Z.M.; Chen, D.L.; Yin, J.K.; et al. Comprehensive structure-function characterization of DNMT3B and DNMT3A reveals distinctive de novo DNA methylation mechanisms. Nat. Commun. 2020, 11, 3355. [Google Scholar] [CrossRef]
- Woods, L.C.; Li, Y.K.; Ding, Y.; Liu, J.N.; Reading, B.J.; Fuller, S.A.; Song, J.Z. DNA methylation profiles correlated to striped bass sperm fertility. BMC Genom. 2018, 19, 244. [Google Scholar] [CrossRef] [Green Version]
- Montjean, D.; Zini, A.; Ravel, C.; Belloc, S.; Dalleac, A.; Copin, H.; Boyer, P.; Mcelreavey, K.; Benkhalifa, M. Sperm global DNA methylation level: Association with semen parameters and genome integrity. Andrology 2015, 3, 235–240. [Google Scholar] [CrossRef]
- Zhao, W.L.; Gu, N.H.; Li, Z.Z.; Wang, G.S.; Cheng, C.Y.; Sun, F. Autism-Like behaviors and abnormality of glucose metabolism in offspring derived from aging males with epigenetically modified sperm. Aging 2020, 12, 19766–19784. [Google Scholar] [CrossRef]
- Huypens, P.; Sass, S.; Wu, M.; Dyckhoff, D.; Tschöp, M.; Theis, F.; Marschall, S.; De Angelis, M.H.; Beckers, J. Epigenetic germline inheritance of diet-induced obesity and insulin resistance. Obstet. Gynecol. Surv. 2016, 71, 719–720. [Google Scholar] [CrossRef]
- Watkins, A.J.; Dias, I.; Tsuro, H.; Allen, D.; Emes, R.D.; Moreton, J.; Wilson, R.; Ingram, R.J.M.; Sinclair, K.D. Paternal diet programs offspring health through sperm- and seminal plasma-specific pathways in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10064–10069. [Google Scholar] [CrossRef] [Green Version]
- Potabattula, R.; Dittrich, M.; Schorsch, M.; Hahn, T.; Haaf, T.; EI Hajj, N. Male obesity effects on sperm and next-generation cord blood DNA methylation. PLoS ONE 2019, 14, e0218615. [Google Scholar] [CrossRef] [PubMed]
- Kläver, R.; Bleiziffer, A.; Redmann, K.; Mallidis, C.; Kliesch, S.; Gromoll, J. Routine cryopreservation of spermatozoa is safe-Evidence from the DNA methylation pattern of nine spermatozoa genes. J. Assist. Reprod. Genet. 2012, 29, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milekic, M.H.; Xin, Y.; O’Donnell, A.; Kumar, K.K.; Bradley-Moore, M.; Malaspina, D.; Moore, H.; Brunner, D.; Ge, Y.; Edwards, J.; et al. Age-Related sperm DNA methylation changes are transmitted to offspring and associated with abnormal behavior and dysregulated gene expression. Mol. Psychiatry 2015, 20, 995–1001. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J.; Bondarenko, O.; Linhart, O. Sperm motility in fishes: (III) diversity of regulatory signals from membrane to the axoneme. Theriogenology 2019, 136, 143–165. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Linhart, O.; Coward, K.; Rodina, M. Implication for aquaculture management. In Fish Spermatology; Alavi, S.M.H., Cosson, J., Coward, K., Rafiee, G., Eds.; Alpha Science Ltd.: Oxford, UK, 2008; pp. 397–460. [Google Scholar]
- Billard, R.; Cosson, J.; Perchec, G.; Linhart, O. Biology of sperm and artificial reproduction in carp. Aquaculture 1995, 129, 95–112. [Google Scholar] [CrossRef]
- Linhart, O.; Kudo, S.; Billard, R.; Slechta, V.; Mikodina, E.V. Morphology, composition and fertilization of carp eggs: A review. Aquaculture 1995, 129, 75–93. [Google Scholar] [CrossRef]
- Stoss, J.; Holtz, W. Successful storage of chilled rainbow trout (Salmo gairdneri) spermatozoa for up to 34 days. Aquaculture 1983, 31, 269–274. [Google Scholar] [CrossRef]
- Sadeghi, S.; Carles, J.N.; Silvestre, S.M.A. Effect of the activation media with different osmolality and cool storage on spermatozoa motility parameters over time in zebrafish, Danio rerio. Turk. J. Fish. Aquat. Sci. 2017, 17, 111–120. [Google Scholar] [CrossRef]
- Rurangwa, E.; Kime, D.E.; Ollevier, F.; Nash, J.P. The measurement of sperm motility and factors affecting sperm quality in cultured fish. Aquaculture 2004, 234, 1–28. [Google Scholar] [CrossRef]
- Dietrich, M.A.; Judycka, S.; Słowińska, M.; Kodzik, N.; Ciereszko, A. Short-Term storage-induced changes in the proteome of carp (Cyprinus carpio L.) spermatozoa. Aquaculture 2021, 530, 735784. [Google Scholar] [CrossRef]
- Cosson, J.; Billard, R.; Cibert, C.; Dreanno, C.; Suquet, M. Ionic factors regulating the motility of fish sperm. In The Male Gamete: From Basic Science to Clinical Applications; Gagnon, C., Ed.; Cache River Press: Vienna, Austria, 1999; pp. 161–186. [Google Scholar]
- Gallego, V.; Pérez, L.; Asturiano, J.F.; Yoshida, M. Relationship between spermatozoa motility parameters, sperm/egg ratio, and fertilization and hatching rates in pufferfish (Takifugu niphobles). Aquaculture 2013, 416, 238–243. [Google Scholar] [CrossRef]
- Ottesen, O.H.; Babiak, I.; Dahle, G. Sperm competition and fertilization success of Atlantic halibut (Hippoglossus hippoglossus L.). Aquaculture 2009, 286, 240–245. [Google Scholar] [CrossRef]
- Linhart, O.; Rodina, M.; Cosson, J. Cryopreservation of sperm in common carp Cyprinus carpio: Sperm motility and hatching success of embryos. Cryobiology 2000, 41, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Poupard, G.P.; Paxion, C.; Cosson, J.; Jeulin, C.; Fierville, F.; Billard, R. Initiation of carp spermatozoa motility and early ATP reduction after milt contamination by urine. Aquaculture 1998, 160, 317–328. [Google Scholar] [CrossRef]
- Hassan, M.M.; Nahiduzzaman, M.; Al Mamun, S.N.; Taher, M.A.; Hossain, M.A.R. Fertilization by refrigerator stored sperm of the Indian major carp, Labeo calbasu (Hamilton, 1822). Aquac. Res. 2013, 45, 150–158. [Google Scholar] [CrossRef]
- Perchec, G.; Jeulin, C.; Cosson, J.; Andre, F.; Billard, R. Relationship between sperm ATP content and motility of carp spermatozoa. J. Cell Sci. 1995, 108, 747–753. [Google Scholar] [CrossRef]
- Rahi, D.; Dzyuba, B.; Xin, M.M.; Cheng, Y.; Dzyuba, V. Energy pathways associated with sustained spermatozoon motility in the endangered Siberian sturgeon Acipenser baerii. J. Fish Biol. 2020, 97, 435–443. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Berger, B.; Weismann, T.; Patzner, R.A. Motility of spermatozoa of Alburnus alburnus (Cyprinidae) and its relationship to seminal plasma composition and sperm metabolism. Fish Physiol. Biochem. 1996, 15, 167–179. [Google Scholar] [CrossRef]
- Taborsky, M. Sperm competition in fish: “Bourgeois” males and parasitic spawning. Trends Ecol. Evol. 1998, 13, 222–227. [Google Scholar] [CrossRef]
- Horvath, L.; Tamas, G.; Seagrave, C. Carp and Pond Fish Culture; Fishing News Books: Oxford, UK, 1992; p. 155. [Google Scholar]
- Billard, R.; Bieniarz, K.; Popek, W.; Epler, P.; Saad, A. Observations on a possible pheromonal stimulation of milt production in carp (Cyprinus carpio L.). Aquaculture 1989, 77, 387–392. [Google Scholar] [CrossRef]
- Zhukinskiy, V.N.; Alekseenko, V.R. Semen quality in common carp, Cyprinus carpio, and white amur, Ctenopharyngodon idella (Cyprinidae) in different periods of the spawning season and as influenced by extraction methods. J. Ichthyol. 1983, 23, 124–133. [Google Scholar]
- Weil, C.; Fostier, A.; Billard, R. Induced spawning (ovulation and spermiation) in carp and related species. In Aquaculture of Cyprinids; INRA: Paris, France, 1986; pp. 119–137. [Google Scholar]
- Courtois, F.; Billard, R.; Takashima, F. Stimulation of spermiation following repeated injection of carp pituitary homogenates in the carp. Nippon Suisan Gakkaishi 1986, 52, 995–997. [Google Scholar] [CrossRef]
- Saad, A.; Billard, R. Spermatozoa production and volume of semen collected after hormonal stimulation in the carp, Cyprinus carpio. Aquaculture 1987, 65, 67–77. [Google Scholar] [CrossRef]
- Anway, M.D.; Cupp, A.S.; Uzumcu, M.; Skinner, M.K. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005, 308, 1466–1469. [Google Scholar] [CrossRef] [Green Version]
- Sujit, K.M.; Sarkar, S.; Singh, V.; Pandey, R.; Agrawal, N.K.; Trivedi, S.; Singh, K.; Gupta, G.; Rajender, S. Genome-Wide differential methylation analyses identifies methylation signatures of male infertility. Hum. Reprod. 2018, 33, 2256–2267. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.G.; Aston, K.I.; Pflueger, C.; Cairns, B.R.; Carrell, D.T. Age-Associated sperm DNA methylation alterations: Possible implications in offspring disease susceptibility. PLoS Genet. 2014, 10, e1004458. [Google Scholar] [CrossRef]
- Takeda, K.; Kobayashi, E.; Nishino, K.; Imai, A.; Adachi, H.; Hoshino, Y.; Iwao, K.; Akagi, S.; Kaneda, M.; Watanabe, S. Age-related changes in DNA methylation levels at cpg sites in bull spermatozoa and in vitro fertilization-derived blastocyst-stage embryos revealed by combined bisulfite restriction analysis. J. Reprod. Dev. 2019, 65, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luján, S.; Caroppo, E.; Niederberger, C.; Arce, J.C.; Sadler-Riggleman, I.; Beck, D.; Nilsson, E.; Skinner, M.K. Sperm DNA methylation epimutation biomarkers for male infertility and FSH therapeutic responsiveness. Sci. Rep. 2019, 9, 16786. [Google Scholar] [CrossRef] [Green Version]
- Potabattula, R.; Zacchini, F.; Ptak, G.E.; Dittrich, M.; Müller, T.; Hajj, N.E.; Hahn, T.; Drummer, C.; Behr, R.; Lucas-Hahn, A.; et al. Increasing methylation of sperm rDNA and other repetitive elements in the aging male mammalian germline. Aging Cell 2020, 19, e13181. [Google Scholar] [CrossRef]
- Oluwayiose, O.A.; Wu, H.T.; Saddiki, H.; Whitcomb, B.W.; Balzer, L.B.; Brandon, N.; Suvorov, A.; Tayyab, R.; Sites, C.K.; Hill, L.; et al. Sperm DNA methylation mediates the association of male age on reproductive outcomes among couples undergoing infertility treatment. Sci. Rep. 2021, 11, 3216. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Dzyuba, B.; Hulak, M.; Rodina, M.; Boryshpolets, S.; Li, Z.H.; Linhart, O. Percoll gradient separation of cryopreserved common carp spermatozoa to obtain a fraction with higher motility, velocity and membrane integrity. Theriogenology 2010, 74, 1356–1361. [Google Scholar] [CrossRef]
- Guerra, S.M.; Valcarce, D.G.; Cabrita, E.; Robles, V. Analysis of transcripts in gilthead seabream sperm and zebrafish testicular cells: MRNA profile as a predictor of gamete quality. Aquaculture 2013, 406–407, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.G.; Sarasquete, C.; Herráez, M.P.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Riesco, M.F.; Valcarce, D.G.; Martínez-Vázquez, J.M.; Robles, V. Effect of low sperm quality on progeny: A study on zebrafish as model species. Sci. Rep. 2019, 9, 11192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, K.A.; Schach, U.; Ordaz, A.; Steinfeld, J.S.; Draper, B.W.; Siegfried, K.R. Dmrt1 is necessary for male sexual development in zebrafish. Dev. Biol. 2017, 422, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, S.; Paasch, U.; Glander, H.J.; Anderegg, U. Mature human spermatozoa do not transcribe novel RNA. Andrologia 2005, 37, 69–71. [Google Scholar] [CrossRef]
- Bansal, S.K.; Gupta, N.; Sankhwar, S.N.; Rajender, S. Differential genes expression between fertile and infertile spermatozoa revealed by transcriptome analysis. PLoS ONE 2015, 10, e0127007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uysal, F.; Akkoyunlu, G.; Ozturk, S. DNA methyltransferases exhibit dynamic expression during spermatogenesis. Reprod. Biomed. Online 2016, 33, 690–702. [Google Scholar] [CrossRef] [Green Version]
- Kamiński, P.; Baszyński, J.; Jerzak, I.; Kavanagh, B.P.; Nowacka-Chiari, E.; Polanin, M.; Szymański, M.; Woźniak, A.; Kozera, W. External and genetic conditions determining male infertility. Int. J. Mol. Sci. 2020, 21, 5274. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.Z.; Vormer, T.L.; Jongejan, A.; Röling, M.D.; Silber, S.J.; de Rooij, D.G.; Hamer, G.; Repping, S.; van Pelt, A.M.M. Unraveling transcriptome dynamics in human spermatogenesis. Development 2017, 144, 3659–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, M.A.; Kumaresan, A.; Sinha, M.K.; Kamaraj, E.; Mohanty, T.K.; Datta, T.K.; Morrell, J.M. RNA-Seq analysis reveals functionally relevant coding and non-coding RNAs in crossbred bull spermatozoa. Anim. Reprod. Sci. 2020, 222, 106621. [Google Scholar] [CrossRef] [PubMed]
- Montjean, D.; De La Grange, P.; Gentien, D.; Rapinat, A.; Belloc, S.; Cohen-Bacrie, P.; Menezo, Y.; Benkhalifa, M. Sperm transcriptome profiling in oligozoospermia. J. Assist. Reprod. Genet. 2012, 29, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Zhou, X. Gene transcripts in spermatozoa: Markers of male infertility. Clin. Chim. Acta 2012, 413, 1035–1038. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Rajender, S. Long non-coding RNAs (lncRNAs) in spermatogenesis and male infertility. Reprod. Biol. Endocrinol. 2020, 18, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.F.; Robles, V. Cryopreservation causes genetic and epigenetic changes in zebrafish genital ridges. PLoS ONE 2013, 8, e67614. [Google Scholar] [CrossRef]
- Linhart, O.; Rodina, M.; Kašpar, V. Common carp (Cyprinus carpio Linneaus, 1758) male fertilization potency with secure number of spermatozoa per ova. J. Appl. Ichthyol. 2015, 31, 169–173. [Google Scholar] [CrossRef]
- Linhart, O.; Cheng, Y.; Xin, M.M.; Rodina, M.; Tučková, V.; Shelton, W.L.; Kašpar, V. Standardization of egg activation and fertilization in sterlet (Acipenser ruthenus). Aquac. Rep. 2020, 17, 100381. [Google Scholar] [CrossRef]
- Horokhovatskyi, Y.; Dietrich, M.A.; Lebeda, I.; Fedorov, P.; Rodina, M.; Dzyuba, B. Cryopreservation effects on a viable sperm sterlet (Acipenser ruthenus) subpopulation obtained by a Percoll density gradient method. PLoS ONE 2018, 13, e0202514. [Google Scholar] [CrossRef]
- Cheng, Y.; Xin, M.M.; Gela, D.; Rodina, M.; Tučková, V.; Kašpar, V.; Siddique, M.A.M.; Shelton, W.L.; Linhart, O. Optimization of sterlet (Acipenser ruthenus) egg incubation. Anim. Reprod. Sci. 2020, 215, 106334. [Google Scholar] [CrossRef]
- Andrews, S. A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 24 November 2010).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, F.; Andrews, S.R. Bismark: A flexible aligner and methylation caller for bisulfite-seq applications. Bioinformatics 2011, 27, 1571–1572. [Google Scholar] [CrossRef] [PubMed]
- Condon, D.E.; Tran, P.V.; Lien, Y.C.; Schug, J.; Georgieff, M.K.; Simmons, R.A.; Won, K.J. Defiant: (DMRs: Easy, fast, identification and ANnoTation) identifies differentially Methylated regions from iron-deficient rat hippocampus. BMC Bioinform. 2018, 19, 31–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramírez, F.; Vesztrocy, A.W.; Naldi, A.; Mungall, C.J.; Yunes, J.M.; Botvinnik, O.; Weigel, M.; et al. GOATOOLS: A python library for gene ontology analyses. Sci. Rep. 2018, 8, 10872. [Google Scholar] [CrossRef]
- Core R Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 12 December 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Library | CpG Methylation State | DMRs | No. of Genes | No. of Intergenic Regions |
|---|---|---|---|---|
| 24 HPS | methylated | 920 | 123 | 797 |
| unmethylated | 385 | 258 | 127 | |
| 96 HPS | methylated | 875 | 300 | 575 |
| unmethylated | 854 | 288 | 566 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Vechtova, P.; Fussy, Z.; Sterba, J.; Linhartová, Z.; Rodina, M.; Tučková, V.; Gela, D.; Samarin, A.M.; Lebeda, I.; et al. Changes in Phenotypes and DNA Methylation of In Vitro Aging Sperm in Common Carp Cyprinus carpio. Int. J. Mol. Sci. 2021, 22, 5925. https://doi.org/10.3390/ijms22115925
Cheng Y, Vechtova P, Fussy Z, Sterba J, Linhartová Z, Rodina M, Tučková V, Gela D, Samarin AM, Lebeda I, et al. Changes in Phenotypes and DNA Methylation of In Vitro Aging Sperm in Common Carp Cyprinus carpio. International Journal of Molecular Sciences. 2021; 22(11):5925. https://doi.org/10.3390/ijms22115925
Chicago/Turabian StyleCheng, Yu, Pavlina Vechtova, Zoltan Fussy, Jan Sterba, Zuzana Linhartová, Marek Rodina, Vladimíra Tučková, David Gela, Azin Mohagheghi Samarin, Ievgen Lebeda, and et al. 2021. "Changes in Phenotypes and DNA Methylation of In Vitro Aging Sperm in Common Carp Cyprinus carpio" International Journal of Molecular Sciences 22, no. 11: 5925. https://doi.org/10.3390/ijms22115925
APA StyleCheng, Y., Vechtova, P., Fussy, Z., Sterba, J., Linhartová, Z., Rodina, M., Tučková, V., Gela, D., Samarin, A. M., Lebeda, I., Xin, M., Zhang, S., Rahi, D., & Linhart, O. (2021). Changes in Phenotypes and DNA Methylation of In Vitro Aging Sperm in Common Carp Cyprinus carpio. International Journal of Molecular Sciences, 22(11), 5925. https://doi.org/10.3390/ijms22115925